Диапауза как регулятор жизненного цикла
Определяющие понятия. Диапауза представляет собою состояние временного физиологического покоя и возникает в жизненном цикле как специальное приспособление к переживанию неблагоприятных условий в областях с сезонно-периодическим климатом. В умеренном климате вырабатываются приспособления к жизни в условиях двух основных сезонов года - благоприятного летнего и неблагоприятного зимнего; в тропиках и субтропиках - к жизни во влажном и сухом сезонах.
Диапаузу нельзя смешивать с оцепенением, возникающим под влиянием низких и высоких температур среды. При холодовом и тепловом оцепенении физиологические процессы также затормаживаются, но это снижение или прекращение активности насекомого не имеет глубокого приспособительного значения; поэтому более или менее длительное холодовое или тепловое оцепенение приводит к неэкономному расходованию питательных резервов в организме, вызывает патологические изменения и может быстро привести организм к гибели. Диапауза же обеспечивает экономное использование резервов в организме и позволяет пережить длительные неблагоприятные периоды.
При диапаузе происходит задержка роста и развития особи в течение более или менее длительного срока. Эта задержка возникает под воздействием эндокринной системы на основе нейро-гуморального регулирующего механизма: нервная система контролирует деятельность эндокринной системы, а последняя обеспечивает взаимодействие между органами через посредство жидкой внутренней среды - гемолимфы. В свою очередь, эндокринные органы находятся под воздействием внешней среды; внешняя среда в сущности программирует жизненный цикл и диапаузу насекомого, вовлекая в работу нервную и эндокринную регулирующие системы, и соответственно настраивает их на выполнение необходимых физиологических процессов.
Следовательно, диапауза находится в сложном взаимоотношении с внешней средой. Более того, диапауза часто возникает задолго до наступления неблагоприятных условий среды и может даже продолжаться после того, как эти неблагоприятные условия уже прошли. Поэтому нельзя считать, что диапауза является лишь непосредственной реакцией на воздействие неблагоприятных условий. Механизм возникновения диапаузы значительно сложнее.
Как приспособление к жизни в условиях сезонно-периодического климата, диапауза находится под контролем тех факторов среды, которые имеют правильную сезонную периодичность. Такими факторами являются длина дня, температура и влажность воздуха, биохимическое состояние кормовых растений. В сезонно-периодическом климате эти факторы среды играют роль сигналов, предупреждающих наступление неблагоприятных для жизни сезонов - зимы в умеренных странах и сухого периода в тропиках и субтропиках. Взаимосвязанное воздействие этих сигнальных факторов среды и вызывает диапаузу у насекомых.
Среди указанных периодических факторов самым совершенным, всегда астрономически точным сигнализатором предстоящего наступления неблагоприятного сезона является длина фотопериода. Сокращение длины дня в странах с умеренным климатом безошибочно сигнализирует о наступлении осени и затем зимы. Менее точно такая сигнализация осуществляется снижением средних температур воздуха, изменением его влажности, изменением состояния кормовых растений; объясняется это тем, что данные факторы среды изменчивы, часто отклоняются от нормы, поэтому не всегда надежны и при резком уклонении от средней нормы могут даже поставить вид в трудные условия жизни.
Признаки и типы диапаузы. Находящиеся в диапаузе личинки и взрослые особи обычно прекращают питание, их мышечная активность нередко снижается и насекомые часто становятся неподвижными или малоподвижными. У личинок прекращается рост и развитие, в яйце прекращается развитие зародыша. Все это свидетельствует о резком снижении обмена веществ в диапаузирующем организме.
Показателем снижения обмена веществ служит уменьшение скорости дыхания и падение потребления кислорода. Так, например, у зимующего жука-чернотелки Anatolica активные питающиеся особи осенью и весной потребляют в час 523-1236 мм3 кислорода на 1 г своего веса, тогда как зимующие диапаузирующие особи потребляют всего 115-437 мм3. Благодаря такому снижению уровня жизненных процессов диапауза обеспечивает очень экономное расходование запасных питательных веществ - жира, белков, гликогена. Вследствие этого продолжительность диапаузы может измеряться не только месяцами, но иногда достигает одного-двух и более лет. При высоком уровне обмена веществ это было бы невозможно.
Низкий уровень обмена веществ увеличивает устойчивость к воздействию неблагоприятных условий среды, повышает сопротивляемость к неблагоприятным внешним воздействиям. Так, диапауза увеличивает устойчивость к сухости воздуха и препятствует иссушению организма, что особенно имеет значение в странах с засушливым летом. Происходит также повышение холодостойкости, что особенно существенно для видов, живущих в странах с умеренным климатом и зимним сезоном. Наконец, отмечено повышение устойчивости к ядовитым веществам, что важно учитывать в практике защиты растений и в других случаях борьбы с вредными насекомыми.
Наступлению диапаузы у активных фаз развития и у куколок всегда предшествует период интенсивного питания и накопления запасных питательных веществ. Благодаря этому в организме насекомого, особенно в жировом теле, откладываются большие запасы жиро-белковых гранул и гликогена, что обеспечивает возможность диапаузирующей фазе длительное время оставаться без питания. Одновременно происходит уменьшение содержания воды в теле. Так, гусеницы яблонной плодожорки в период активной жизни в июле содержат в своем теле 11% жира и 72% воды (по отношению к сухому веществу тела), тогда как в августе к началу диапаузы содержание жира повышается до 18%, а содержание влаги снижается до 58%. Следовательно, обилие резервов и увеличенный объем жирового тела могут служить существенным показателем диапаузы или скорого ее наступления.
Формы, или типы, диапаузы разнообразны. Она может быть во всех фазах развития, но каждый вид обычно имеет только одну диапаузу, связанную с той или иной фазой развития. Так, в фазе яйца имеет место эмбриональная диапауза; она свойственна многим саранчовым, тутовому шелкопряду, дубовой листовертке и др.
Личиночная, или ларвальная, диапауза свойственна, например, среди бабочек боярышнице, златогузке, сосновому шелкопряду, яблонной плодожорке, луговому мотыльку и др.; все эти виды зимуют в фазе гусеницы. Куколочная, или пупальная, диапауза также обычна среди насекомых и наблюдается у капустной и репной белянок, капустной и хлопковой совок, капустной и свекловичной мух и др.; все эти виды зимуют в фазе куколки.
Во взрослой фазе имеет место имагинальная диапауза, которая характеризуется либо обратным развитием яичников, самки, т. е. рассасыванием яиц, либо возникает до половой зрелости; в обоих случаях размножение диапаузирующих имаго становится невозможным. Она свойственна ряду клопов, например клопам-черепашкам, колорадскому жуку и другим листоедам, свекловичному долгоносику, малярийному комару и др.
По времени года диапауза в странах с умеренным климатом может протекать в холодный период года либо в течение вегетационного периода; первая может быть названа зимней, вторая - летней диапаузой. В некоторых случаях диапауза продолжается более одного года, т. е. имеет затяжной характер; это двухлетняя и многолетняя диапаузы.
Следует также различать диапаузу обязательную, или облигатную, и необязательную, или факультативную. Облигатная диапауза характерна для моновольтинных видов и обеспечивает прохождение в течение года только одной генерации. Наблюдается она обычно в таких климатических зонах, где недостаток тепла или других условий не позволяет развиться двум поколениям. Поэтому моновольтинные виды широко представлены в умеренном климате, либо в теплом климате с длительным засушливым сезоном.
Примером могут служить саранчовые и клопы-черепашки. Так, большинство саранчовых в умеренном климате зимует в фазе яйца, но яйца откладываются часто еще летом или ранней осенью. После непродолжительного периода эмбрионального развития яйца саранчовых впадают в диапаузу, хотя внешние условия могут оставаться благоприятными для развития яйца. Особенно интересен пример с мароккской саранчой, населяющей южные жаркие и сухие области СССР и средиземноморские страны. Откладка яиц этим видом производится в начале лета, обычно в июне, а на юге ареала еще раньше; но начавшееся развитие приостанавливается диапаузой, и яйца без развития находятся всю оставшуюся часть лета, осень и зиму, т. е. до 8-9 месяцев в году. Лишь весной, в марте - апреле, наблюдается вылупление личинок. Как оказалось, выход из диапаузы, или реактивация, происходит при воздействии на диапаузирующие яйца пониженных температур.
Следовательно, эмбриональная диапауза у саранчовых препятствует быстрому отрождению личинок и устраняет появление второго поколения. Благодаря этому зимовка происходит в фазе яйца, а не в более требовательных к внешним условиям активных фазах развития - личинке или имаго; помимо того, для развития второго поколения может не хватить тепла и пищи. Для выхода из диапаузы, как уже указывалось, необходимо воздействием пониженных температур; это достигается охлаждением яиц в позднеосенний и зимний период, благодаря чему при наступлении тепла весной яйца уже способны к завершению своего развития.
Интересно отметить, что собранные ранней осенью яйцекладки саранчовых при переносе их в теплое помещение остаются в диапаузирующем состоянии длительное время; хотя яйца находятся, казалось, в благоприятной среде, отрождения личинок не происходит, что прежде всегда вызывало изумление. Однако путем воздействия на яйца пониженных температур можно добиться отрождения личинок даже осенью.
Таким образом, обязательная эмбриональная диапауза у саранчовых в умеренном климате обеспечивает синхронизацию их жизненного цикла с сезонной сменой внешних условий, т. е. создает возможность наиболее совершенного приспособления к жизни в таком климате.
У клопов-черепашек, серьезных вредителей хлебных злаков, имеется обязательная имагинальная диапауза. Питание взрослых клопов летом на созревающих хлебах обеспечивает накопление в организме богатых питательных резервов, сильное развитие жирового тела; это происходит в преддиапаузный период, после которого клопы отлетают в леса, скрываются под лесной подстилкой, впадают в диапаузу и в таком состоянии остаются зимовать. Весной клопы перелетают на всходы озимых и после дополнительного питания становятся половозрелыми и приступают к размножению. Следовательно, имагинальная диапауза обеспечивает возможность зимовки во взрослой фазе и создает условия для обильного весеннего питания на молодых растениях, богатых питательными веществами. Такой же тип диапаузы наблюдается у свекловичного долгоносика, земляных блошек и других моновольтинных видов, зимующих во взрослом состоянии.
У бабочек боярышницы и златогузки гусеницы отрождаются в конце лета, короткий период питаются листьями древесных пород, но к началу осени собираются в гнезда, прекращают питание, впадают в диапаузу еще задолго до листопада и наступления холодов и зимуют. Такая диапауза молодых гусениц обеспечивает перенос основного роста гусениц и их питания на весенний период, когда молодая листва содержит больше питательных веществ, нежели осенняя, закончившая рост листва.
Но обязательная диапауза становится биологически невыгодной в тех случаях, когда климатические условия и возможности питания допускают развитие двух и более поколений. В этих условиях возникает факультативная диапауза. Рост и развитие одного или нескольких поколений происходит без диапаузы, но при наступлении или приближении неблагоприятных условий включается механизм диапаузы; это обрывает развитие и обеспечивает подготовку соответствующей фазы к переживанию неблагоприятного периода - зимы или сухого сезона.
При факультативной диапаузе ведущую регулирующую роль приобретают внешние факторы. Эти факторы включают нервно-гуморальный механизм регуляции сезонного цикла развития насекомого и тем самым создают программу его жизни; развитие вначале идет беспрепятственно, но в том или ином поколении прекращается.
Как показали недавние исследования, среди которых особо следует отметить работы А. С. Данилевского и его сотрудников, ведущим регулирующим и программирующим фактором при факультативной диапаузе в большинстве случаев является продолжительность дня, т. е. длина фотопериода. При этом многие виды насекомых, развивающиеся с факультативной диапаузой, оказались длиннодневными: длинный летний день с продолжительностью светлого периода 14-17 и более часов благоприятствует беспрепятственному развитию, а укорочение дня к осени вызывает диапаузу (рис. 59). К таким видам, в частности, относятся капустная и репная белянки, луговой мотылек, яблонная и восточная плодожорки, озимая совка, малярийные комары, колорадский жук и др.
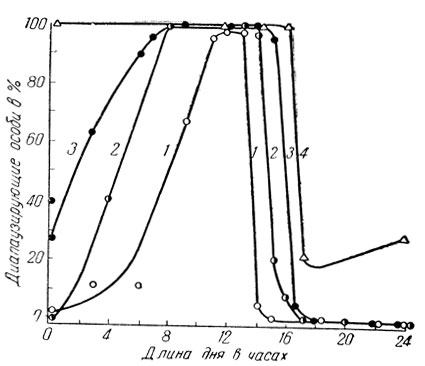
Рис. 59. Изменение числа диапаузирующих особей под влиянием фотопериода у длиннодневных видов (по Данилевскому): 1 - восточная плодожорка (при 24°С), 2 - капустная белянка (при 24°С), 3 - совка Acronycta rumicis (при 26°С), 4 - колорадский жук (при 24°С)
Короткодневные виды немногочисленны и нередко принадлежат к представителям более теплых климатических зон. Непрерывное развитие у них происходит только при коротком дне, менее 13-16 ч в сутки (рис. 60), а длинный день вызывает диапаузу. Сюда относится тутовый шелкопряд, а также цикадка Stenocranus minutus F. и немногие другие виды. Естественно, что есть виды и промежуточного типа; у них бездиапаузное развитие происходит только в пределах 18-20 ч света в сутки, тогда как при более коротком и более длинном дне гусеницы впадают в состояние диапаузы (рис. 61). Этот тип отмечен у ивового шелкопряда, златогузки и ее родичей, у северной формы стеблевого мотылька и др.
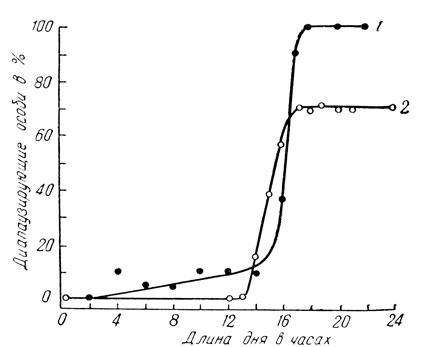
Рис. 60. Изменение числа диапаузирующих особей под влиянием фотопериода у короткодневных видов (по Данилевскому): 1 - цикадка Stenocranus minutus (при 20°С), 2 - тутовый шелкопряд (при 15°С)
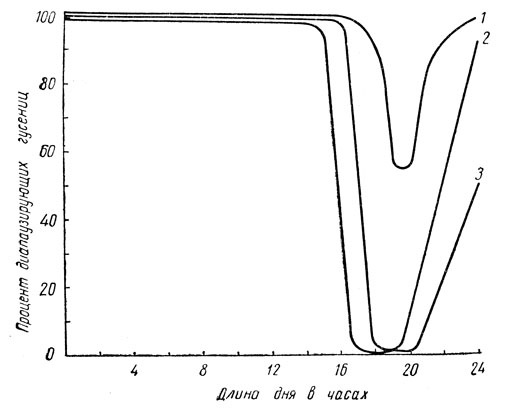
Рис. 61. Изменение числа диапаузирующих особей под влиянием фотопериода у видов промежуточного типа (по Данилевскому): 1 - ивовая волнянка (Leucoma salicis L.), 2 - желтогузка (Euproctis similis Fssl.), 3 - стеблевой мотылек (северная форма)
Факультативная диапауза под воздействием фотопериода у одних видов происходит в фазе яйца (тутовый шелкопряд), у других - в фазе личинки (яблонная и другие плодожорки, луговой мотылек, ивовый шелкопряд, златогузка, озимая совка и др.), куколки (капустная и репная белянки, капустная совка и др.) или имаго (колорадский жук, малярийный комар и др.).
Помимо зависимости от длины фотопериода, факультативная диапауза наблюдается и под воздействием других факторов среды. Так, гессенская муха в более влажном климате и при достаточно влажном лете развивается без диапаузы, дает за сезон 3-5 поколений. Но в степных районах, а также при сухом и жарком лете куколки весеннего поколения при недостатке контактной влажности вступают в диапаузу и в таком состоянии остаются до осени, т. е. до периода с осадками. В этих случаях развивается только два поколения - весеннее и осеннее, а летние поколения выпадают.
Выход из диапаузы. Важнейшей стороной диапаузы является необходимость своевременного выхода из нее, или реактивация. Одним из широко распространенных механизмов реактивации является воздействие пониженных температур; очень часто для выхода из диапаузы достаточно воздействия пониженных положительных температур, т. е. умеренного охлаждения диапаузирующей фазы. Так, охлаждение диапаузирующих куколок бабочек устраняет диапаузу и возобновляет развитие насекомого.
Как показало изучение гигантского шелкопряда Platysamia, охлаждение куколок стимулирует деятельность нейросекреторных клеток головного мозга, которые начинают выделять активационный мозговой гормон. Последний переносится через аксоны этих клеток к кардиальным телам и через них попадает в кровь, активизируя обмен веществ и проторакальные железы; эти последние выделяют гормон экдизон, который и способствует линьке куколки и выходу бабочки. Благодаря такому механизму зимующие куколки бабочек через охлаждение выводятся из диапаузы и весной дают бабочек. При отсутствии же охлаждения нейросекреторные клетки мозга остаются неактивными и куколка продолжает находиться в диапаузе.
Таким образом, короткий фотопериод осени способствует переходу куколок в диапаузу, а зимнее охлаждение реактивирует куколку и обеспечивает весной с наступлением тепла вылупление бабочек.
Воздействие пониженных температур вызывает также прекращение эмбриональной диапаузы у зимующих яиц саранчовых, тутового шелкопряда и других насекомых, обеспечивая в весеннее время завершение развития зародыша и вылупление личинок. Как было показано японскими исследователями С. Фукуда и К. Хазегава, нейросекреторные клетки бабочек тутового шелкопряда выделяют гормон диапаузы, который попадает в гемолимфу самки и через яичники передается яйцам; под воздействием этого гормона яйца впадают в диапаузу, развитие их тормозится. Охлаждение яиц, видимо, способствует разрушению этого гормона, что и обеспечивает выход из эмбриональной диапаузы.
Большое значение в реактивации имеют также высокие температуры. Так, у колорадского жука короткие осенние дни вызывают диапаузу имаго, диапаузирующие жуки зимуют. Выход их из диапаузы весной требует воздействия высоких температур; при 25°С выход жуков из почвы и последующее питание и размножение происходит за 2-3 месяца, но при 38°С на это достаточно только трех недель. У хлопковой совки короткий осенний фотопериод, воздействуя на гусениц, вызывает диапаузу куколки, которая и зимует; выход из диапаузы достигается воздействием на куколок высоких температур весеннего периода.
Высокая температура нередко также тормозит возникновение диапаузы даже при коротком дне осенью. Благодаря этому в более южных широтах эффект короткого дня ослабляется или снимается высокими температурами конца лета - осени, что обеспечивает здесь возможность развития дополнительных генераций. Сходным образом действует и жаркая осень, что также позволяет в этих условиях развиться дополнительному поколению. Следовательно, под воздействием избытка тепла происходит в сущности сдвиг порога фотопериодической реакции в сторону более короткого дня; другими словами, высокая температура среды делает диапаузу возможной при более коротком фотопериоде. Это явление отмечено у хлопковой совки, колорадского жука и других насекомых (рис. 62).
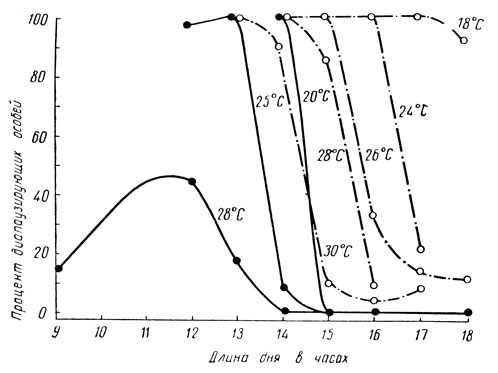
Рис. 62. Сдвиг фотопериодической реакции под воздействием различных температур (по Данилевскому): сплошные линии - хлопковая совка; прерывистые - колорадский жук
Реактивация может также происходить под воздействием повышенной влажности или, наоборот, сухости. Выше уже отмечалась необходимость контактной влажности для выхода из диапаузы коконов гессенской мухи. С другой стороны, диапаузирующие яйца южноафриканской бурой саранчи (Locustana pardalina Walk.) лучше всего реактивируются после полуторамесячной выдержки в сухой среде и последующего увлажнения; тогда дружное отрождение происходит через 8 дней.
Таким образом, механизм реактивации разнообразен и определяется теми условиями среды, в которых существуют те или иные виды насекомых.
Многообразие типов диапаузы у вида. В последние годы было также установлено, что один и тот же вид насекомого может иметь несколько форм диапаузы. Так, для зимней пяденицы характерны две формы диапаузы - эмбриональная и куколочная. Зимовка у этого вида происходит в фазе яйца и эмбриональная диапауза обеспечивает ее успех; куколочная же диапауза происходит летом и приводит к тому, что выход бабочек из куколок оттягивается до поздней осени.
У колорадского жука отмечено даже несколько форм диапаузы: зимняя, летняя и затяжная (многолетняя) диапауза жуков. Сходное явление отмечено и у китайского дубового шелкопряда: неустойчивая кратковременная летняя диапауза, зимняя диапауза и затяжная диапауза.
Многообразие форм физиологического покоя обеспечивает наиболее успешное переживание видом неблагоприятных условий, создающихся в разные сезоны года. На эту сторону диапаузы впервые было обращено внимание Р. С. Ушатинской, которая справедливо усматривает во множественности форм диапаузы вида совершенный приспособительный эколого-физиологический механизм, увеличивающий экологическую пластичность вида. Возникновение этого механизма является одним из путей биологического прогресса видов, обеспечивая им высокую численность особей и расширение ареала.
Заключение. В целом факторы среды, вызывающие диапаузу, и факторы среды, выводящие организм из диапаузы, обеспечивают наиболее выгодную для вида синхронизацию своего жизненного цикла с местными сезонными явлениями и через посредство нейро-гуморальной регуляции создают этот жизненный цикл. При этом роль эндокринной системы сводится, видимо, к тому, что отдельные ее элементы тормозят развитие, т. е. способствуют переходу в диапаузу и становятся, следовательно, ингибиторами роста; другие же компоненты эндокринной системы играют роль активаторов, или стимуляторов роста, и выводят организм из состояния диапаузы.
Следовательно, диапаузу можно представить себе как результат антагонистически действующих гормонов, количественное соотношение между которыми программируется в организме воздействиями внешней среды.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'