16.2. Сообщества пустынь
Важными членами экосистем засушливых местообитаний являются термиты (табл. 16.1).
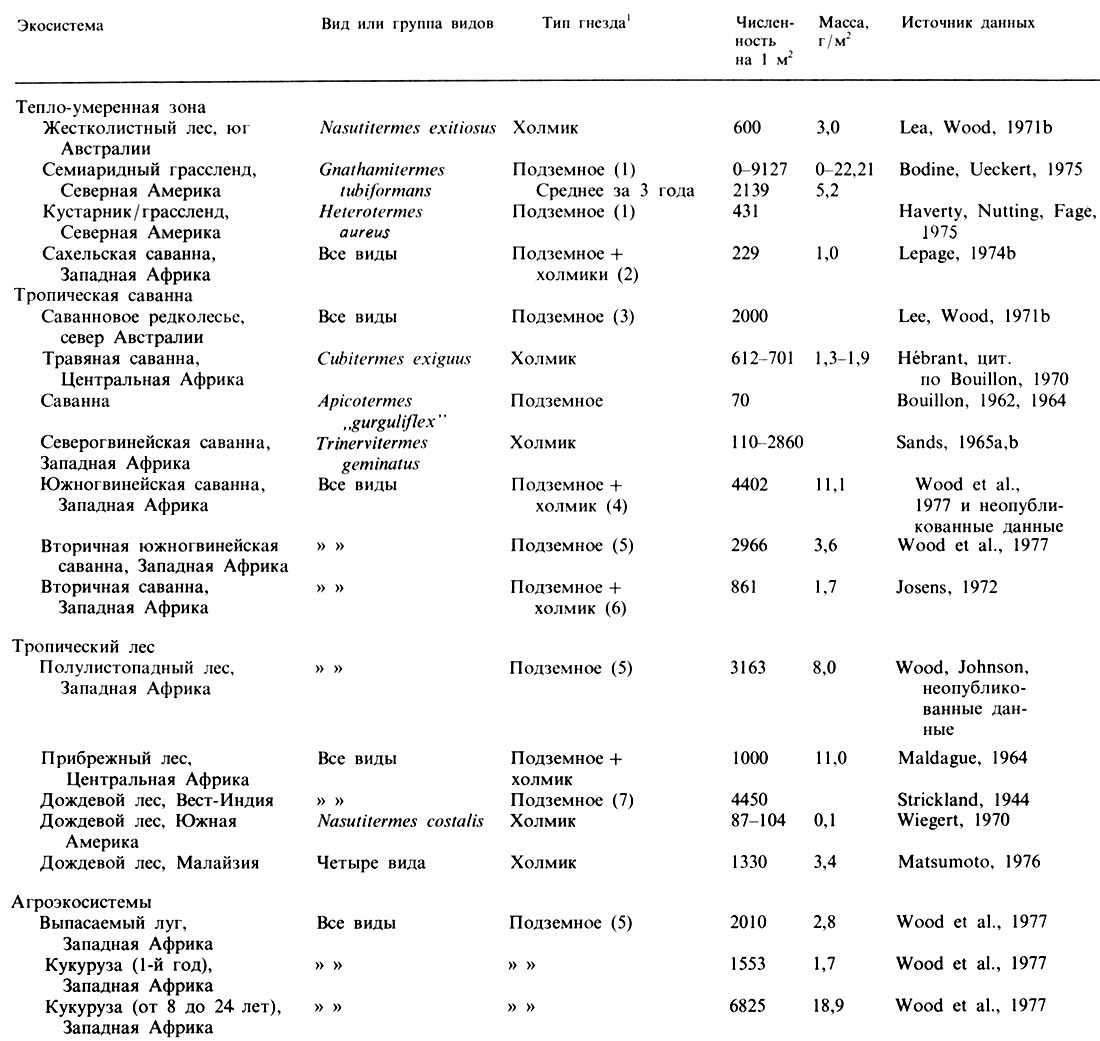
Таблица 16.1. Обилие и биомасса (живой вес) термитов в различных системах (Wood, Sands, 1978)
1(Пробы почвы брались с глубины 30 см (1), 75 см (2), 8 см (3), 200 см (4), 100 см (5), 50-60 см (6) и 7,5 см (7). В работах Josens (1972) и Lepage (1974b) пробы в случае подземных видов брались из ямок, выкопанных вручную; во всех остальных работах использовались керновые пробы.)
16.2.1. Термиты
Недавно Johnson и Wood (1979) опубликовали обзор по распространению термитов в аридных зонах Африки и Аравийского полуострова. Род Psammotermes (четыре вида), относящийся к семейству Phinotermitidae, встречается только в засушливых областях. В Сахаре эти термиты живут в оазисах и по краям пустыни; их гнезда расположены под землей на глубине до 5 м, что позволяет им избегать вредных последствий колебаний температуры, перевевания песка и иссушения. Они могут добывать воду для увлажнения стенок своих гнезд из грунтовых вод на глубине 40 м. Виды Psammotermes фуражируют под прикрытием лепных тоннелей, используя самую разнообразную пищу: навоз, выкопанную почву, подстилку, а в более влажные периоды - древесину.
В штате Аризона в горах Санта-Рита на высоте 950 м пять подземных видов потребляют разную пищу при разных способах питания (Haverti, Nutting, 1975; Haverty et al., 1975). Один из них, Heterotermes aureus (Rhinotermitidae), поедает мертвую древесину на поверхности почвы (56% Opuntia, 17% Acacia, 13% Prosopis), тогда как другие термиты предпочитают опунции акацию и поедают также Cercidium или злаки. Lee и Wood (1971) перечисляют 9 подземных видов, обитающих в зарослях кустарниковых эвкалиптов на юге Австралии. В африканской саванне, поросшей туссоковым злаком Loudetia, Bodot (1966) нашел 20 видов термитов, по большей части подземных, хотя некоторые из них строят холмики. Эти виды тоже различаются по своей пище; многие срезают сухие злаки и запасают их (Trinervitermes), другие поедают корни злаков (Amitermes evuncifer). Есть также гумусоядные виды, такие как Cubitermes severus и два вида Trinervitermes, которые кормятся одними и теми же злаками, но используют их по-разному (Bodot, 1966). Bodot описывает также, каким образом тип почвы, влажность и растительный покров влияют на распространение термитов в саванне Берега Слоновой Кости. В этом районе наблюдается постепенное исчезновение Macrotermes natalensis, возможно в результате вмешательства человека или какого-то изменения климата (или обоих этих факторов). Amitermes evuncifer поселяется в старых разрушающихся холмиках, а также вторгается в холмики Cubitermes severus и некоторых видов Trinervitermes, перестраивая их на свой лад (Bodot, 1966). Половые особи, по-видимому, падают на землю и обламывают крылья вблизи этих холмиков, а затем постепенно проникают в них; это относится в особенности к Т. geminatus (Josens, 1972).
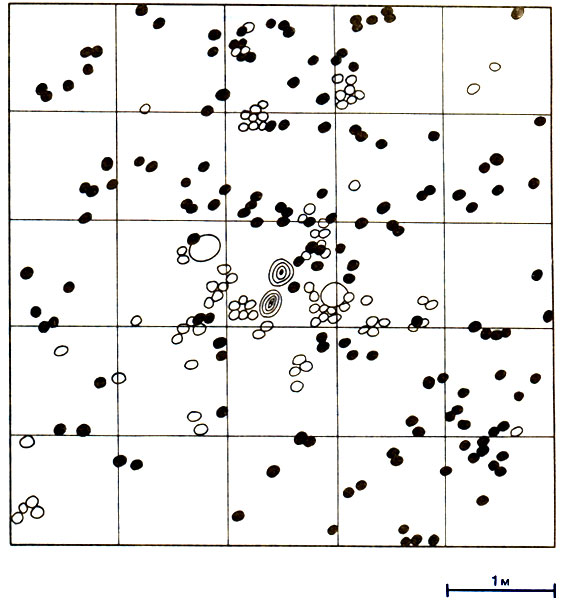
Рис. 16.5. Горизонтальное распределение грибных садов у двух термитов - Ancistrotermes cavithorax (белые кружки) и Microtermes toumediensis (темные кружки). В центре изображены основания двух деревьев (концентрические кольца). На этом участке находятся две репродуктивные камеры A. cavithorax, возможно, принадлежащие двум разным семьям (большие белые кружки). (Josens, 1972.)
Как установил Josens, в Ламто (Берег Слоновой Кости) Macrotermitinae, вероятно, конкурируют за пищу, но не за место для гнезда, поскольку большинство из них строят диффузные подземные гнезда; тем, кто строит холмики, могут помешать найти подходящее место другие термиты; злаки, которыми они питаются, имеются в изобилии, и 90% обитающих в этой области наземных термитов потребляют злаки. Josens обнаружил здесь 26 видов, из которых 11 встречаются очень редко. Остальные 15 видов можно разделить на следующие экологические группы: наземные - жнецы рода Trinervitermes (четыре вида) и один вид, питающийся древесиной (Amitermes evuncifer); подземные - Ancistrotermes cavithorax, Microtermes toumediensis, Odontotermes (вид неизвестен) и Pseudacanthotermes militaris, выращивающие грибы, один вид, питающийся древесиной без участия грибов (Microcerotermes parvulus), и пять гумусоядных видов (Josens, 1972). Идентифицировать отдельные семьи Macrotermitinae не удалось - имеются лишь косвенные сведения о существовании семей, и возможно, что они слились; однако Josens произвел подсчет среднего числа грибных камер на гектар и получил следующие данные: Ancistrotermes cavithorax - 21 500, Microtermes toumediensis - 24 400, Odontotermes - 3300 и Pseudacanthotermes militaris - 8000. Были получены также подробные сведения о составе этих популяций, их биомассе и превращении энергии (рис. 16.5 и 16.6). После ежегодных пожаров растительных остатков бывает мало, хотя кустарники и деревья сбрасывают листву; в таком случае между термитами должна быть конкуренция, несмотря на их диффузные гнезда, распределение по разным ярусам и различия в питании. Из 9920 приманок (связок травяных стеблей), расположенных в шахматном порядке, только 40 использовались одновременно двумя видами (Ancistrotermes cavithorax и Microtermes toumediensis). Поскольку в одиночку A. cavithorax использовал 2485, а М. toumediensis - 2059 приманок, то, если эти виды безразличны друг другу, число общих приманок должно было бы составить 511. В тех случаях, когда оба вида используют одну и ту же приманку, они устраивают земляную перегородку, чтобы отделиться друг от Друга.
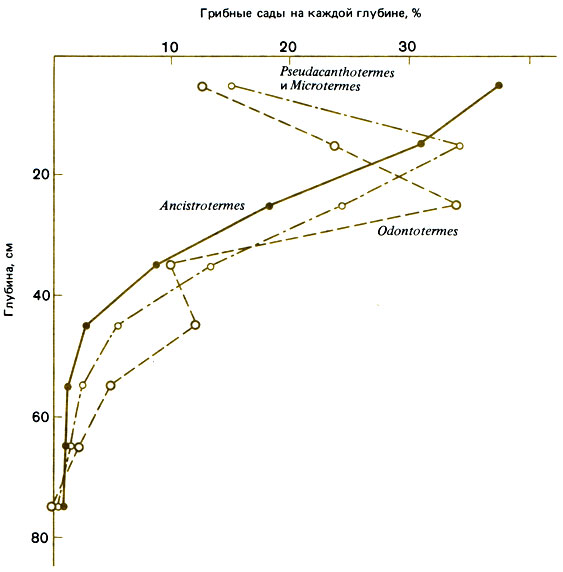
Рис. 16.6. Вертикальное распределение грибных камер (в процентах от их общего числа) четырех видов термитов: Ancistrotermes cavithorax; Odontotermes spp.; Pseudocanthotermes militaris и Microtermes toumediensis вместе. (Josens, 1972.)
Хотя примеры сообществ термитов и их описания многочисленны (см. Krishna, Weesner, 1970), прямых данных о взаимодействиях термитов мало. Согласно Noirot (1959), по крайней мере разные роды избегают встреч, a Bouillon (1970) приводит интересный пример взаимодействия, когда наличие гнездового холмика Macrotermes natalensis явно препятствует устройству гнезда Cubitermes sankurensis в радиусе 5-10 м. Такого рода конкуренция не обязательно приводит к схваткам; она связана с тем, что пища используется более эффективно, если ее источник находится вблизи гнезда. Hodotermes mossambicus и Trinervitermes trinervoides питаются злаками в одной и той же местности и обычно избегают друг друга, а если их насильственно смешать, то между ними возникают драки (Nel, 1968). В южной Австралии разные виды Coptotermes обитают в разных бревнах (Calaby, Gay, 1956, цит. по Lee, Wood, 1971). Плотность Nasutitermes exitiosus достигает 8,6-9,2 холмиков на гектар, а при наличии в данной местности Coptotermes lacteus - всего 2,4 на гектар; они рассеяны по занимаемой ими территории. У трех родов термитов, питающихся злаками, тоже наблюдается тенденция к дисперсии. Nasutitermes triodei в присутствии Hemitermes laurensis чаще покидает свои холмики; оба эти вида питаются злаками, и хотя первый из них поедает, кроме того, помет животных и растительные остатки, это его, очевидно, не спасает (Lee, Wood, 1971).
Сходные сообщества частично наземных термитов изучал Josens (1972) в Ламто. Он нашел на 1 гектаре в среднем 40,5 гнезда Trinervitermes geminatus (60% гнезд и 80% всей биомассы термитов), 14,7 гнезда Т. togoensis, 3,2 гнезда Amitermes evuncifer, 2,6 гнезда Т. oeconomus и в меньшем числе гнезда двух других видов Trinervitermes. В травяной саванне (главным образом Loudetia) есть бугорки, образовавшиеся из старых распадающихся холмиков Macrotermes; на этих хорошо дренируемых островках обосновывается Т. togoensis, получая таким образом возможность селиться в сырых, покрытых травой местах. Т. geminatus обитает на более открытых, но поросших кустарником участках, а численность Т. oeconomus достигает максимума в очень густой кустарниковой саванне. Гнезда Macrotermitinae, ведущих подземный образ жизни, настолько диффузны, что, по мнению Josens, они конкурируют только за пищу; что же касается наземных видов, среди которых 9/10 составляют термиты-жнецы, то, как показывают его расчеты, из 4-5,5 т/га (по сухому весу) общего количества собранного ими корма съедается только несколько десятков килограммов.
Sand (1965), изучавший Trinervitermes в саваннах Нигерии, уделял главное внимание сравнению видов, принадлежащих к одному роду, а не исследованию сообществ; тем не менее интересно отметить, что, как он обнаружил, в гнездах Trinervitermes обычно живут и другие термиты или же муравьи, а иногда те и другие. Обычно никаких контактов между разными видами как будто бы не бывает, но если вскрыть холмик, то "немедленно возникает сумбурный конфликт". В гнездах Trinervitermes trinervius живет один вид Microtermes, в гнездах Macrotermes natalensis - другой вид, но хозяева не обращают на них внимания. Ввиду резкого различия в размерах между сожителями эти взаимоотношения можно, по-видимому, считать типичным случаем парабиоза. La Fage и Nutting (1978), изучая взаимоотношения термитов в пустыне вблизи Таксона (Аризона), помещали бумажные приманки либо на сеточки, либо под поверхностный слой почвы. Термиты Heterotermes aureus прокладывали к 53% этих приманок узкие неровные галереи, a Gnathotermes perplexus - более широкие и аккуратные галереи к 96% приманок; к тем приманкам, которые использовали оба вида, каждый из них подходил по своим галереям, избегая галерей, проложенных другим видом.
Термиты нередко поселяются в гнездах других видов в качестве сожителей. Вид-сожитель часто бывает гораздо мельче вида-хозяина и менее активен; такие два вида могут быть либо родственными, либо очень далекими (Bouillon, 1970). Иногда этот парабиоз бывает облигатным, оба вида могут пользоваться одними и теми же галереями, и их репродуктивные пары находятся в тесной близости, а в других случаях вид-хозяин изолирует от сожителя свою царскую пару. Josens (1972) обнаружил вокруг камеры термита Ancistrotermes cavithorax и в ее стенках галереи значительно более мелкого гумусоядного термита, а также аналогичные галереи в стенках репродуктивной камеры Pseudacanthotermes militaris. Помимо такиях явных примеров использования одним видом гнезда другого вида термиты, строящие большие холмики, разрыхляют почву, помогая другим видам проникать в нее. В Южной Америке Comitermes cumulans строит в грассленде холмики высотой до 4 м. Если эти термиты покидают свои холмики, то в них поселяются общественные осы, муравьи и даже рептилии, а также другие термиты. Sands в своих экспериментах показал, что некоторые виды термитов предпочитают землю, взятую из холмика другого вида, обычной почве. Термиты размельчают не только почву; они могут размельчать древесину, чтобы снабжать гумусоядных термитов перегноем, создавая тем самым сукцессию от видов, питающихся древесиной, к гумусоядным видам.
На термитов часто охотятся муравьи; Camponotus, Crematogaster и Pheidole поедают Trinervitermes. Megaponera foetens не только охотится на многих Macrotermitinae, но и поселяется в их старых холмиках. В районе Моквы (9° с. ш.) из 83 изученных гнезд этого муравья 37 находились в заброшенных холмиках Macrotermes bellicosus, 3 - в холмиках Trinervitermes, 36 - в земле под кустами и 7 - на открытых местах (Longhurst, Howse 1979а, b). Доля общих гнезд с Trinervitermes geminatus варьирует в зависимости от местообитания и возрастает там, где много старых холмиков. При парабиозе муравьи обычно избегают термитов и безусловно не нападают на них, пока термитник не поврежден. Многие Dorylinae истребляют термитов, солдаты которых либо оказывают им сопротивление, защищая шаг за шагом свое гнездо от проникновения муравьев, либо уходят колоннами, двигаясь по поверхности земли, и возвращаются после прекращения атаки. Аnоmmа kohli может затратить несколько дней, чтобы проникнуть в большой холмик Cubitermes. Среди термитов Macrotermes bellicosus в Мокве, где их многочисленные холмики беспорядочно разбросаны по саванне, очень высока смертность от нападения хищников, и, как считает Collins (1981а), их плотность регулируется скорее хищниками, чем внутривидовой конкуренцией.
На одном гектаре в Ламто (Берег Слоновой Кости) можно обнаружить 1,6 млн. собирающих или срезающих траву термитов вместе с 4,5 млн. особей гумусоядных видов и 5 млн. термитов, выращивающих грибы (Josens, 1972). Фуражиры или жнецы потребляют 30-50 кг сухой травы в год, гумусоядные - примерно 30 кг целлюлозы в год и перекапывают не менее 15 тонн почвы за год, тогда как виды, выращивающие грибы, ежегодно вносят в почву около 1,3 тонны растительных остатков (сухой вес).
16.2.2. Муравьи
Муравьям, для того чтобы они могли выжить в пустыне, следует потреблять разнообразную пищу и включать в свой рацион такие растительные продукты, как, например, семена и листья. Из 97 видов муравьев, обитающих в Сахаре, 55 используют подземные источники воды (Delye, 1968); их кутикула менее проницаема для воды, чем у сходных европейских видов, и они активно избегают чрезмерной жары и сухости. Они устраивают гнезда везде, где есть растения, например вокруг оазисов, собирая и запасая семена во время короткого влажного периода и потребляя также насекомых, если они имеются. Ofer et al. (1978) описали почвы и климат, а также фауну муравьев (в общей сложности 49 видов) по трансекте, проведенной через пустыню в Израиле от побережья к горам. По статистическим данным о температуре, местообитания здесь можно разделить на теплую прибрежную пустыню, холмы и область ручьев, стекающих по северным склонам; оказалось, что в каждой зоне обитает свой, характерный для нее вид, а в целом виды, принадлежащие к одному роду, занимают разные местообитания. В самых прохладных районах был обнаружен даже Lasius alienus! Обследование, проведенное в горах Чирикауа в Аризоне, где прохладный влажный лес сменяется на юге сухими кустарниками с примесью суккулентов (Eastlake et al., 1980), показало, что Myrmicinae обычны на северо-западе и редки на юго-востоке, тогда как Dolichoderinae редки на севере, обычны на юго-западе и обильны на юго-востоке. Из семи зерноядных видов пять относятся к роду Pheidole (все они - мелкие муравьи), но три зерноядных вида, обычные на северном склоне, значительно различаются по величине и, возможно, поедают семена разных растений.
Корреляция между величиной муравьев и семян была установлена на трансекте, проходящей через пустыни Мохаве и Сонора (Калифорния и Нью-Мексико), выбранной потому, что она соответствует градиенту количества осадков (Davidson, 1977). В Калифорнии, где за год выпадает меньше 100 мм осадков и средняя температура июля 29-34°С, было найдено три вида зерноядных муравьев, а в Нью-Мексико (225-275 мм и 26-27°С соответственно) - 8 видов. Ни один вид не встречался по всей трансекте, но некоторые имелись на 6 из 10 станций. В Калифорнии обычны Veromessor pergandei и два вида Pogonomyrmex, тогда как в Нью-Мексико преобладают Novomessor cockerelli, Solenopsis xyloni, Pogonomyrmex negosus и Pheidole desertorum. Davidson нашла, что во влажных продуктивных местообитаниях не только больше видов, но и семьи их расположены плотнее, чем в более сухих местах. Дополнительные виды - это главным образом виды с мелкими рабочими особями (< 3 мм в длину), фуражирующими на расстоянии не более 3-4 м от своих гнезд, но есть также виды с крупными (> 9 мм) рабочими, например N. cockerelli, Pogonomyrmex rugosus и P. barbatus, фуражирующие в радиусе до 40 м (Hölldobler, 1976а). Длина Veromessor pergandei варьирует в пределах от 3,5 до 8,4 мм (при общем диапазоне длины особей всех видов от 1,8 до 9,8 мм), однако сильная конкуренция со стороны других видов ограничивает размеры его особей.
Все это позволяет предполагать, что пища как лимитирующий фактор играет более важную роль, чем место для гнезда, хотя Davidson не могла собрать веских данных, которые указывали бы на распределение этих зерноядных видов по разным зонам или разным ярусам или на какое-либо иное разграничение, обусловленное их биологией. Адаптация к семенам определенной величины, конечно, способствует подбору сосуществующих видов, однако Davidson (1977) приводит данные о том, что виды, различающиеся по размерам тела, сосуществуют только в том случае, если они различаются также по способу фуражировки. Виды, у которых рабочие фуражируют в одиночку, собирают семена на отдельно стоящих редких растениях, а виды, для которых характерна групповая фуражировка, используют растения, образующие плотные пятна. Таким образом, способ фуражировки муравьев соответствует характеру распределения их пищи. Виды, фуражирующие группами, усердно собирают семена при подходящей погоде и складывают их про запас, тогда как одиночные фуражиры работают все время, при любой погоде. Согласно Davidson, крупные виды могут фуражировать как группами, так и в одиночку (например, Pogonomyrmex rugosus), но все мелкие виды фуражируют группами (например, Pheidole).
Были обследованы пробные площадки в пустыне Большого Бассейна (от 44 до 36° с. ш.) и в пустыне Мохаве (35° с. ш.), расположенные на разной высоте, вплоть до 1500 м (Bernstein, 1975, 1979а, b; Bernstein, Gobbel, 1979). Novomessor pergandei был обнаружен только на четырех нижних площадках (500, 610, 700 и 830 м), a Pogonomyrmex rugosus - только на трех верхних (876, 1150 и 1500 м), тогда как P. californicus встречался на всех площадках от 610 до 1500 м, хотя и пятнами. На самых малых высотах - 75 и 500 м - обитает шесть видов, в том числе Veromessor pergandei, P. californicus и Conomyrma insana. На высоте 1500 м V. pergandei и С. insana отсутствуют, но P. californicus все еще встречается вместе с P. rugosus, Pheidole xerophila и Iridomyrmex pruinosum. Pogonomyrmex rugosus и P. californicus фуражируют в одиночку, а не группами; если только между разными популяциями P. rugosus нет какого-то генотипического различия, то придется предположить (основываясь на наблюдениях Hölldobler, 1976а), что они могут фуражировать в одиночку в тех случаях, когда их на это толкает характер распределения пищи. Фуражировка у V. pergandei может варьировать от групповой, если семян мало и они распределены пятнами, до индивидуальной в период образования семян, когда они распределены более или менее равномерно.
Диапазон температур, при которых муравьи отправляются на фуражировку, расширяется с высотой над уровнем моря и с широтой, а тем самым увеличивается и перекрывание между разными видами во времени. В жаркой и сухой малопродуктивной пустыне сбор корма у каждого вида ограничен коротким периодом, когда корма много и собирать его выгодно. Этот период, конечно, у разных видов приходится на разное время. Никаких признаков столкновений между фуражирами из разных семей не было. В исследованных диапазонах высоты над уровнем моря и широты обычна чрезмерная дисперсия гнезд муравьев какого-либо одного вида, обусловленная, вероятно, столкновениями между фуражирами, т. е. защитой кормовых участков. Переходя от сообществ больших высот и высоких широт к более низким местам и низким широтам, можно заметить, что у всех видов расположение семей группами, характерное для пятнистой многолетней растительности, которая создает местообитание, разделенное на зоны и ярусы, сменяется рассредоточенным и более однородным. Места для гнезд распределяются между видами случайным образом (Byron et al., 1980). Вообще, чем больше имеется видов, тем меньше перекрывание кормовых территорий независимо от широты или высоты. Bernstein (1979b) изучал также состав пищи муравьев. Муравьи потребляют либо семена, либо животную пищу, либо нектар. С уменьшением высоты местности в пустыне Большого Бассейна пища становится менее разнообразной и перекрывание пищевых спектров разных видов возрастает. В пустыне Сонора конкуренция между Novomessor cockerelli и Veromessor pergandei в периоды недостатка пищи усиливается, и в эти периоды первый из них потребляет больше животных, живых и мертвых, и меньше семян.
В пустыне Чиуауа были изучены (Whitford, 1978) два местообитания: бахада - широкий аллювиальный веер у входа в старое озеро, поросший креозотовым кустом (Larrea tridentata), и плайя - область с песчаной почвой вокруг высохшего озера, поросшая мескитовым деревом (Prosopis) и юккой (Yucca). В бахаде преобладают зерноядные муравьи - пять видов Pogonomyrmex и четыре вида Pheidole; они активны главным образом днем, и дожди стимулируют у них фуражировку. В плайе преобладают всеядные виды, такие как Novomessor cockerelli, Formica perpilosa, Iridomyrmex pruinosum и Myrmecocystus, если наибольшее разнообразие насекомых наблюдается ночью. Trachymyrmex smithi (Attini), собирающий детрит для своего сада, и хищный Neivamyrmex nigrescens (Dorylinae) распространены в обоих этих местообитаниях, хотя очевидно, что различия в пище, способах питания и изменения условий среды в течение суток имеют важное значение. Whitford установил также, что сосуществование видов облегчается различиями в общих размерах и в сезонной активности.
Интересную форму межвидовой конкуренции описали Möglich и Alpert (1979). В горах Чирикауа в Аризоне муравей Conomyrma bicolor (Dolichoderinae) собирает мелкие камешки и другие предметы и сбрасывает их через входные отверстия в гнезда других муравьев, в том числе трех видов Myrmecocystus, Novomessor cockerelli и Pogonomyrmex desertorum, с которыми он конкурирует за тлей на юкке и за термитов, Эти действия С. bicolor препятствуют фуражировке видов Myrmecocystus, но не P. desertorum. Когда муравьи С. bicolor находят гнездо, единственное отверстие которого охраняется привратниками, они собирают камешки и бросают их внутрь по одному или два в минуту; менее пяти камешков достаточно, чтобы прекратить фуражировку. Камешки не оказывают никакого механического воздействия, поскольку не производят эффекта, если их бросает экспериментатор; видимо, на них нет и феромона, однако они служат угрозой, предупреждающей о враждебных намерениях. Муравьи Conomyrma bicolor активны круглые сутки, за исключением полуденного времени, когда слишком жарко, так что они взаимодействуют с Myrmecocystus mextcanus ночью, а с двумя другими видами Myrmecocystus - днем. Если идет дождь (что случается редко), С. bicolor совершенно прекращает фуражировку и все три вида Myrmecocystus выходят из гнезд. Агрессивное поведение С. bicolor выражается гораздо сильнее возле многочисленных входов в его гнездо. Möglich и Alpert показали также, что это особое проявление общей реакции на запахи, так как С. bicolor кладет камешки на кусочки ваты, смоченной химическими веществами. Это весьма распространенная особенность поведения у муравьев вообще. Myrmica прикрывает растворы многих веществ (но не воду), причем выраженность этой реакции экспоненциально ослабевает с увеличением расстояния от входного отверстия собственного гнезда (Frith, 1979). Использование мелких камешков и частичек почвы для блокирования отверстий, ведущих в гнезда других муравьев, заставляет также вспомнить о баррикадах, которые строит Lasius flavus во время подземных схваток.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'