Абиотические факторы
Температура среды. Среди абиотических факторов важнейшее значение в создании условий жизненной среды имеют климатические факторы - тепло, влажность, свет, движение воздуха. Из них особо важную роль для насекомых приобретает тепловой, или термический, фактор. Объясняется это тем, что насекомые, как уже указывалось в разделе "Анатомия и физиология", являются пойкилотермными организмами, т. е. имеют непостоянную температуру тела. В связи с этим жизнедеятельность насекомых, их поведение, скорость развития, а также и популяционная динамика нередко определяются температурными условиями среды; эти условия часто приобретают значение главнейшего, или ведущего экологического фактора.
Температура тела насекомого и его состояние закономерно изменяются при изменении внешних температур (рис. 172). Активная жизнедеятельность насекомого возможна лишь в пределах определенного диапазона температур, который у разных видов может быть различным; на рисунке 172 этот диапазон находится между 15-38°С, при оптимуме 26°С. Выше и ниже этой температурной зоны активности насекомые утрачивают способность быть активными, впадают в состояние либо теплового, либо холодового оцепенения (окоченения); однако при этом еще сохраняется способность вернуться к активному состоянию, если температура среды вновь вернется в пределы зоны активности.
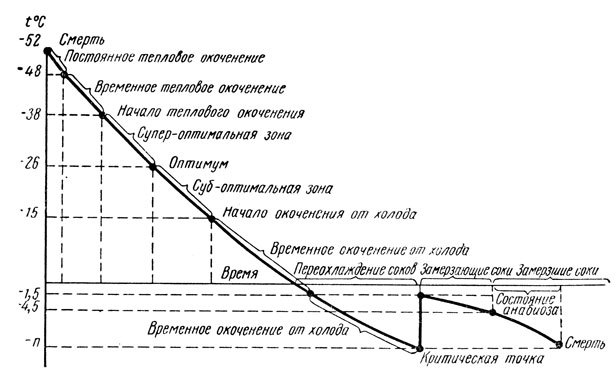
Рис. 172. Кривая Бахметьева, характеризующая изменение состояния насекомого под воздействием температуры среды (из Яхонтова по Бахметьеву)
Но слишком сильное повышение или понижение температуры среды, выходящее за пределы физиологических возможностей особи или вида, вызовет гибель организма. Произойдет полное нарушение соответствия организма и среды, возникнет дисгармония жизненных процессов, т. е. необратимые биохимические изменения в теле.
Понижение температуры среды ниже 0°С вначале вызывает переохлаждение соков: они остаются незамерзающими вследствие содержания в них солей. При дальнейшем охлаждении среды понижение температуры соков может достичь так называемой критической точки, или точки максимального переохлаждения. В этом случае произойдет частичное замерзание и частичная кристаллизация соков с освобождением скрытой теплоты плавления; последняя на короткий промежуток времени поднимает температуру тела. Такое поднятие температуры соков тела приобретает значение последней защитной реакции организма и может спасти жизнь. Но дальнейшее понижение температуры среды вызывает новое падение температуры соков, окончательное их замерзание и гибель насекомого. Такие закономерности воздействия температуры на насекомых впервые были вскрыты П. И. Бахметьевым в самом конце XIX - начале XX в.; вследствие этого изображенная на рис. 172 кривая нередко обозначается как кривая Бахметьева.
Холодостойкость, или способность переживать воздействие пониженных температур, имеет очень большое значение в динамике численности и в распространении насекомых. Начатая изучением еще Бахметьевым, холодостойкость насекомых изучалась впоследствии многими авторами.
Как оказалось, холодостойкость не является величиной постоянной для одного и того же вида и в сильной степени изменяется в зависимости от его физиологического состояния и биохимических особенностей. С другой стороны, разные виды обладают различной холодостойкостью; одни из них, особенно виды из более теплого климата, гибнут при сравнительно незначительных опусканиях температуры ниже 0°С, другие же способны выдерживать значительное охлаждение, в эксперименте - до -30, -50 и даже до -80°С. Объясняется это тем, что у многих насекомых гибель наступает вскоре после начала кристаллизации воды в их теле, т. е. вскоре по достижении критической точки, или точки максимального переохлаждения, тогда как у немногих других видов имеется способность к глубокому охлаждению, при котором они выдерживают воздействие температур, лежащих значительно ниже критической точки.
Переохлаждение соков играет физиологически защитную роль против кристаллизации воды, которая часто приводит к гибели насекомого. При этом оказалось, что уровень холодостойкости и, следовательно, точки максимального переохлаждения, зависит в высокой степени от содержания воды в организме. Это содержание определяется процентным количеством всей воды по отношению к сырому весу тела.
Так, активные гусеницы стеблевого мотылька при содержании в теле 67-69% воды имели критическую точку -11 -15°С; у диапаузирующих гусениц при содержании воды 56-57% точка максимального переохлаждения понизилась до -21 -25°С. Установлено также, что холодостойкость некоторых видов может быть повышена путем искусственного отнятия у них воды, например, при помещении их в эксикатор с хлористым кальцием или в другую сухую среду. Например, у личинок комнатной мухи подсушивание их тела со снижением веса на 4% понизило их точку переохлаждения с -12 до -22°С. Эти опыты с искусственной холодовой закалкой также свидетельствуют о зависимости холодостойкости от содержания воды в организме.
Однако в других случаях такой простой зависимости не получается. Оказалось, что нередко холодостойкость определяется не одним содержанием воды в организме, а ее физическим состоянием. Так, опытами Н. Л. Сахарова было установлено, что хорошо выкормившиеся, богатые резервами гусеницы озимой совки вымерзают только при -11°С, тогда как у водянистых недокормившихся гусениц с недостаточным содержанием резервов в теле холодостойкость снижается и они гибнут уже при -6°С. Это явление объясняется тем, что у хорошо выкормившихся упитанных особей насекомых в теле накапливаются гидрофильные коллоиды, например, в виде гликогена, способные удерживать воду как на своей поверхности, так, возможно, и путем химической связи. Такая прочно удерживаемая вода получила название связанной воды; она иногда составляет до 50% и более от всей содержащейся в теле воды и отличается способностью к большому переохлаждению. Этим связанная вода может быть противопоставлена свободной воде, легко теряемой организмом; потеря эта происходит вследствие отсутствия в нем физических и химических механизмов, удерживающих свободную воду. Наоборот, связанная вода оказывает большое сопротивление воздействию холода и замерзает при более низких температурах, нежели свободная вода. Замерзание же воды при высоком содержании ее в организме приводит к его обезвоживанию и к необратимым изменениям протоплазмы, влекущим за собою смерть.
Помимо роли связанной воды, существенное значение в повышении холодостойкости приписывается также и интермицеллярной воде - она заполняет ультрамикроскопические капиллярные пространства в протоплазме (Р. С. Ушатинская). Интермицеллярная вода, как и связанная, обладает большой способностью к переохлаждению в силу физических причин; известно, что помещенная в капилляры обыкновенная вода может быть тем сильнее переохлаждена, чем меньше диаметр капилляров.
Таким образом, холодостойкость насекомого обусловливается как содержанием воды в организме, так и ее физическим состоянием, т. е. соотношением между свободной водой, с одной стороны, и связанной и интермицеллярной водой - с другой.
Скорость развития насекомых как пойкилотермных животных в значительной мере зависит от температурных условий среды. Как оказалось, развитие насекомого может протекать лишь в пределах определенных температурных границ - верхней и нижней; выше верхней температурной границы и ниже нижней температурной границы физиологические процессы тормозятся и развитие прекращается. Соответственно сказанному эти границы получили название верхнего и нижнего порогов развития; иногда их называют также биологическими нулями развития.
Температуры, лежащие выше нижнего порога развития и не выходящие за пределы верхнего порога, получили названия эффективных температур; только они могут обеспечить развитие насекомого, т. е. только они обладают способностью вызвать активизацию физиологических процессов и дать надлежащий эффект - вывести насекомое из нулевого состояния развития.
Определение величины эффективной температуры невозможно без знания нижнего порога развития. Если последний известен, то эффективная температура легко узнается по наблюдаемой температуре (t) с вычетом температуры порога развития (t1), т. е. по разности t-t1. Например, наблюдаемая температура равна 25°С, а порог развития 10°С; следовательно, эффективная температура равна 15°С.
Для завершения всего своего развития каждому виду насекомого необходимо определенное количество тепловой энергии; это ее количество обозначается понятием сумма эффективных температур, или просто сумма тепла (C). Она может быть определена путем перемножения эффективной температуры (t-t1) на продолжительность развития в днях (n), т. е. по формуле
Эта зависимость длительности и скорости развития от температуры может быть показана и графически (рис. 173). Линия длительности развития при этом приобретает характер гиперболы, а линия скорости развития в процентах - прямой.
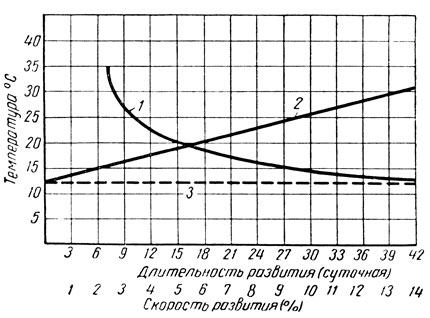
Рис. 173. Схема зависимости развития насекомого от температуры (по Кожанчикову): 1 - длительность развития в сутках, 2 - скорость развития в процентах, 3 - порог развития
Как показали многочисленные исследования, сумма эффективных температур, необходимая для развития особей того или иного вида, является для этого вида величиной более или менее постоянной, если нет осложняющих влияний или если прочие условия жизни находятся в оптимуме. В связи со сказанным, сумма эффективных температур, необходимая для развития одной полной генерации, нередко обозначают также понятием термальная константа. Она является довольна характерным видовым признаком и величина ее определяет степень требовательности вида к теплу как к экологическому фактору.
Ниже приводятся данные о пороге развития и сумме эффективных температур у некоторых видов насекомых (по И. В. Кожанчикову и другим авторам).

Таблица 1
На основании этих данных можно примерно определить продолжительность развития при тех или иных постоянных температурах. Так, например, при 21°С амбарный долгоносик будет развиваться около 36 дней: (21-11)×36=360, а комнатная муха -25,5 дней: (21-12)×25,5=230.
По порогу развития и термальной константе можно рассчитать также число возможных поколений того или иного вида в какой-либо географической точке. Для этого необходимо взять средние многолетние температуры за все месяца вегетационного периода и подсчитать сумму тепла, т. е. эффективных температур, которая набирается в данной географической точке выше порога развития изучаемого вида.
Например, расчет показывает, что в районе Ленинграда (г. Пушкин) в течение мая - сентября набирается примерно 740°С эффективных температур при пороге 8°С.

Таблица 2
Сопоставление этой полученной для окрестностей Ленинграда суммы эффективных температур с термальной константой шведской мухи создает впечатление, что она здесь не имеет достаточного тепла для развития двух полных поколений. Фактически же шведская муха имеет здесь три поколения.
Несовпадение расчетных и фактических данных объясняется тем, что шведская муха является обитателем приземного слоя воздуха, который имеет более высокую температуру, нежели метеорологические показатели, полученные в специальной будке. Следовательно, необходимо ввести поправку на разницу между показаниями температуры в метеорологической будке и в приземном слое воздуха; наблюдения показывают, что эта разница достигает в среднем 3-5°С. Если принять для района Ленинграда разницу в 3°С, то к цифре 739° надо добавить еще 459° (получаемых от перемножения 153 дней на 3); таким образом, получается цифра 1198°, что примерно и соответствует трем поколениям шведской мухи.
Применяя ту же методику, можно рассчитать число возможных генераций в разных зонах СССР, например, у капустной моли, которая также живет в приземном слое воздуха.

Таблица 3
Приведенные расчеты довольно хорошо совпадают с фактическим числом генераций капустной моли в перечисленных географических точках; разница может быть объяснена тем, что расчет сделан по средним многолетним температурам, тогда как в отдельные жаркие или прохладные годы набирающаяся сумма тепла может быть больше или меньше средних показателей.
Суммы эффективных температур используются также для определения сроков возможного появления в природе тех или иных фаз развития вредителей, сроков яйцекладки и пр.
Однако постоянство суммы эффективных температур не следует преувеличивать. Она не является константой в математическом смысле, так как обнаруживает изменчивость при отклонении условий среды от оптимума и у особей из разных частей ареала. Кроме того, скорость развития, а отсюда и продолжительность его может сильно изменяться под воздействием длины фотопериода, условий питания, сухости воздуха. Эти факторы осложняют развитие явлением диапаузы, благодаря чему расчет числа генераций или сроков появления той или иной фазы только на основании термальной константы может привести к ложным данным. Она является всего лишь примерной биологической величиной, устойчивой только в строго определенном диапазоне других факторов среды. Из этого следует, что расчет возможного числа поколений и фенологических показателей вида на основании суммы эффективных температур следует производить с большой осмотрительностью и лишь в том случае, если есть уверенность, что другие факторы среды не искажают роль термического фактора.
Влажность среды. Показатели влажности среды разнообразны, но в экологии насекомых наибольшее значение имеет относительная влажность воздуха, т. е. процент насыщения его водяными парами. Действие влажности оказывается разнообразно; оно изменяет содержание воды в тканях насекомого, а в связи с этим влияет на его поведение, его выживаемость и плодовитость. В одних случаях влажность среды действует непосредственно, в других - она действует через посредство принимаемой насекомым пищи или через другие факторы среды.
Обладая малыми размерами тела и, следовательно, большой поверхностью испарения, насекомые оказываются очень зависимыми от влажности среды как экологического фактора. Особенно это относится к обитателям открытой воздушной среды; здесь относительная влажность воздуха лишь в особых случаях достигает 100%, в большинстве же она ниже точки росы, что неизбежно создает условия для испарения насекомыми воды. Удержание в организме влаги в нужных пределах требует специальных механизмов как особых приспособлений для поддержания должного равновесия между организмом и средой. Такими приспособительными механизмами, регулирующими водный обмен у насекомых, являются морфологические, физиологические и экологические адаптации.
К числу морфологических приспособлений, или адаптации, относятся развитие на покровах водонепроницаемой эпикутикулы, образование на теле у ряда тлей, червецов и других насекомых воскового налета или покрова, утолщение кутикулы, строение дыхалец, строение куколок и образование кокона и пр.
Вопрос о гигрофобной функции эпикутикулы уже рассматривался раньше; аналогичную функцию выполняют и восковые образования на коже. Что касается толщины кутикулы, то особенно значительна она у многих обитателей пустынь и сухих степей из числа саранчовых, кузнечиков, тараканов, жуков и других насекомых; в частности, среди жуков очень толстая кутикула свойственна характерным обитателям сухих ландшафтов - чернотелкам.
Строение дыхалец играет также существенную роль в регулировании испарения в разных по увлажнению средах, так как испарение воды из тела насекомых происходит в значительной степени с поверхности трахейной системы через дыхальца. В связи с этим особо большое значение может иметь размер дыхалец. Показателем величины дыхалец является так называемый дыхальцевый индекс - процентное отношение длины переднегрудного дыхальца к длине всей груди.
Оказалось, что у ряда кровососущих двукрылых, как комары, москиты и мокрецы, величина дыхальцевого индекса характеризует степень влаголюбивости видов: более влаголюбивые виды имеют и больший дыхальцевый индекс, следовательно, испаряют воду менее экономно и способны жить только во влажной среде, тогда как сухоустойчивые виды имеют малый дыхальцевый индекс, испаряют более экономно и способны жить в более сухой среде.
Например, белый малярийный комар (Anopheles pulcherrimus Theob.) имеет малые размеры дыхалец (дыхальцевый индекс 7, 53), поэтому отличается большой сухоустойчивостью и способен в дневные часы оставаться в местах с сухим воздухом и высокой температурой - среди зарослей растений, в сараях, пещерах и пр. Другой вид малярийного комара - A. algeriensis Theob., наоборот, чрезвычайно влаголюбив, имеет большие дыхальца (дыхальцевый индекс 10,75) и при попадании в сухой воздух быстро гибнет.
В особенностях строения куколок насекомых с полным превращением также можно найти черты приспособления к жизни в условиях различной влажности. Так, открытые куколки жуков и перепончатокрылых имеют нежные, тонкие покровы, легко проницаемые для воды и газов, и у многих видов располагаются в почве, обычно в земляных колыбельках, где влажность воздуха близка к 100%. Развитие таких открытых куколок в воздушной среде было бы сопряжено с большой потерей влаги и неизбежной гибелью; но у открыто живущих видов куколка заключена в кокон, который и предохраняет от излишней потери влаги. У имеющих покрытую куколку чешуекрылых образование кокона наблюдается обычно у видов, окукляющихся открыто на растениях (коконопряды, шелкопряды, павлиноглазки и др.) или в поверхностном, легко высыхающем горизонте почвы (луговой мотылек из огневок); виды же, окукляющиеся глубоко в почве, где влажность всегда высокая, не делают кокона (совки). Иногда один и тот же вид насекомого ведет себя по-разному в условиях различного увлажнения: во влажной среде не делает кокона, а в сухой - прядет его; это, например, отмечено у личинок жука известного амбарного вредителя - рыжего мукоеда (Laemophloeus ferrugineus St.) из семейства плоскотелок (Cucujidae). Значение кокона как приспособления к уменьшению потери влаги особенно наглядно вытекает из такого опыта: у вынутых из шелкового кокона куколок тутового шелкопряда потеря веса в 5 раз больше, чем у нормально развивающихся, т. е. заключенных в коконе куколок.
Жизнь под щитком характерна для многих кокцид: у них под щитком происходит развитие яйца и большая часть жизни вида. У яблонной моли (Hyponomeuta mallinelus Zell.) яйца также откладываются под щиток, образованный выделениями придаточных половых желез; здесь же происходит зимовка гусениц, которые весной вбуравливаются в листья, делают в них мины и, следовательно, живут под защитой тканей растения. Лишь позднее, достигнув более крупных размеров, гусеницы начинают вести открытый образ жизни.
К числу физиологических механизмов регулирования водного баланса тела насекомого относятся такие приспособления, как отсасывание воды задней кишкой из непереваренных остатков пищи, как поглощение влаги покровами при контакте с ней, как поступление влаги в организм с пищей и пр.
Что касается экологических приспособлений, то они прежде всего проявляются в изменении местообитания особи. Так, почвообитающие насекомые при подсыхании верхних горизонтов почвы совершают вертикальные миграции, спускаясь в более глубокие слои, где сохраняется 100% влажность воздуха или близкие к ней показатели. При увлажнении же верхних горизонтов происходит перемещение вверх. Вертикальные миграции отмечены у личинок жуков, - таких, как проволочники (личинки щелкунов) и личинки хрущей, у почвообитающих гусениц и др. Подвижные наземные формы насекомых - такие, как саранчовые, кузнечиковые, цикадовые, клопы и др. в засушливое время перемещаются на более пониженные, сырые места.
Многие насекомые перед окуклением, зимовкой или диапаузой перемещаются на новые места, чтобы поставить себя в условия высокой влажности. Одни из них собираются под растительными остатками и в лесной подстилке; к ним относятся клопы-черепашки, многие долгоносики, например, гороховые, клеверный и другие, земляные блошки и пр. Другие виды уходят для переживания этих периодов жизни в почву, где влажность среды более постоянна, чем в наземных условиях; таковы гусеницы многих совок, многие жуки-долгоносики и пр. Некоторые укрываются в трещинах на коре деревьев или под отставшей корой или в дуплах, например, гусеницы яблонной плодожорки; другие делают зимние гнезда, например, гусеницы бабочки-златогузки; третьи остаются в тканях растений - в стеблях, в галлах и пр., например, трипсы, гусеницы и куколки бабочек-стеклянниц, личинки и куколки некоторых жуков-усачей и др.; четвертые скручивают листья, например, тли, жуки-трубковерты, листовертки и пр. Усиленное выделение тлями медвяной росы также рассматривается как приспособление к жизни в условиях недостаточной влажности воздуха. Пропуская через кишечник большое количество воды, эти насекомые тем самым препятствуют высыханию тела, создавая и в своих тканях и в окружающей воздушной среде высокое содержание влаги; в условиях насыщенного влагой воздуха выделение медвяной росы прекращается.
Однако по степени требования к влажности среды насекомые неоднородны, проявляют различную избирательность и среди них можно различать три группы видов: крайне влаголюбивых, или гигрофилов, средне влаголюбивых, или мезофилов, и сухолюбивых, или ксерофилов. Может наблюдаться и такое положение, когда в одних фазах развития насекомое является ксерофилом, а в других - мезо- или гигрофилом; в частности, повышенная требовательность к влажности среды часто наблюдается в фазе яйца, молодых личиночных стадиях и в фазе куколки, тогда как более развитые личинки и взрослые особи нередко оказываются ксеро- или мезофилами. Выше, при рассмотрении приспособлений к регулированию водного баланса тела, уже приводились примеры высокой требовательности к влажности среды куколок ряда насекомых. Известно также, что при засухе и суховеях может происходить массовая гибель яиц гессенской и шведской мухи; с другой стороны, у такого довольно сухолюбивого вида, как мароккская саранча, слишком сырая весна может вызвать гибель перезимовавших в почве яиц.
В соответствии со своей избирательностью к влажности среды различные виды заселяют неодинаковые местообитания. Гигрофильные виды (или те или иные фазы их развития) заселяют сырые места, мезофильные - среднеувлажненные, а ксерофилы - сухие местообитания. Так, например, стеблевой мотылек может считаться гигрофилом, так как явно предпочитает такие местообитания, как долины рек и другие пониженные места с избыточным увлажнением; оптимальная относительная влажность воздуха у этого вида находится в пределах 80-100%.
Озимая совка может считаться мезофилом, так как ее оптимум относительной влажности воздуха лежит в пределах 50-80%; поэтому она обычно заселяет такие места, как паровые поля, пропашные культуры и другие аналогичные местообитания и лишь на крайнем юге переходит на более увлажненные участки. Примером ксерофильного насекомого может быть пустынная саранча (Schistocerca gregaria Forsk.), распространенная в сухих районах Африки и Юго-Западной Азии; оптимальная влажность воздуха для взрослой фазы пустынной саранчи равна примерно 40-60%.
Однако все эти показатели являются в значительной степени условными, так как в реальной природе действие влажности на насекомых часто практически неотделимо от действия других экологических факторов, в первую очередь температуры и пищи.
Влияние влажности через посредство пищи особенно хорошо прослеживается на амбарных вредителях. Так, амбарный долгоносик питается зерном с содержанием влаги не более 20%, но наиболее оптимальные условия для жизни и размножения создаются при 14-16% влажности. Зерно с влажностью 12% тормозит размножение долгоносика, а при 11% он гибнет. На этом основан один из важнейших предупредительных методов борьбы с данным вредителем - хранение хорошо просушенного зерна.
Недостаток влаги в природе во время засухи может ухудшать условия питания у ряда растениеядных видов. Так, известны случаи массовой гибели личинок саранчовых при сильном выгорании травянистой растительности, служащей пищей этим насекомым. Засуха также угнетает цветение растений и резко сокращает накопление нектара в цветках. Все это сильно ухудшает нектарное дополнительное питание бабочек озимой совки, лугового мотылька и других потребителей нектара; в результате у таких видов происходит резкое снижение плодовитости, а иногда наблюдается и полное бесплодие. Недостаток капельной влаги (росы и пр.) снижает плодовитость яблонной плодожорки.
Засуха и высокая температура воздуха приводит к образованию в семеизвергательном канале самца лугового мотылька сгустка в виде пробки, препятствующей продвижению семенной жидкости; такие самцы практически становятся стерильными. С другой стороны, сухой воздух и высокая температура способствуют повышению концентрации Сахаров в цветочном нектаре; как оказалось, высокая концентрация Сахаров (свыше 25%) вызывает у самок лугового мотылька дегенерацию и рассасывание зачатков яиц в яйцевых трубочках.
Совместное действие температуры и влажности. Температура среды в сильнейшей степени может изменить характер воздействия влажности на насекомых. В связи с этим многие авторы особо подчеркивают необходимость изучения совместного влияния различных показателей температуры и влажности на скорость развития, состояние и плодовитость насекомых.
Так, лабораторные исследования смертности куколок яблонной плодожорки при совместном воздействии температуры и влажности позволили выявить оптимальные и пессимальные их показатели для данного вида (рис. 174); полученные таким методом графики называются термогигрограммами. Оказалось, что минимальная смертность (не более 10%) у этого вида наблюдается в пределах 53-95% относительной влажности при температуре 21-28°; за пределами этих показателей, особенно при сильном снижении влажности воздуха, а также при сильном снижении или повышении температуры, смертность резко возрастает.
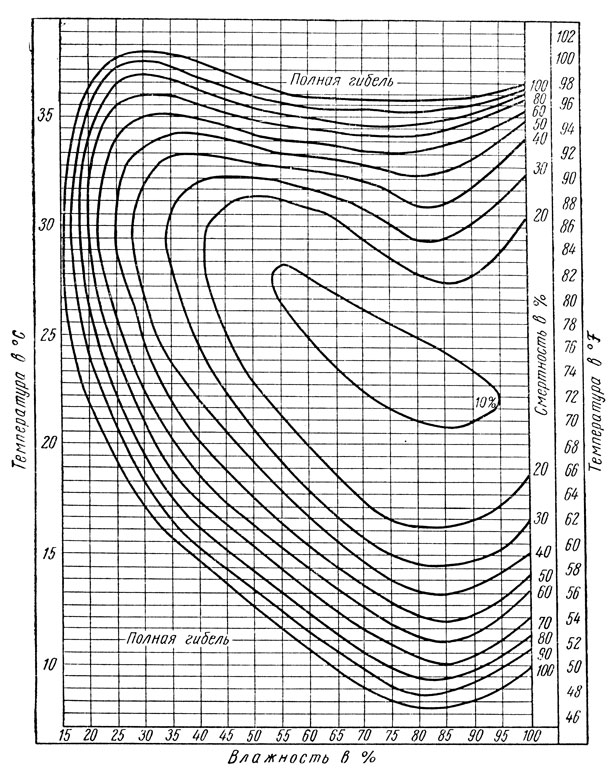
Рис. 174. Тсрмогигрограмма развития куколок яблонной плодожорки (на ординатах нанесена температура, на абсциссах - влажность в %; кривые дают смертность в %) (по Шелфорду)
Изменение длительности развития у куколок озимой совки показано на другой термогигрограмме (рис. 175). Из нее следует, что различные комбинации влажности и температуры среды могут изменять продолжительность развития куколок озимой совки в пределах широкого диапазона времени - от 10 до 70 суток. Плодовитость бабочек этого же вида также сильно изменяется в зависимости от сочетания температуры и влажности среды, в условиях которых развивались куколки (по И. В. Кожанчикову):

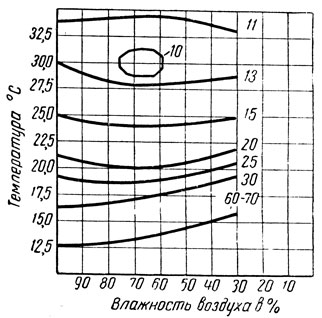
Рис. 175. Изменение длительности развития куколок озимой совки в сутках (цифры от 10 до 70 суток) в зависимости от сочетания температуры и влажности (по Кожанчикову)
Из приведенных данных следует, что наибольшая плодовитость наблюдается при 85% относительной влажности, но повышение температуры до 30°С способствует уменьшению числа отложенных яиц, особенно при 55 и 95% относительной влажности воздуха.
Таким образом, все основные показатели жизни и состояния насекомых - смертность, продолжительность развития и плодовитость - в сильнейшей степени зависят от конкретных сочетаний температуры и влажности среды:
Для оценки и анализа влияния сочетаний температуры и влажности на насекомых в природной обстановке применяется метод климограмм. Эти климограммы составляют по среднемесячным показателям температуры и количества выпавших осадков; по оси ординат откладываются температуры, по абсциссе - количество осадков (в миллиметрах). Точки пересечения затем последовательно соединяются от предшествующего месяца к последующему; в результате получается неправильный многоугольник, который и представляет собою климограмму (рис. 176). Вытянутость этой климограммы вверх говорит о жарком и сухом лете, вниз - о суровой зиме, вправо - о высокой влажности в соответствующие месяца и пр.
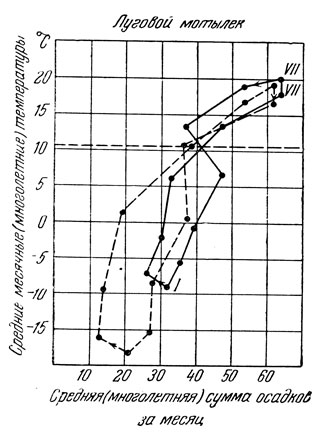
Рис. 176. Климограммы оптимума для лугового мотылька в Европейской части (сплошная линия) и в Сибири (прерывистая линия) (по Кожанчикову)
Такие климограммы строятся для анализа условий жизни того или иного вида в различных частях ареала, либо для анализа популяционной динамики, т. е. изменения численности особей вида во времени. Например, сопоставление климограмм, построенных для годов резкого возрастания численности особей взятого вида, с климограммами годов его численной депрессии может позволить вскрыть значение конкретных показателей температуры и количества осадков в создании благоприятных и неблагоприятных условий жизни этого вида.
Более удобной для экологического анализа условий жизни насекомых является биоклимограмма, предложенная Б. Уваровым (рис. 177) Но для ее построения необходимо знать сроки развития всех фаз в природе. Соответствующие линии многоугольника обозначаются по-разному для этих фаз; например, прерывистой линией обозначают фазу яйца, жирной разорванной линией - фазу личинки и сплошной линией - фазу взрослого насекомого.
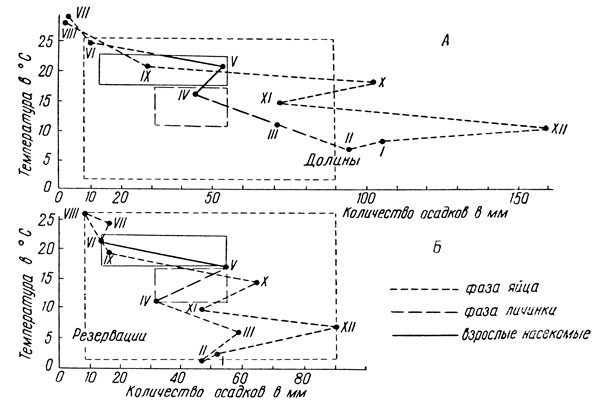
Рис. 177. Биоклимограмма Уварова для мароккской саранчи (по Уварову): А - неблагоприятные условия жизни; Б - благоприятные условия жизни
Если теперь построить такую биоклимограмму для Оптимальной части ареала или для годов массового появления вида, то таким путем будут выявлены наиболее благоприятные для него показатели температуры и влажности. Построив для всех фаз по крайним точкам их развития соответствующие четырехугольники, мы получаем показатели оптимальных условий жизни для яйца, личинки и взрослого насекомого. Эти четырехугольники затем наносятся на биоклимограммы неблагоприятных условий жизни вида; в результате выявляются те конкретные условия температуры и влажности, которые выходят за пределы оптимума и, следовательно, могут играть отрицательную роль в жизни анализируемого таким методом вида. На рисунке 177, А как раз и показано, что зимний период характеризуется излишним обилием осадков, что у мароккской саранчи вызывает гибель зимующих яиц; слишком влажным оказывается и период развития личинок, что может сопровождаться появлением у них грибковых заболеваний.
Соотношение температуры и влажности имеет существенное значение и при выборе видами для своей жизни соответствующих местообитаний. Особенности температурного режима и режима влажности в том или ином местообитании характеризуют его микроклимат, т. е. климат того приземного слоя воздуха, где произрастают растения и протекает жизнь многих животных. Этот микроклимат, следовательно, может быть охарактеризован соответствующими величинами температуры и влажности воздуха, т. е. гидротермическими показателями.
Так, например, сырой луг с густым травостоем будет отличаться высокой влажностью воздуха, близкой к 100%, и сравнительно невысокими дневными температурами, лежащими в пределах 15-30°С; расположенный же неподалеку возвышенный песчаный участок с редким травостоем будет отличаться более сухим и теплым микроклиматом - относительная влажность воздуха может здесь опускаться в дневное время до 20-40%, а температура воздуха может подниматься до 40-60°С.
В соответствии со своими требованиями к среде разные виды будут по-разному относиться к микроклимату местообитаний; одни из этих местообитаний будут заселяться одними видами, другие - комплексом других видов. На сухих песчаных участках будут встречаться жуки-чернотелки, сухолюбивые и теплолюбивые виды жуков-долгоносиков, саранчовых и других насекомых, тогда как фауна сырых лугов будет складываться из влаголюбивых и менее теплолюбивых видов - различных комаров, мошек, мокрецов, подур, соответствующих видов саранчовых, цикадок и пр.
Однако гидротермические показатели местообитаний зависят также от погодных условий и от климатической зоны. В связи с этим одно и то же местообитание может оказаться неодинаковым по микроклимату в годы с засушливой и жаркой погодой по сравнению с прохладными, сырыми годами; точно также в более южных климатических зонах микроклимат внешне одного и того же местообитания будет иметь иные гидротермические показатели нежели на севере. В результате таких гидротермических отклонений от нормы многие виды вынуждены менять свои местообитания, переселяясь в засушливые жаркие годы на более прохладные места. Сходным образом одни и те же виды на юге своего ареала заселяют более тенистые и влажные местообитания, чем на севере. Таким образом, изменение микроклиматических особенностей в местообитаниях во времени и в пространстве порождает у видов смену местообитаний. Как особый экологический принцип смена местообитаний будет рассмотрена более подробно ниже.
Действие света. Роль света как экологического фактора в жизни насекомых несомненно велика и часто не уступает, а в некоторых случаях даже превосходит роль других климатических факторов - температуры и влажности. Выше в разделе "Биология" уже отмечалась выдающаяся роль света как фактора, регулирующего годичный жизненный цикл видов: длинный фотопериод способствует беспрепятственному развитию многих видов, тогда как короткий фотопериод, наступающий в конце лета, - начале осени, стимулирует переход в состояние диапаузы.
Приспособительное значение такой реакции насекомого на короткий день очевидно и уже объяснялось. Напомним, что короткий фотопериод является точным и безошибочным сигналом скорого наступления неблагоприятного осенне-зимнего периода жизни, и переход в состояние диапаузы по этому сигналу обеспечивает виду своевременную физиологическую перестройку для подготовки к зимовке.
Оказалось также, что фотопериодическая реакция насекомых проявляется не только при сильной, но и при слабой освещенности: многие виды чувствительны к силе света, исчисляемой всего лишь 1-5 люксами. Иногда эта чувствительность еще более значительна, например, у колорадского жука, который обнаруживает первые признаки фотопериодической реакции даже при силе света в 0,1 люкс. Следовательно, даже очень слабая освещенность действует на многих насекомых, как "день", вызывая соответствующую фотопериодическую реакцию.
Такая высокая чувствительность к свету несомненно имеет приспособительное значение; благодаря ей исключается влияние на фотопериодическую реакцию колебаний интенсивности дневного света при смене погодных условий, например в безоблачные солнечные и пасмурные периоды дня.
Но действие света оказывается не только в том, что свет играет выдающуюся роль в формировании годичного и сезонного циклов развития. Свет влияет на жизнь насекомых и по-иному. Прежде всего насекомые различаются неодинаковой жизнедеятельностью в течение светлой и темной частей суток: одни виды активны при дневном свете, другие при сумеречном свете, третьи при ночном.
Например, вся группа дневных, или булавоусых, бабочек (Papilionoidea) относится к числу типично дневных насекомых; и, наоборот, бабочки из семейства совок, или ночниц (Noctuidae), активны ночью и в большинстве - типично ночные насекомые. Эта разная активность в течение суток нашла свое отражение и в самих названиях указанных систематических групп бабочек. К числу дневных насекомых относятся также саранчовые, многие двукрылые, жуки, перепончатокрылые и пр. С другой стороны, к ночным или сумеречным насекомым относятся многие ближайшие родичи саранчовых - кузнечики и сверчки; поэтому их стрекотание мы часто слышим вечером или ночью, тогда как у саранчовых оно наблюдается днем.
К характерным сумеречным насекомым относятся многие бабочки из семейства бражников (Sphingidae), а также многие хрущи, например майские и июньские и др. Днем эти виды не активны, но с заходом солнца начинают интенсивно летать; в это время они спариваются, а нуждающиеся в дополнительном питании - питаются. Заход солнца и снижение интенсивности освещения до определенного уровня становится для этих видов активизирующим сигналом. Нередки и виды, жизнедеятельность которых протекает не только в сумеречное, но и в ночное время. К числу таковых относятся некоторые бабочки из семейства огневок, например луговой мотылек, некоторые совки и др.
Существенный интерес представляет также реакция на искусственный свет ночных, сумеречных и дневных насекомых. Как оказалось, многие насекомые особенно активно летят ночью на ультрафиолетовое излучение (длина волны 300-440 ммк). Коротковолновое излучение этого диапазона привлекает наиболее разнообразный состав насекомых; при этом, по сравнению с обычной лампой накаливания (с длиной волны излучаемого света примерно 500-700 ммк), ультрафиолетовые лучи привлекают в 5-20 раз больше особей одного и того же вида. Установлено также, что интенсивность привлечения на свет в значительно меньшей степени зависит от мощности источника света, нежели от его спектрального состава.
Особо эффективным источником искусственного освещения ультрафиолетовыми лучами являются ртутные лампы. На свет этих ламп прилетают многие вредные виды насекомых; с другой стороны некоторые полезные насекомые-энтомофаги - такие, как сетчатокрылые, божьи коровки, отчасти наездники прилетают на ртутные лампы в меньшей численности, чем вредные виды. Указывается также на то, что значительная часть прилетающих на ультрафиолет вредителей находится в неполовозрелом состоянии; по мере старения особей их реакция на ультрафиолет падает. В связи со сказанным открываются перспективы использования ртутных ламп как одного из биофизических методов борьбы с вредными насекомыми, а также как метода учета численности и сроков появления вредителей.
Реакция на источник искусственного света может оказаться совершенно различной даже у близких видов. Так, в теплые ночи на юге СССР могут быть активными и некоторые саранчовые, которые в таких случаях прилетают на свет; к их числу относится, в частности, широко распространенный вид - пустынный прус (Calliptamus barbarus Costa). Но его ближайший родич и известный вредитель - итальянская саранча, или обыкновенный прус (C. italicus L.), на свет не летит.
Очевидно реакция насекомых на искусственный свет разнообразна: она может быть неодинаковой у разных, иногда даже близких видов, может изменяться от физиологического состояния - половой зрелости и возраста особей, она в сильнейшей степени зависит от спектрального состава излучения, температуры среды и пр.
Воздушные токи. Существенную роль как один из факторов расселения насекомых могут играть воздушные токи - горизонтальные и вертикальные. Многие мелкие насекомые, также как тли, мелкие двукрылые, бабочки и др., не способны оказывать при полете сопротивление ветру и поэтому, в сущности, имеют пассивный полет. Ветер как горизонтальный поток воздушных масс является для таких насекомых одним из важнейших факторов расселения.
Однако воздушные токи способствуют расселению в некоторых случаях и бескрылых насекомых и личинок, обладающих парусностью. К числу таковых относятся молодые, покрытые длинными волосками гусеницы бабочек; благодаря длинным волоскам и малому весу тела такие гусеницы обладают большой парусностью, подхватываются ветром и расселяются на местности. Примером могут быть гусеницы первого возраста непарного шелкопряда; их волоски вдвое длиннее самого тела. Такие гусеницы могут быть отнесены ветром на расстояние до 20 км, что позволяет этому виду быстро заселить территорию в период своего массового размножения.
Насекомые, обладающие активным полетом, также часто разносятся ветром. Так, пустынная саранча имеет сильный активный полет, но уже при силе ветра 7 км в час передвижение стай саранчи идет по ветру. При этом особи саранчи в летящей стае находятся в активном состоянии и держатся в воздухе совместно, но стая не имеет своего собственного направления полета. Следовательно, миграции стай саранчи за десятки и сотни километров представляют собою пассивное перемещение под влиянием ветра находящихся в активном движении особей. В целом стаи пустынной саранчи перемещаются из области высокого барометрического давления в область барометрического минимума и в общем их полет совпадает с направлением ветра. Барометрический минимум сопровождается выпадением осадков и, таким образом, осевшие массы саранчи оказываются в условиях высокой влажности среды, благоприятствующей быстрому половому созреванию и размножению саранчи. Особо благоприятной для пустынной саранчи является зона, расположенная между ветрами двух типов - муссонами и пассатами; в области встречи этих воздушных потоков выпадают обильные осадки, что и обеспечивает здесь оптимум условий для пустынной саранчи и вспышки ее численности.
Перемещению насекомых способствуют также и вертикальные воздушные токи, например, движение теплого воздуха вверх в дневные и вечерние часы суток. Эти воздушные токи подхватывают насекомых, перемещают их в более высокие горизонты, где они попадают под воздействие сильных и постоянных ветров и вынуждены совершать воздушный дрейф. Дрейфующие насекомые могут оказаться на высоте 1-2 тыс. метров и разносятся на сотни километров. Это относится к насекомым не только с активным, но и пассивным полетом.
Некоторые авторы перемещение насекомых в вечернее время из более низких воздушных горизонтов в более высокие объясняют создающейся в этот период температурной инверсией: приземные слои воздуха начинают охлаждаться скорее, нежели верхние слои, что и вызывает движение насекомых в сторону термического оптимума, т. е. вверх. Поднявшиеся таким путем особи попадают под воздействие горизонтальных токов и перемещаются на то или иное расстояние. Так, в частности, объясняют перелеты масс лугового мотылька. Другие исследователи считают стимулом вечернего подъема летящих насекомых вверх не температурную инверсию, а разность освещения, создающуюся на различных горизонтах после захода солнца; более сильная освещенность верхних горизонтов воздушной среды способствует перемещению сюда летающих насекомых вследствие их положительного фототаксиса.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'