Органы чувств
Органы чувств являются посредниками между внешней средой и организмом, т. е. своего рода органами информации о состоянии внешних условий жизни. В соответствии с этими внешними стимулами, или раздражителями, насекомое и совершает те или иные действия; из этих действий складывается в целом поведение насекомого.
По аналогии с органами чувств высших животных и человека у насекомых обычно различают органы осязания, слуха, обоняния, вкуса и зрения. Однако это деление антропоморфично и более целесообразно различать у насекомых следующие чувства с их рецепторами: механическое чувство, слух, химическое чувство, гигротермическое чувство и зрение.
Основу органов чувств составляют их нервно-чувствительные единицы - сенсиллы (рис. 35), которые в типичных случаях состоят из двух компонентов: кожной структуры и прилегающих к ней нервных чувствительных клеток, нередко в числе одной в каждой сенсилле. Сенсиллы в зависимости от особенностей воздействий и восприятия раздражений устроены разнообразно, но в целом условно можно различать два их основных типа - непогруженные и погруженные сенсиллы; первые выступают над поверхностью кожи в виде волоска, щетинки, конуса или другого образования, вторые же располагаются под кутикулой или в коже.
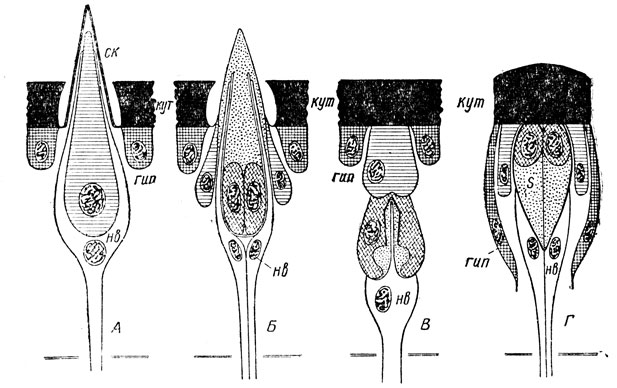
Рис. 35. Схема различных типов сенсилл: А - непогруженная осязательная; Б - непогруженная химическая; В - погруженная слуховая, Г - погруженная зрительная (по Берлезе); кут - кутикула, гип - гиподерма, ск - колпачок, нв - нервная клетка
Механическое чувство. Механическое чувство представлено механорецепторами, которые воспринимают различные механические воздействия и очень часто представлены всего лишь одной клеткой. Сюда относятся осязательные рецепторы, а также чувствительные структуры, воспринимающие сотрясение, положение тела, его равновесие и пр.
Осязательные, или тактильные, рецепторы разбросаны по всему телу в виде простых непогруженных сенсилл с сочлененным чувствительным волоском. Изменение положения волоска при прикосновении твердого предмета или под воздействием движения воды или воздуха передается чувствительной клетке, где и возникает возбуждение, передаваемое по ее отросткам в нервный центр.
Другой вид механорецепторов составляют колоколовидные сенсиллы, отличающиеся от предыдущих отсутствием чувствительного волоска и погруженные в кожу. Их рецепторная поверхность имеет вид кутикулярного колпачка или колокола и находится на поверхности кутикулы; к этому колпачку снизу подходит стержневидный концевой отросток чувствительной клетки - штифт, или сколопс. Колоколовидные сенсиллы располагаются на различных частях тела: на крыльях, церках, ногах, щупальцах и пр.; они, видимо, воспринимают сотрясения тела, а также механические изменения покровов, например сгибы, натяжения и др.
К числу механорецепторов следует отнести и хордотональные органы, нередко рассматриваемые как органы слуха. Однако у насекомых трудно провести границу между восприятием механических сотрясений и слуховым чувством, воспринимающим давление воздуха. Основываясь на морфологических особенностях, хордотональные органы целесообразнее рассмотреть среди механорецепторов. Их нейроны, подобно нейронам колоколовидных сенсилл, заканчиваются стержневидным штифтом. Однако хордотональные органы представляют собой серию особых сенсилл, натянутых между двумя участками кутикулы. Хордотональные сенсиллы называются сколопофорами, или сколопидиями, и каждая из них состоит из трех клеток: чувствительного нейрона, колпачковой и обкладочной (рис. 36).
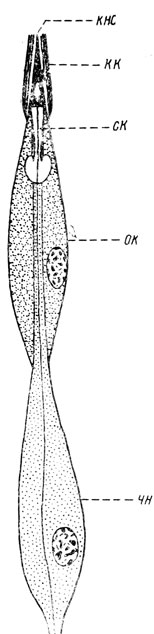
Рис. 36. Схема сколопофора личинки жука-усача (по Иммсу): кнс - концевая нить сколопса, кк - колпачковая клетка, oк - обкладочная клетка, ск - сколопс, чн - чувствительный нейрон
Чувствительный нейрон имеет, как уже указывалось, стержневидный концевой отросток - штифт, или сколопс; последний удален от кутикулы, но связан с ней при помощи концевой нити. Вершинная часть штифта прикрыта удлиненной колпачковой клеткой, которая противоположным своим концом прикреплена к кутикуле; эта клетка пронизана концевой нитью штифта. Обкладочная клетка расположена в срединной части сколопофора и охватывает часть нервной клетки и значительную часть штифта. Характерно, что кутикула в месте присоединения к ней такой хордотональной сенсиллы не имеет снаружи каких-либо отклонений от обычного строения и лишь иногда снабжена небольшой ямкой или слабым выступом.
Хордотональные органы, как совокупность описанных сенсилл, располагаются на различных частях тела - на брюшке, усиках, ногах, крыльях и пр. Распределены они обычно симметрично и метамерно и встречаются в большом числе - десятками пар. Из-за сходства сколопофор этих органов со сколопофорами тимпанальных органов слуха хордотональным органам прежде приписывалась слуховая функция. Однако выше уже указывалось, что кутикула хордотональных сенсилл не обнаруживает особых изменений и тем более каких-либо утончений или превращения в барабанную перепонку. Поэтому в настоящее время хордотональным органам приписывают восприятие механических напряжений и их изменения, изменений внутреннего давления, механических вибраций, видимо включая частично и некоторые звуковые воздействия. Метамерность и симметричность их расположения указывают также на то, что они играют роль проприорецепторов - органов, регулирующих положение тела или его частей, особенно во время полета.
Особой специализованной формой хордотональных органов является Джонстонов орган. Он располагается на втором членике усиков и считается органом, воспринимающим движение и сотрясение воздуха или воды, а также контакт с твердым субстратом. У настоящих комаров (сем. Culicidae) этот орган устроен особенно сложно и выполняет слуховую функцию.
Слух. Слух развит далеко не у всех насекомых. У прямокрылых (саранчовые, кузнечики, сверчки), певчих цикад, некоторых клопов и ряда чешуекрылых слуховые рецепторы представлены тимпанальными органами. Особенно хорошо развиты они у тех насекомых, которые обладают звуковыми органами и, следовательно, могут издавать звук в виде стрекотания или пения; таковыми являются прямокрылые и певчие цикады.
Анатомически тимпанальные органы представляют собой скопление сколопофоров, которые связаны с утонченными в виде барабанной перепонки (по латыни tympanum) участками кутикулы. В целом они устроены очень сложно и здесь не рассматриваются. Снаружи они особенно хорошо выражены у прямокрылых. У саранчовых по бокам I сегмента брюшка находится затянутое тонкой прозрачной барабанной перепонкой полулунное или неправильно овальное отверстие с утолщенными краями (рис. 1). Кузнечики и сверчки имеют отверстия тимпанального органа на голенях передних ног - в виде затянутого перепонкой овала или в виде узкой щели (рис. 37).
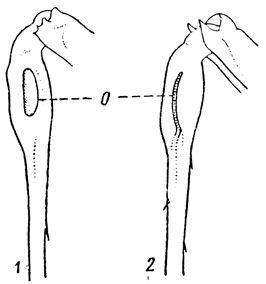
Рис. 37. Передняя голень кузнечиков с отверстием тимпанального органа (О) (по Бей-Биенко): 1 - открытое овальное, 2 - щелевидное
У певчих цикад пара тимпанальных органов располагается у основания брюшка и связана со звуковыми органами. У чешуекрылых и клопов они развиты слабее и располагаются в разных местах. Так, у дневных бабочек тимпанальные органы находятся на вздутом основании передних крыльев, у совок и других групп - между грудью и брюшком и пр.
Функцию органа слуха у комаров выполняет также Джонстонов орган, уже упоминавшийся выше при описании механорецепторов. Помимо того, на церках у некоторых тараканов и прямокрылых и на теле гусениц многих бабочек располагаются слуховые волоски, обладающие способностью улавливать звуковые волны.
Значение органов слуха у насекомых двоякое. С одной стороны, с их помощью воспринимаются сигналы, идущие от особей своего вида, что обеспечивает связь полов; следовательно, улавливание звука - стрекотания или пения - необходимо рассматривать как одну из форм локации полового сигнала. С несомненностью это доказано у прямокрылых. С другой стороны, органы слуха улавливают и иные звуки, идущие из внешней среды; в частности, некоторые насекомые реагируют на свисток или резкий звук, некоторые паразитические виды, возможно, разыскивают жертву по издаваемым ею звукам и пр.
Диапазон воспринимаемых частот в целом у насекомых широк, охватывая области от инфразвука (например, 8 колебаний в секунду) до ультразвука (от 40 тыс. колебаний и более в секунду). Однако каждый отдельный вид имеет свойственный только ему диапазон улавливаемой частоты колебаний. Например, слуховые волоски некоторых гусениц бабочек воспринимают звуковые волны в диапазоне 32-1024 колебаний в секунду, тогда как у египетской кобылки (Anacridium aegyptium L.) тимпанальные органы способны воспринимать звук в пределах 2480-40000 колебаний в секунду.
Химическое чувство. Химическое чувство служит для восприятия химизма среды, именно запаха и вкуса, и представлено хеморецепторами; ввиду того, что в ряде случаев уловить отличия между обонятельными и вкусовыми рецепторами трудно, целесообразно говорить об органах химического чувства в целом. Однако физиологические различия между этими рецепторами заключаются в том, что обоняние воспринимает и анализирует газообразную среду с низкой концентрацией вещества, а вкус - жидкую среду с высокой его концентрацией.
Сенсиллы хеморецепторов разнообразны по строению и могут быть непогруженными - в виде волосков или конусов, либо в виде пластинок или погруженных конусов. Среди них обонятельную функцию несут обильно развитые на усиках пластинчатые и конусовидные (рис. 35, Б) сенсиллы; например, рабочая пчела имеет около 6 тыс. пластинчатых сенсилл на каждом усике. Иногда эти сенсиллы собраны в ямки, например на третьем членике усиков у мух. Количество обонятельных сенсилл зависит от образа жизни вида и способов и характера добывания пищи. У самцов их обычно больше, чем у самок, и это связывается с активным розыском последних. Удаление усиков у пчелы лишает ее обоняния и способности розыска пищевого субстрата.
Обоняние служит насекомым разнообразно - для разыскания пола, распознавания особей своего вида, для отыскания пищи и мест откладки яиц. Несомненно, многие насекомые выделяют привлекающие вещества - половые аттрактанты. Известно, например, что неоплодотворенные самки ряда бабочек способны привлекать самцов с расстояния в 3-9 км; привлекающий секрет, выделяется особыми железами на брюшке. Оплодотворенная самка или самка с удаленными железами уже не привлекает самцов. Удивительна в этих фактах необычайная чувствительность обонятельных сенсилл; они способны улавливать половой аттрактант на большом расстоянии и при ничтожно малой его концентрации - вероятно, исчисляемой немногими молекулами на кубометр воздуха.
Издаваемый клопами характерный запах связан с веществом, которое, видимо, играет разную роль: видового, или полового, аттрактанта в одних случаях и отпугивающего вещества, или репеллента - в других. Во всяком случае установлено, что клоп-черепашка (Eurygaster integriceps Put.) усиленно выделяет пахучее вещество в период образования массовых скоплений; очевидно, запах в этом случае играет роль распознавательного видового сигнала и хорошо улавливается обонятельными рецепторами.
Способность воспринимать распознавательные запахи свойственна также общественным насекомым, таким, как пчелы и муравьи; они легко узнают особей своей колонии и нападают на особей из других колоний, хотя и относящихся к своему же виду.
Обоняние несомненно играет выдающуюся роль в отыскании и распознавании пищи. Так, пчелы способны различать и запоминать разнообразные цветочные запахи, играющие роль пищевых сигналов. Растительноядные насекомые нередко обладают резко выраженной избирательностью к растениям, т. е. питаются строго определенными видами или родственными группами растений - представителями тех или иных родов или семейств и пр. И вот такие насекомые легко находят свои кормовые растения. Установлено, что эти растения распознаются по запаху и вкусу имеющихся в их тканях специфических веществ.
Известно, что родственные растения имеют общие биохимические свойства, т. е. содержат в своих тканях и органах сходные или одинаковые химические вещества - глюкозиды, жирные или эфирные масла, органические кислоты, белки и пр. Например, крестоцветные и родственные им каперцовые и резедовые содержат глюкозиды, дающие при расщеплении горчичные масла; розоцветные содержат цианогенные глюкозиды, например амигдалин, дающие при расщеплении цианосодержащие вещества, и т. д. Эти соединения очень специфичны, легко улавливаются и распознаются хеморецепторами насекомых и играют роль своеобразных сигнальных веществ - фагостимуляторов, по которым насекомые разыскивают необходимые им кормовые растения. Поэтому питание, например, огородных блошек (род Phyllotreta) не только на крестоцветных, но и на резедовых и каперцовых находит свое объяснение в сходстве сигнальных веществ этих растений.
Сигнальные вещества нередко играют существенную роль и при розыске подходящего субстрата для яйцекладки. Так, капустная белянка (Pieris brassicae L.), привлекаемая горчичными глюкозидами, откладывает яйца не только на крестоцветные, но и на каперцовые, хотя для самой бабочки эти растения, если они не цветут, не имеют пищевого значения. Навозные и падальные жуки и мухи быстро обнаруживают появившийся помет или падаль и используют их для откладки яиц; как оказалось, эти насекомые легко улавливают и привлекаются запахами аммиака, индола, скатола и других специфических соединений, образующихся при бактериальном разложении органических веществ.
Приспособление к восприятию хеморецепторами тех или иных сигнальных веществ (фагостимуляторов, аттрактантов), очевидно, возникает в процессе эволюционного развития насекомых и служит надежным средством в розыске не только пищи, но и субстрата при откладке яиц, особей своего пола и пр.
Вкус играет более специфическую роль, нежели обоняние, так как служит лишь для распознавания пищи. Установлено, что насекомые различают четыре основных вкуса - сладкий, горький, кислый и соленый. Большинство Сахаров, такие, как глюкоза, фруктоза, мальтоза и др., привлекают пчел и мух даже при сравнительно низкой концентрации; другие сахара, как галактоза, манноза и пр. распознаются лишь при высокой концентрации, причем пчелы отвергают их. Очень чувствительны к сахарам некоторые бабочки, отличающие от чистой воды раствор сахара с ничтожной концентрацией - 0,0027%. Известно также, что муравьи отбирают кристаллы обычного сахара из смеси его с сахарином и не трогают последний.
Многие другие вещества - кислоты, соли, аминокислоты, масла и пр. могут отвергаться при высокой концентрации, но иногда слабые растворы некоторых кислот и солей оказывают привлекающее воздействие. Показано также, что пчелы очень чувствительны к поваренной соли и отличают незначительную ее примесь к сахару - всего 0,36%, что человек обычно не различает. С другой стороны, пчелы могут пить сахарный раствор с примесью 0,02% хинина, тогда как для человека этот раствор очень горек.
Вкусовые рецепторы располагаются преимущественно на ротовых частях, но возможна и другая их локализация. Так, у пчелы, некоторых мух и ряда дневных бабочек они находятся на лапках ног и обнаруживают высокую чувствительность; при прикосновении подошвенной стороны лапок к раствору сахара голодная бабочка реагирует развертыванием хоботка. Наконец, у пчелы и складчатокрылых ос (Vespidae) эти рецепторы обнаружены и на концевых члениках усиков.
Высокая степень развития у насекомых химического чувства является существенной стороной их физиологии и служит научной основой при изыскании и применении некоторых приемов химической борьбы с вредными видами. В практике борьбы с вредителями применяют приманочный метод, сущность которого заключается в том, что те или иные привлекающие пищевые вещества обрабатываются ядами и распределяются в местах концентрации вредителя; такие отравленные приманки широко и очень успешно применяются в борьбе с саранчовыми. Для избавления от "укусов" таких докучливых кровососов, как комары, мошки, слепни и др., применяют отпугивающие вещества - репелленты, которыми смазываются открытые части тела. Мыслимо также применение в борьбе с вредителями и привлекающих веществ, или аттрактантов; в этом направлении практически еще ничего не сделано.
Гигротермическое чувство. Гигротермическое чувство имеет существенное значение в жизни ряда насекомых и в зависимости от условий влажности и температуры среды регулирует поведение особи; оно также контролирует водный баланс и температурный режим тела. Соответствующие рецепторы изучены недостаточно, но установлено, что ощущение влажности локализовано у некоторых насекомых на голове и ее придатках - усиках и щупальцах, ощущение тепла - на усиках, лапках и других органах.
Мучной хрущак (Tenebrio molitor L.) чувствителен к относительной влажности воздуха, избегает сырых мест и концентрируется в более сухих местах; следовательно, соответствующие чувствительные механизмы здесь действуют подобно гигрометру. Личинки жуков-щелкунов из рода Agriotes, называемые проволочниками, наоборот, стремятся к большей влажности воздуха и улавливают отличия в 7,5% влажности.
Восприятие тепла сильно развито у насекомых и отдельные виды имеют свою оптимальную температурную зону, к которой они стремятся. Так, у мухи осенней жигалки (Stomoxys calcitrans L.) температурный оптимум лежит в пределах 23-30°С, а у перелетной (Locusta migratoria L.) и пустынной саранчи (Schistocerca gregaria Forsk.) - в пределах примерно 30-42°С. Однако границы температурного оптимума зависят также от условий температуры и влажности среды, в которой развивалось насекомое, а также и от фазы его развития.
Зрение. Зрение вместе с химическим чувством вероятно играет ведущую роль в жизни насекомых. Органы зрения имеют сложное строение и представлены двоякого рода глазами: сложными и простыми. Сложные, или фасеточные, глаза в числе двух расположены по бокам головы, нередко очень сильно развиты и тогда могут занимать значительную часть головы. Каждый фасеточный глаз состоит из многих зрительных единиц - сенсилл, которые называются омматидиями (рис. 38, А); число их в сложном глазу может достигать многих сотен и даже тысяч.
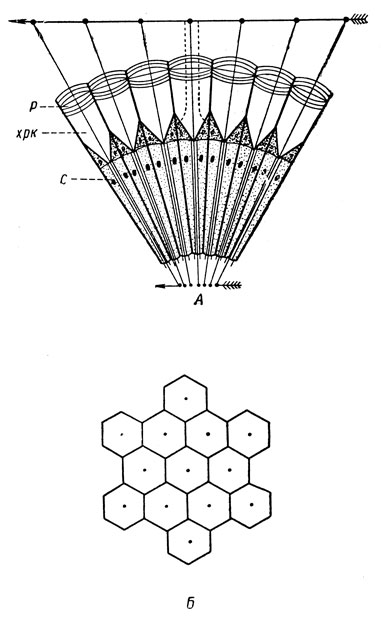
Рис. 38. Схематический разрез (А) и фасетки на поверхности (Б) сложного глаза (из Болдырева): р - роговица, хрк - хрустальный конус, с - клетки сетчатки
Омматидий (рис. 39) состоит из трех видов клеток, образующих оптическую, чувствительную и пигментную часть. Снаружи каждый омматидий образует на поверхности глаза округлую или шестигранную ячейку - фасетку (рис. 38, Б), отчего сложные глаза и получили свое название. Оптическая, или преломляющая, часть омматидия состоит из прозрачного хрусталика и лежащего под ним также прозрачного хрустального конуса. Хрусталик, или роговица, является в сущности прозрачной кутикулой, обычно имеет вид двояковыпуклой линзы. Хрустальный конус образован четырьмя удлиненными прозрачными клетками и совместно с хрусталиком составляет единую оптическую систему - цилиндрическую линзу; длина ее оптической оси значительно превосходит ее диаметр. Чувствительная часть располагается под оптической, образует воспринимающую световые лучи сетчатку, или ретину, и состоит из серии (6-13) ретинальных клеток. Эти клетки вытянуты вдоль омматидия, располагаются секториально и образуют обкладку его центрального стержня - зрительной палочки, или рабдома. У своего основания ретинальные клетки переходят в нервные волокна, идущие к зрительным долям головного мозга. Пигментная часть образована пигментными клетками, которые в совокупности составляют обкладку чувствительной части и хрустального конуса; благодаря этому каждый омматидий оптически изолирован от соседнего омматидия. Следовательно, пигментная часть выполняет функцию аппарата оптической изоляции.
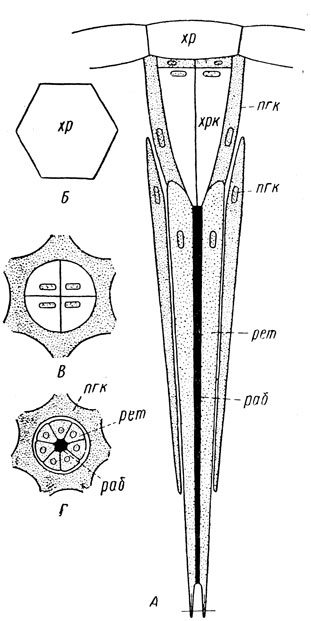
Рис 39. Схема строения омматидия аппозиционного глаза на продольном (А) и поперечном разрезах (Б-Г) (по Иммсу): хр - хрусталик, хрк - хрустальный конус, пгк - пигментные клетки, рет - ретинальные (зрительные) клетки, раб - рабдом
Дневные насекомые имеют так называемое аппозиционное зрение. Благодаря оптической изоляции с помощью пигментных клеток каждый омматидий превращен в изолированную тонкую трубку; поэтому в него могут проникнуть только лучи, идущие через хрусталик и притом только строго совпадающие с продольной осью омматидия. Эти лучи и достигают зрительной палочки, или рабдома; последняя как раз и является воспринимающим элементом сетчатки. Следовательно, поле зрения каждого омматидия очень мало и он видит только ничтожную часть рассматриваемого предмета. Но большое число омматидиев позволяет резко увеличить поле зрения путем взаимного приложения друг к другу, или аппозиции; в результате из отдельных мельчайших частей изображения образуется как в мозаике единое общее изображение. Таким образом, насекомые обладают мозаичным зрением.
Ночные и сумеречные насекомые обладают суперпозиционным зрением, что связано с морфологическими и физиологическими отличиями их омматидиев. В супперпозиционном глазе чувствительная часть более удалена от оптической, а пигментные клетки изолируют преимущественно оптическую часть. Благодаря этому к зрительной палочке проникают два вида лучей - прямые и косые; первые попадают в омматидий через его хрусталик, а вторые - из соседних омматидиев, что усиливает световой эффект. Следовательно, изображение предмета получается в данном случае не только путем объединения отдельных восприятий, но и путем их наложения, или суперпозиции.
При сильном дневном освещении суперпозиционный глаз приобретает некоторое физиологическое сходство с аппозиционным глазом. Происходит это потому, что пигмент в пигментных клетках на свету начинает перемещаться и распределяется так, что образует темную трубку вокруг омматидия; благодаря этому омматидии оптически почти изолируются друг от друга и получают лучи преимущественно от своей линзы. Эта способность глаза реагировать на степень освещения может рассматриваться как аккомодация. В некоторой степени она свойственна и аппозиционному глазу, что позволяет дневным насекомым быстро приспособлять глаз к зрению на ярком свету и в тени, например при перелете из открытого места в лес.
С помощью сложных глаз насекомые различают форму, движение, окраску и расстояние до предмета, а также поляризованный свет. Однако большое разнообразие насекомых, их образа жизни и повадок несомненно создает и разнообразие особенностей их зрения. Последние зависят от особенностей строения глаз и их омматидии; диаметр, длина, число последних и другие свойства определяют качество зрения. Считается, что многие виды близоруки и на расстоянии различают только движение; это подтверждается многими опытами. Так, личинки стрекоз бросаются на движущуюся добычу и не замечают неподвижной. Помещенная перед гнездом ос сетка с превосходящими длину их тела ячеями все же преграждает вход в гнездо; только через некоторое время осы научаются пролезать через ячеи этой сетки.
Большинство насекомых слепы к красному цвету, но видят ультрафиолетовое излучение и привлекаются им; диапазон видимых световых волн лежит в пределах 2500-8000 Å. У медоносной пчелы открыта способность различать поляризованный свет, испускаемый голубым небом, что позволяет ей ориентироваться в направлении при полете. Для ряда насекомых установлено также изменение движения в зависимости от направления солнечных лучей, т. е. ориентация по солнечному компасу. Сущность этого заключается в том, что угол падения лучей на те или иные части сетчатки сохраняет свое постоянство в течение какого-то времени; прерванное движение возобновляется под тем же углом, но ввиду перемещения солнца направление движения изменяется на то же число градусов (рис. 40, А).
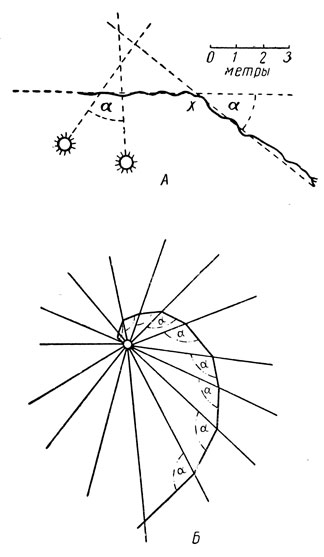
Рис. 40. Ориентация по солнцу и искусственному источнику света. А - путь муравья до и после точки х (в ней он был изолирован от солнца на 11/2 ч); Б - светокомпасное движение к искусственному источнику света (по Брюну и Буденброку): а - постоянный угол
Близким является светокомпасное движение, которое объясняет прилет ночных насекомых на свет. Световые лучи расходятся радиально и при косом движении по отношению к ним угол их падения будет меняться; для сохранения фиксированного угла насекомое вынуждено все время изменять свой путь в сторону источника света. Движение идет по логарифмической спирали и в конце концов приводит насекомое к самому источнику света (рис. 40, Б).
Простые глаза, или глазки, располагаются между сложными глазами на лбу и темени либо только на темени. Они малы, обычно в числе трех и расположены треугольником. Вследствие своего положения в верхней части головы они нередко называются также дорсальными глазками. Морфологически глазки не соответствуют омматидиям сложных глаз. Так, они иннервируются не из зрительных долей головного мозга, а из срединной части протоцеребрума. Помимо того, на одну оптическую часть у них приходится серия чувствительных частей. Они также лишены хрустального конуса и их оптическая часть представлена только кутикулярной линзой, т. е. одним хрусталиком (рис. 41).
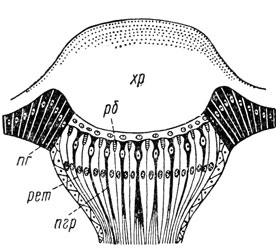
Рис. 41. Схема простого глаза (по Шванвичу): хр - хрусталик, рб - рабдом, пг - пигментированная гиподерма, рет - ретинальные клетки, пгр - пигментные клетки ретины
Глазки развиты далеко не у всех насекомых, в частности отсутствуют у многих двукрылых и бабочек. У бескрылых или короткокрылых насекомых они также отсутствуют или рудиментарны. Их функция недостаточно ясна. Установлено, что у ряда форм фокус глазка лежит за чувствительной частью, поэтому восприятия изображения в данном случае не может быть. Вместе с тем существует анатомическая связь глазковых нервов с нервами сложных глаз, что указывает на существование функциональной связи между этими органами. Присутствие глазков у хорошо летающих подвижных форм (стрекозы, тараканы, прямокрылые, перепончатокрылые и пр.) и отсутствие у многих бескрылых насекомых возможно указывает на регулирующую движения функцию. Считается также, что они стимулируют сложные глаза, усиливая их фотокинетическую реакцию.
От дорсальных глазков следует отличать боковые, или латеральные, глазки, свойственные личинкам насекомых с полным превращением. Эти глазки, называемые также стеммами, располагаются на боковых частях головы на месте, где у взрослых особей лежат сложные глаза. Число их различно и даже изменчиво в пределах одного и того же вида. Одни виды имеют всего лишь по одному глазку с каждой стороны, у других число их достигает шести и более пар. При переходе насекомого во взрослое состояние боковые глазки атрофируются и заменяются сложными глазами.
Стеммы разнообразны по деталям строения, но для них характерно присутствие хрусталика. У гусениц бабочек есть также хрустальный конус и развит всего один рабдом, что делает такой глазок сходным с омматидием сложного глаза. Но у личинок пилильщиков, некоторых жуков и других насекомых в глазке присутствует несколько или даже множество рабдомов, а хрустальный конус может отсутствовать; это делает такие стеммы сходными не с омматидиями, а с дорсальными глазками.
Боковые глазки иннервируются от зрительных долей головного мозга и их зрительная функция бесспорна.
Некоторые насекомые сохраняют способность реагировать на свет при удалении глаз и глазков или покрытии их черным лаком; тараканы при этом избегают света, как и в нормальном состоянии, а гусеницы сохраняют положительную реакцию и движутся к источнику света. Безглазые пещерные насекомые также могут реагировать на свет. Очевидно, поверхность тела насекомых способна ощущать свет и поэтому можно говорить о кожной светочувствительности.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'