Биология размножения
Размножение насекомых имеет ряд особенностей, которые составляют существенную сторону их биологии и требуют своего рассмотрения. К этим особенностям относятся способы размножения, дополнительное питание, встреча полов и оплодотворение, половая продуктивность.
Способы размножения. Большинство насекомых является яйцекладущими организмами; вылупление из яйца происходит уже после его откладки. Помимо того, размножение у большинства насекомых сопровождается спариванием и оплодотворением, т. е. связано с участием обоих полов; поэтому оно может быть названо обоеполым, или бисексуальным, но также обозначается как гамогенетическое размножение. Однако насекомым свойственны и другие способы размножения - живорождение, партеногенез, педогенез и полиэмбриония.
Живорождение состоит в том, что эмбриональное развитие завершается в теле матери; поэтому вместо откладки яиц на свет производятся личинки или даже куколки. Оно наиболее характерно для девственных самок тлей, а также для некоторых мух, как кровососки и овода; но наблюдается и у немногих видов других отрядов. Наиболее частная форма - обычное живорождение, неудачно называемое также яйцеживорождением. Оно состоит в том, что вылупление личинки происходит в яйцевых трубках или в процессе прохождения по яйцеводам; никакого специального органа для развития зародыша нет. Наблюдается у некоторых таракановых, кокцид, трипсов, жуков и мух.
Более специализованной формой является ложноплацентное живорождение, для которого характерно образование сходной с плацентой структуры в маткообразном расширении непарного яйцевода; через эту ложную плаценту происходит питание яиц, которые в большинстве случаев лишены хориона. Этот тип свойствен тлям, а также некоторым паразитам теплокровных, в частности гемимеридам (Hemimerida). Можно назвать также куклорождение, свойственное мухам-кровососкам и мухе це-це; единственная личинка развивается в маткообразном расширении непарного яйцевода матери, где питается через рот за счет выделений придаточных желез. Самка откладывает лишь вполне созревшую готовую к окуклению личинку, которая вскоре превращается в куколку.
Партеногенез, или девственное размножение, характеризуется отсутствием оплодотворения и наблюдается как у яйцекладущих, так и живородящих насекомых, а также и при педогенезе. Он наблюдается у отдельных представителей почти всех отрядов насекомых и не найден лишь у стрекоз и клопов.
В биологическом отношении партеногенез разнообразен и представлен рядом форм. Так, из неоплодотворенных яиц могут развиваться либо только самцы (арренотокия) или самки (телитокия), либо оба пола (амфитокия). Помимо того, партеногенез может быть факультативным, постоянным и циклическим. Однако цитологически все это многообразие может быть сведено к двум основным типам партеногенеза - генеративному и соматическому. Первый характеризуется гаплоидным, т. е. половинным числом хромосом в соматических клетках зародыша, а второй - нормальным диплоидным числом или даже полиплоидным.
Объясняется это следующим. Известно, что незрелые половые клетки вступают в мейоз, т. е. деление с редукцией числа хромосом; этим половые клетки отличаются от соматических, которые имеют двойной, диплоидный, набор хромосом. Такое деление является нормой и еще сохраняется при генеративном партеногенезе; благодаря этому зародыш и имеет уменьшенное вдвое против нормы, т. е. гаплоидное число хромосом. Естественно, что такое состояние организма - отклонение от нормы, и поэтому возникают механизмы восстановления у потомков диплоидного набора. Это и происходит при соматическом партеногенезе путем выпадения редукционного деления или путем слияния образовавшихся гаплоидных ядер.
У ряда видов партеногенез носит частичный характер, т. е. наблюдается непостоянно, возникая лишь при некоторых внешних воздействиях или при определенном физиологическом состоянии яйцекладущей самки. Такой партеногенез называют факультативным. Одной из наиболее обычных форм факультативного партеногенеза как раз и служит генеративный партеногенез; ткани зародыша имеют в клетках, как уже указывалось выше, гаплоидный набор хромосом. При этой форме из неоплодотворенных яиц развиваются, как правило, самцы, т. е. размножение приобретает характер арренотокического партеногенеза. Это характерно для медоносной пчелы, ряда наездников и других паразитических перепончатокрылых, некоторых кокцид, алейродид, трипсов и др.
Другая форма партеногенеза - факультативная телитокия, при которой из оплодотворенных яиц развиваются самцы и самки, а из неоплодотворенных - только самки. Образование яйца сопровождается редукционным делением; но при этом ядро в неоплодотворенном яйце не остается гаплоидным, а становится диплоидным или даже полиплоидным вследствие слияния образующихся гаплоидных ядер. Следовательно, это уже пример соматического партеногенеза. Он известен у некоторых палочников, саранчовых - тетригид (Tetrigidae), кокцид, пилильщиков и др. и характеризуется обычно наличием в популяциях не только самок, но и самцов.
Особую форму соматического партеногенеза составляет постоянная телитокия, при которой самцы отсутствуют или редки и из неоплодотворенных яиц развиваются только самки. Процесс развития яйца протекает без редукции хромосом, поэтому в ядре поддерживается диплоидное состояние, но нередки случаи и полиплоидии. Этот вид партеногенеза широко распространен и известен у некоторых кузнечиков, тараканов, кокцид, жуков, в частности долгоносиков, у пилильщиков и др.
Наконец, у тлей, орехотворок и некоторых мух-галлиц наблюдается циклический партеногенез. Он состоит в правильном чередовании обоеполых и девственных поколений, т. е. представляет собою одну из форм чередования поколений, или гетерогонии. При этом потомство неоплодотворенной самки может длительное время состоять только из самок, но в конце концов всегда наступает также арренотокия и даже амфитокия; благодаря этому восстанавливается обоеполое размножение. Подробнее цикл развития тлей рассмотрен в разделе систематики и классификации.
В целом партеногенез играет существенную роль в жизни соответствующих видов насекомых и несомненно имеет приспособительное значение. Благодаря партеногенезу вдвое увеличивается потенциал размножения, так как вместо двух особей разных полов репродукцией занят только один пол; следовательно, приобретение способности к партеногенезу равноценно удвоению половой продукции, если соотношение полов у исходного вида или поколения составляло 1:1. Партеногенез также увеличивает шансы на расселение вида; поэтому девственно размножающиеся виды часто имеют обширный географический ареал. Факультативный партеногенез позволяет также многим видам преодолеть воздействие неблагоприятных условий среды.
Помимо охарактеризованного выше естественного партеногенеза возможен и искусственный партеногенез, т. е. искусственная активация неоплодотворенных яиц различными внешними воздействиями. Впервые А. А. Тихомиров еще в 1886 г. добился развития неоплодотворенных яиц у тутового шелкопряда, а в наше время Б. Л. Астауров разработал методы регуляции пола у данного шелкопряда. Благодаря этому создались возможности повышения продуктивности шелководства путем искусственного получения самцов, коконы которых крупнее и ценнее, чем у самок. Нет сомнения, что разработка методов искусственного партеногенеза и регуляции пола у других полезных насекомых, например у энтомофагов, может также иметь значение для практики.
Педогенез, или детское размножение, представляет собою размножение на фазе личинки. Впервые он был открыт русским исследователем Н. Вагнером в 1862 г. у личинок галлиц (Cecidomyidae) из рода Miastor, но затем обнаружен у других галлиц, а также у отдельных видов жуков и клопов. В яичниках личинки происходит партеногенетическое развитие яиц, из которых возникают личинки, поедающие тело материнской личинки при выходе из нее наружу; личинки нового поколения в свою очередь размножаются педогенетически и так происходит развитие нескольких поколений, которое в конце концов сменяется серией обоеполых поколений взрослых особей. Таким образом, педогенез является по существу одной из форм партеногенеза и одной из форм гетерогонии, т. е. смены поколений.
Полиэмбриония, или многозародышевое размножение, представляет собою размножение на фазе яйца, свойственно некоторым паразитическим перепончатокрылым и веерокрылым и впервые было открыто французским исследователем П. Маршалем в 1898 г. При полиэмбрионии в тело хозяина откладывается одно яйцо, которое затем путем сложных преобразований разрастается в длинную цепочку из многих десятков яиц; каждое яйцо дает свою личинку, которые затем дорастают до куколки и дают взрослых особей. Яйца размножающихся с помощью полиэмбрионии насекомых мелки и бедны питательным материалом; но за счет использования веществ хозяина они приобретают возможность к резкому увеличению своей массы, а последующие фазы - возможность дальнейшего развития и достижения взрослого состояния.
Полиэмбриония является, следовательно, выгодным приспособлением паразитических насекомых и обеспечивает резкое увеличение численности потомков при ничтожном расходе живого вещества матери. Она по существу должна рассматриваться как бесполое размножение, чем резко отличается от обычного гамогенетического, а также партеногенетического и педогенетического размножения, которые являются уже половыми его формами.
Дополнительное питание. Способность к размножению проявляется у одних насекомых вскоре после окрыления, у других - через более или менее продолжительный срок. Происходит это вследствие неодинаковой половозрелости окрылившихся особей.
Некоторые уже сразу после превращения в имаго имеют вполне созревшие половые продукты и способны к спариванию и яйцекладке, не нуждаясь при этом в питании. Таковы поденки, многие бабочки (коконопряды, шелкопряды, волнянки и др.), ряд двукрылых (комары-долгоножки, овода, гессенская муха и другие галлицы) и некоторые другие насекомые. Все они имеют недоразвитые ротовые органы и не способны к приему пищи. Жизнь имаго у них непродолжительна и часто ограничивается немногими днями или даже часами, необходимыми лишь для спаривания и откладки яиц; после выполнения этой функции насекомое умирает. Следовательно, у таких насекомых разделение биологических функций между личинкой и взрослой фазой достигло наибольшей степени.
Однако в большинстве случаев окрылившиеся особи неполовозрелы, нуждаются в более или менее продолжительном питании и только после этого созревают для размножения. Питание в имагинальной фазе, необходимое для созревания половых продуктов, называется дополнительным. Период дополнительного питания может быть различной продолжительности - в зависимости от вида насекомого, условий жизни взрослой фазы, а иногда и личинки. Поэтому половозрелое состояние может наступить в одних случаях через 5-10 дней, в других - через месяц и более.
Дополнительное питание характерно и обязательно прежде всего для зимующих во взрослом состоянии видов, так как зимовка сопряжена с растратой питательных резервов жирового тела. Такие виды весной после зимовки всегда нуждаются в дополнительном питании, в связи с чем растениеядные виды в это время могут сильно вредить. Этим объясняется сильная вредоносность в весенний период таких вредителей, как клопы-черепашки, свекловичный долгоносик, клубеньковые долгоносики, земляные блошки, майские хрущи и др. Перезимовавшие самки комаров-кровососов (малярийный комар и другие виды) для полового созревания также нуждаются в приеме пищи и без кровососания остаются неполовозрелыми даже если они перед зимовкой приняли порцию крови.
Дополнительное питание возможно и у видов, не зимующих во взрослой фазе - таких, как большинство саранчовых, многие бабочки и пр. Иногда дополнительное питание может быть необязательным и происходит в тех случаях, когда личинки жили в неблагоприятных условиях, недостаточно питались и не накопили необходимых резервов; тогда взрослые особи оказываются неполовозрелыми и нуждаются в дополнительном питании. Если же жизнь и питание личинок протекали в благоприятных условиях, необходимость в дополнительном питании может отпасть, что и наблюдается, например, у лугового мотылька и озимой совки.
Встреча полов и оплодотворение. Важным условием размножения является встреча полов, спаривание и оплодотворение.
Встреча самца и самки обеспечивается применением различных опознавательных видовых сигналов - звуковых, зрительных, химических. Звуковые сигналы часто имеют характер стрекотания и особенно типичны для прямокрылых (саранчовые, сверчки, кузнечики) и певчих цикад. Каждый вид имеет свой, только ему свойственный набор подаваемых звуков, легко распознаваемый особями этого вида; записанное на магнитофон стрекотание привлекает особей только данного вида. Такие насекомые имеют свои звуковые органы, причем в одних случаях способностью издавать звук обладают оба пола (певчие цикады, многие саранчовые, некоторые кузнечики и пр.), в других - только самец (большинство кузнечиков и сверчки). Строение звуковых органов и способы издавать звуки весьма разнообразны и описываются в разделе "Систематика и классификация" при характеристике отдельных систематических групп.
Зрительные опознавательные сигналы часто проявляются в яркой окраске тех или иных частей или всего тела в целом. Так, дневные бабочки часто имеют ярко окрашенную верхнюю сторону крыльев и могут опознаваться не только при посадке, когда они держат крылья плоско на субстрате и демонстрируют окраску верхней стороны, но даже и во время полета. Многие саранчовые имеют ярко окрашенные задние голени и бедра, у других ярко окрашены задние крылья; при взлете и раскрывании крыльев возникает резкий цветовой сигнал, привлекающий особей своего вида. У ряда видов бабочек с внешне одинаково окрашенными крыльями обнаружены резкие половые отличия в интенсивности ультрафиолетового излучения; следовательно, крылья у таких бабочек могут служить световым сигналом, скрытым от человеческого глаза.
Химические видовые сигналы, очевидно, широко распространены у насекомых и частью рассмотрены раньше (стр. 85). Благодаря выделению привлекающего вещества - полового аттрактанта - обеспечивается встреча полов.
Оплодотворение составляет важнейшую сторону биологии размножения, отличается у насекомых большим разнообразием форм и специфично у тех или иных систематических групп. Закономерности эволюционного развития способов оплодотворения и связь их со средой недавно были вскрыты М. С. Гиляровым.
У тех низших насекомых (Apterygota), которые еще сохранили связь с влажной средой и живут в почве или гнилой древесине (двухвостки, часть подур), оплодотворение не сопровождается контактом самца и самки, т. е. происходит без спаривания. Самцы рассеивают капельки спермы или сперматофоры в местах обитания вида, притом часто даже в отсутствие самки. Оплодотворение происходит при встрече самки с этими порциями семени путем захватывания его половым отверстием самки. Следовательно, сперма первоначально находится во внешней среде и лишь затем попадает без участия самца в половые пути самки, т. е. во внутреннюю среду. Таким образом, это оплодотворение является наружно-внутренним без спаривания и ведет свое происхождение от наружного неизбирательного оплодотворения, свойственного водным организмам, когда сперма просто выводилась в водную среду (низшие водные членистоногие, водные кольчатые черви). Наружно-внутреннее оплодотворение без спаривания, очевидно, является у насекомых наиболее примитивной его формой и могло возникнуть лишь в среде, насыщенной влагой; поэтому оно и характерно для влаголюбивых низших насекомых, живущих в почве, гнилой древесине и в сходной среде.
В условиях воздушной среды такое наружно-внутреннее оплодотворение без спаривания становится невозможным, так как поведет к быстрому высыханию и гибели открыто рассеянных порций семени. В связи с этим при переходе к наземному и воздушному образу жизни возникает необходимость в спаривании и в сокращении периода пребывания семени во внешней среде.
Первоначально оплодотворение в такой среде еще имеет наружно-внутренний характер, но самец уже сближается с самкой, а порции откладываемого семени быстро подхватываются самкой в одних случаях с помощью самца, в других - без его участия. Таким образом, здесь уже наблюдаются зачатки спаривания, сокращается срок пребывания семени во внешней среде, но нет еще настоящей копуляции, т. е. тесного сближения обоих полов при спаривании. Этот тип оплодотворения наблюдается у тех низших насекомых, которые живут в менее влажной, чем почва, среде, но еще связаны с ее поверхностью, т. е. живут в подстилке, под камнями, в толще густого травостоя и пр. Сюда относятся щетинохвостки и представители слитнобрюхих подур.
У высших, или крылатых, насекомых (Pterygota) в связи с переходом их взрослой фазы к жизни в воздушной среде, т. е. в условиях дефицита влаги, наружно-внутреннее оплодотворение заменяется внутренним и сопровождается копуляцией. В низших отрядах и подотрядах крылатых оплодотворение сопровождается выделением сперматофоры, которая сразу подхватывается половыми придатками самки (рис. 57). Так происходит оплодотворение у таракановых, богомоловых, длинноусых прямокрылых, сетчатокрылых. Но в высших группах крылатых сперматофора либо вводится непосредственно в половые пути самки (саранчовые, часть клопов, жуков, ручейников, перепончатокрылых, некоторые низшие двукрылые, чешуекрылые), либо сперматофоры полностью утрачиваются и в половые пути самки при копуляции вводится одна сперма (термиты, клопы, трипсы, скорпионовы мухи, часть жуков, ручейников, перепончатокрылых, большинство двукрылых). В обоих случаях для введения половой продукции у самца вырабатывается соответствующий аппарат - копулятивный орган, или эдеагус.
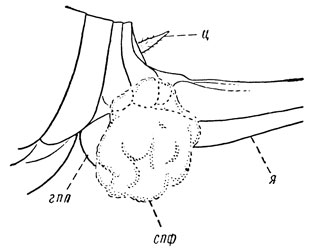
Рис. 57. Конец брюшка самки кузнечика со сперматофорой (по Болдыреву): ц - церк, спф - сперматофора, гпл - генитальная пластинка, я - яйцеклад (часть)
Описанная выше эволюция типов оплодотворения у насекомых возникла, по М. С. Гилярову, в результате смены сред обитания, именно в связи с выходом насекомых в течение их филогенеза из почвы в воздушную среду. А в целом она является еще одним доказательством развиваемой Гиляровым концепции о происхождении наземных членистоногих, включая и насекомых, от водных предков через посредство почвы как промежуточной среды, совмещающей свойства водной и воздушной сред жизни.
После оплодотворения самка вскоре приступает к откладке яиц (или живорождению). В ряде случаев наблюдается повторное спаривание и повторное размножение, особенно если продолжительность жизни имаго исчисляется не немногими днями (как, например, у поденок, шелкопрядов и пр.), а более длинным периодом; у отдельных видов (некоторые долгоносики и чернотелки среди жуков) она может достигать даже нескольких лет.
Плодовитость. Плодовитость насекомых нередко очень велика, но не является величиной постоянной. Определяется она двумя факторами: наследственными свойствами вида (строение и величина яичников), т. е. его потенциалом размножения, и воздействиями внешней среды. При благоприятных внешних условиях потенциал размножения реализуется в наибольшей степени. Так, самка хлопковой совки может отложить за свою жизнь до 2700 яиц, озимая совка - до 1200-1800 яиц, луговой мотылек - до 800, гессенская муха - до 500, хлебные пилильщики - до 50. В отдельных случаях плодовитость может достигать поразительных размеров, особенно у общественных насекомых. Например, самка (матка) медоносной пчелы откладывает в день до 3 тыс. яиц, а у термитов - даже до 30 тыс.
Однако потенциал размножения далеко не всегда реализуется полностью и плодовитость может быть сильно снижена под влиянием неблагоприятных условий внешней среды - погоды, условий питания и пр. Следовательно, в конечном счете плодовитость контролируется экологическими факторами, что и будет более подробно рассмотрено в разделе "Экология".
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'