16.1. Луговые и лесные сообщества умеренной зоны
16.1.1. Грассленды Европы
В умеренных областях проводится много превосходных фаунистических исследований, но лишь в некоторых из них анализируются взаимоотношения между видами. В морском климате Стратклайда (56° с. ш.) на высотах 100-200 м над уровнем моря в сообщество помимо двух видов Myrmica входят еще два муравья: небольшой Leptothorax acervorum (Myrmicinae) и крупный Formica lemani (Formicinae). Поскольку ни тот ни другой не могут устраивать гнезда в траве, они живут только в местах, поросших кустарником, где есть участки голой почвы или же разлагающиеся пни и ветки. F. lemani строит гнезда под корой на южной стороне пней, Myrmica scabrinodis - в более прохладных местах, на сторонах, обращенных к востоку и западу, a M. ruginodis - в затененных участках. Такая температурная зональность устанавливается самими самками, когда они закладывают гнездо, а затем усиливается рабочими особями: F. lemani выселяют или убивают и съедают рабочих особей и расплод Myrmica. Leptothorax acervorum избегают уничтожения, устраивая тоннели в твердой неразрушенной древесине, куда за ними не могут последовать более крупные муравьи. По мере разрушения пней F. lemani переходят в растущую поблизости низкую траву, и хотя они могут нарушить рост травы, они не способны строить холмики, так что в конце концов затенение вынуждает их переселяться. Если они не могут найти подходящего места, они постепенно вымирают. Myrmica выживают в восстанавливающемся лесу, если на земле есть участки, освещаемые солнцем. Перед тем как в прошлом веке в Шотландии были сведены леса, можно было ожидать, что по мере появления молодых деревьев здесь будут поселяться F. lugubris и F. aquilonia, однако эти виды не очень подвижны, и им может понадобиться несколько десятилетий, чтобы пройти несколько километров.
Помимо такого зонального распределения гнезд разные виды муравьев фуражируют на разных уровнях. Formica залезает на кустарники и молодые деревца; Leptothorax acervorum фуражирует в нескольких метрах от гнезда и забирается в кусты, но главный источник его пищи неизвестен. Любые лесные муравьи, конечно, заползают еще выше, на деревья. F. lemani фуражирует главным образом в середине дня в солнечную погоду. Эта особенность поведения F. lemani уменьшает вероятность его контактов и конкуренции с двумя видами Myrmica, которые фуражируют утром и в позднее послеполуденное время, а также в пасмурную (но не сырую и ветреную) погоду. Ни у одного из этих видов нет охраняемых территорий; они энергично защищают свое гнездо и несколько скоплений тлей, прикрытых земляными навесами, но между гнездами существует лишь мало заметная система троп. Этот небольшой набор видов муравьев представляет собой сообщество, поскольку они занимают общее местообитание и делят его на сферы влияния, которые лишь незначительно перекрываются в пространстве и во времени. Их стремление избегать друг друга при изобилии ресурсов может перейти во враждебность, если ресурсов будет нехватать. В таких случаях система доминирования одних видов над другими может превратиться в пищевую цепь. Взаимовыгодным может оказаться кооперирование для защиты от позвоночных. Несколько южнее (51°с. ш., Дорсет) и всего лишь на 20 м выше уровня моря среди кустов вереска, все еще составляющего основную растительность, там и сям попадается Ulex minor. Это растение, возможно фиксирующее азот с помощью корневых клубеньков, дает семена, от которых L. alienus отгрызает присеменники, а на его стелющихся побегах обитают тли (Aphis ulicis), за которыми этот муравей ухаживает. L. niger обитает на дне укрытых прохладных сырых лощин и в относительно тенистых местах, заросших кустарником, его поселения и территории занимают площадь во много квадратных метров. Взаимоотношения этой пары видов мы уже обсуждали.
В центральной части этого сообщества доминирует третий вид - Tetramorium caespitum (Myrmicinae). Он происходит с юга и занимает наиболее прогреваемые зоны; для него характерны территориальная организация и подземные тоннели, в которых всегда имеются муравьи, готовые в случае надобности защищать гнездо от вторжения или отправиться к найденному фуражирами источнику пищи (рис. 16.1). Кроме того, он использует семенные ресурсы местообитания, не ограничиваясь одними лишь семенами вересковых, но собирая также семена различных злаков; муравьи собирают их на земле, а семена вереска в конце лета - прямо из коробочек, хранят всю зиму и поедают весной. Т. caespitum охотится также на мелких членистоногих, выращивает тлей и фактически обладает множеством "экологических шупалец", глубоко проникающих своими разветвлениями в биом верещатника. Таким образом, ядро этого местообитания состоит из одного эволюционно продвинутого представителя Myrmicinae и двух видов Lasius. В нем встречаются также, хотя и на периферии сообщества, представители родов, обитающих севернее: Formica lemani здесь замещен довольно сходным с ним южным вариантом - F. fusca, который не является доминирующим видом, а по существу живет в ассоциации с Т. caespitum; он устраивает гнезда на территории последнего, но избегает контактов благодаря тому, что в его гнезде имеется лишь одно маленькое входное отверстие, и фуражирует он на более удаленных кустах и на большей высоте; эти два вида, возможно, полезны друг другу. На сухих верещатниках Myrmica scabrinodis замещен видом М. sabuleti, который больше нуждается в тепле и может жить на грасслендах как с нейтральной, так и с кислой почвой. М. ruginodis на большей части травянисто-кустарниковых участков с нейтральной почвой замещен видом М. rubra, но сохраняется в кустарниках, в лесах и на верховых болотах. В грасслендах с нейтральной почвой М. rubra встречается с Lasius flams, но занимает несколько иную микростацию, более затененную, с широкими плоскими камнями для гнезд; здесь он питается, по крайней мере частично, за счет тлей, живущих на травах. В местах, достаточно хорошо прогреваемых солнцем, L. flavus может вытеснить М. rubra и покрыть землей камень, под которым жили эти муравьи (Elmer, 1974). Лесные муравьи опять-таки встречаются здесь спорадически, но представлены южным F. rufa, а не северными F. lugubris или F. aquilonia.
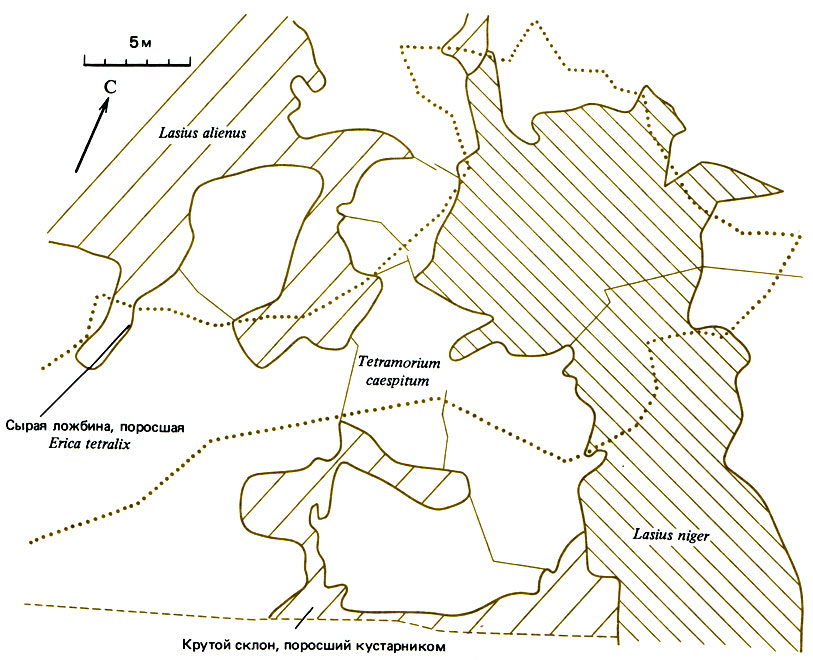
Рис. 16.1. Сообщество верещатника в Дорсете. Толстыми линиями разделены территории, занимаемые разными видами, а тонкими - разными семьями. Lasius niger обитает в тени крутого склона и в сырой ложбине (пунктирная линия), L. alienus - в более возвышенной местности к западу, а Tetramorium caespitum - между ними
Развитие лесов приостанавливается из-за вызываемых человеком пожаров. Лишь небольшая доля муравьев гибнет непосредственно от тепловой волны, распространяющейся от поверхности почвы вглубь, но последующий недостаток корма может оказать опустошающее действие. Муравьи, обитающие в почве, могут еще питаться членистоногими и собирать мелких насекомых, падающих на землю во время расселительных полетов, но необходимые для этого более обширные поиски пищи вскоре приводят к встрече с соседними семьями и видами. Lasius alienus временно расширяет свои территории за счет Tetramorium, но по мере восстановления растительности на прежних местах возвращаются на них и эти муравьи, так что численность обоих видов колеблется вместе с циклом пожар - восстановление и никогда не достигает стабильности. Как показывают результаты анализа главных компонент, в каждой микростации имеется один представитель Formicinae и один - Myrmicinae (Brian el al., 1976). Очевидно, оба эти подсемейства проникли в местообитания всех типов.
Мы уже рассматривали муравьев острова Рамси - это виды, принадлежащие к родам Lasius и Mymrica. Важное различие между этими родами состоит в том, что расположение их гнезд, наиболее плотное у побережья, различается по степени агрегации: у Myrmica гнезда менее агрегированы, чем у Lasius, а из трех видов Lasius наименее агрегированы гнезда L. flavus. Несмотря на агрегированность, распределение плотностей этих видов близко к случайному, причем большая часть учетных площадок содержит по одному виду или не содержит их вовсе, а площадки, содержащие по 4 или 5 видов, редки. Это может быть обусловлено двумя противоположными тенденциями: стремлением собирать корм в местах, предпочитаемых всеми видами, и взаимным ограничением плотностей в результате конкуренции (Doncaster, 1981). Сообщество острова Рамси служит примером состояния, промежуточного между ситуациями на западе Шотландии и на юге Англии: хотя вид Formica lemani замещен видом F.fusca, Myrmica ruginodis и М. scabrinodis здесь присутствуют, так же как и М. sabuleti. Tetramorium caespitum встречается только в двух из наиболее укрытых прибрежных местообитаний, но ни в одном из них он не распространен настолько, чтобы доминировать над всей фауной муравьев, как это имеет место на юге Англии. То же самое можно сказать о Lasius niger, который вместе с L. flavus преобладает в одной области, поросшей злаками (Festuca rubra) и защищенной от сильных ветров; он устраивает также гнезда у каменных стен. Что касается отношения к главным компонентам среды, то виды Myrmica чувствительны к дренированности почвы (выбирают влажные поля в низинах) и, по-видимому, безразлично относятся к освещенности, которая влияет на распределение видов Lasius: L. niger и L. flavus занимают затененные места, a L. alienus - открытые. Таким образом, в этой местности выявляются только три локальные системы доминирования: во главе одной из них стоит Т. caespitum, другой - L. niger, а третьей - L. alienus. На возвышенных местах, заросших орляком, популяция L. flavus достигает высокой плотности и вытесняет другие виды в результате более интенсивного потребления ресурсов, так как это не агрессивный муравей. В тех местах, где его биомасса невелика, с ним сосуществуют другие виды, живущие в почве ближе к поверхности. Виды с широкой адаптивной способностью (L. flavus, Myrmica sabuleti и М. scabrinodis) встречаются в разнообразных местообитаниях, тогда как специализированные доминанты (Tetramorium caespitum, L. niger и L. alienus) ограничены в своем распространении и агрегированы. Во всех местообитаниях имеются виды Myrmica и Lasius. Myrmica чаще встречаются на тех площадках, где обитают виды Lasius, чем в тех, где их нет; в одной трети площадок с L. alienus обитает и какой-нибудь вид Myrmica. Виды Myrmica могут сосуществовать с Lasius и Formica, занимая более влажные промежуточные участки, как в Дорсете. Представителей рода Formica может ограничивать отсутствие достаточно толстого слоя почвы, необходимого им для устройства гнезда и обеспечивающего рост кустов, в которых можно было бы фуражировать. Таким образом, на острове Рамси сливаются сообщества Стратклайда и Дорсета.
На высокогорных лугах в Апеннинах, на известняковых скалах на высоте 1700 м (42° с.ш.) Baroni Urbani (1969) также изучал Tetramorium caespitum и Lasius alienus. Здесь, выше границы леса, среди злаков преобладают Brachypodium pinnatum и Festuca ovina, а наряду с главными видами муравьев - L. alienus (55%) и Т. caespitum (36%) - встречаются также Formica fusca, F. rufibarbis и Myrmica sulcinodis. L. alienus более обилен на сравнительно оголенной почве, где растет Festuca, но никакой другой зональности в распределении этих видов не заметно, а гнезда L. alienus и Т. caespitum распределены случайным образом. Baroni Urbani не мог выявить никаких признаков территориального образа жизни, столь ясно выраженного в Дорсете, но он заметил, что в августе L. alienus был склонен фуражировать по ночам и в одиночку. Обычно оба эти вида начинают фуражировать с восходом солнца, усиливают свою активность к полудню, а затем отдыхают до вечера, после чего пики их активности вновь совпадают. У L. alienus наблюдается более строгая периодичность, чем у Т. caespitum. С чем связано отсутствие территориальности у Т. caespitum в Апеннинах - с климатом или с условиями среды в грасслендах, - остается не ясным; возможно, что устройство горизонтальных тоннелей в дернине слишком затруднено. Муравьи заселяют и болота. В верещатниках на юге Англии в заболоченных местах обитают главным образом Lasius niger, Myrmica ruginodis и M. scabrinodis, но кое-где, на покрытых травой кочках и других островках растительности, устраивает свои гнезда Formica transcaucasica (F. picea)*. Lasius niger, несомненно, приспособлен к жизни в местах, затопляемых водой. В Венгрии (47° с. ш.) он живет в ложбинках, прорытых ветром в песчаной почве, a L. alienus строит гнезда на разделяющих эти ложбинки гребнях (Galle, 1980). На песчаной прибрежной равнине между Северным морем и Ваддензе L. niger проникает в места, которые зимой оказываются под водой, особенно на участки, где растет лишь несколько злаков (главным образом овсяница Festuca rubra), а другой растительности мало. Myrmica rubra, но не его северный двойник М. ruginodis, встречается в нижней зоне дюн в дернине, образованной овсяницей, и в зарослях Linum catharticum на песчаной, но не слишком засоленной почве. М. scabrinodis в дюнах редок, но встречается на небольших холмиках между ними с дерниной, образованной овсяницей, где почва содержит больше ила и солей и где растут галофиты. Lasius flavus здесь отсутствует, вероятно из-за тонкозернистой скудной почвы и недостатка растительности, однако он очень часто встречается в близлежащих местах (Boomsma, de Vries, 1980). L. flavus выживает в соленой воде при 5°С в течение нескольких дней (Nielsen, 1977), а на юге штата Мичиган обитает по краям водоемов, где он строит гнезда близко к поверхности почвы, никогда не проникая в глубину и не насыпая холмики выбранной земли (Talbot, 1965). Другие данные о том, что L. flavus может жить по краям соленых маршей и что его гнезда зарастают сверху овсяницей Festuca rubra, которую щиплют кролики, приводит Woodell (1974), изучавший этих муравьев в восточной Англии. Он обнаружил здесь на южной части земляных холмиков Frankenia laevis - кустарник, обычно встречающийся в Средиземноморье. L. flavus выносит из подпочвенных слоев частицы средних размеров, и его гнездо имеет очень пористую структуру.
*(В нашей литературе принято второе из этих названий. - Прим. ред.)
16.1.2. Степь и кустарники
На Британских островах в сообществе грасслендов виды Formica представлены слабо, но в степях Западной Сибири есть сообщества, в которых доминируют F. pratensis и F. uralensis (Стебаев, Резникова, 1972, 1974). Между обширными территориями, занятыми этими двумя видами, умещаются небольшие территории других видов, например F. subpilosa и F. transcaucasica. Каждое утро территория этих видов расширяется, а поздним вечером сокращается, причем F. subpilosa, менее трудолюбивый муравей, чем F. transcaucasica, обычно фуражирует только после того, как F. pratensis удалится в гнездо на ночь (в 22 часа); если же вид F. subpilosa занимает данную территорию один, то пик его активности приходится на 18 часов. Поселяющийся между ними вид F. transcaucasica служит временным хозяином для самок F. uralensis (Collingwood, 1979); он, кроме того, помогает F. uralensis отыскивать спрятанные мясные приманки, но не приманки, выставленные на виду. Очевидно, у F. uralensis выработалась привычка проверять, чем заняты муравьи F. transcaucasica, или реагировать на оставленный ими след и мобилизационные феромоны. В сущности, F. uralensis и F. transcaucasica не враждебны друг другу; можно предполагать, что последний вид пользуется некоторым покровительством со стороны F. uralensis в обмен на помощь, оказываемую их самкам-основательницам, и нахождение для них пищи; их отношения носят характер симбиоза (мутуализма), а не парабиоза (разделения ресурсов).
В западной Сибири, где чистая степь перемежается с березовыми колками, можно встретить другое сообщество, в которое входят Formica pratensis и F. canicularia (вид, близкий к F. fusca), а также широко распространенные Tetramorium caespitum, Lasius alienus и Myrmica scabrinodis (Peznikova, Kulikov, 1978). Значительную часть их пищи составляют живые и мертвые луговые растительноядные беспозвоночные, в том числе различные кобылки, сверчки, мухи, цикадки и иные насекомые, но прежде всего - другие муравьи, сменяющие друг друга в сезонном цикле, среди которых преобладают молодые особи. В частности, у М. scabrinodis пища на 70% состоит из высохших остатков, среди которых 40% приходится на других муравьев, главным образом Formica. Однако 80% пищи самих Formica составляют свежеубитые беспозвоночные на ювенильных стадиях развития, причем разнообразие потребляемых видов у F. pratensis больше, чем у муравья F. cunicularis, который поедает мелких обитателей травяного яруса. На кормовых территориях F. pratensis может находиться 4-7 семей F. cunicularis, а также 10-12 семей М. scabrinodis. F. pratensis отправляется на охоту группами, нападая на живых прямокрылых, тогда как F. cunicularis умеет находить этих насекомых, но не может справиться с ними; как показала в своих экспериментах Резникова (1971, 1975), F. cunicularis повышает эффективность фуражировки F. pratensis, отыскивая для них этих крупных насекомых. Возможно, что F. pratensis отнимает у F. cunicularis значительную часть их крупной добычи; если отгородить муравьев F. pratensis сеткой, то они сами добывают для себя больше крупных насекомых. Как указывает Yasuno (1964a,b, 1965а-с), в Японии виды муравьев с крупными рабочими особями и большими семьями распределены независимо от видов с мелкими рабочими и небольшими семьями. К последней группе относятся Tetramorium caespitum, Pheidole fervida и Paratrechina flaviceps. Поскольку все крупные виды принадлежат к подсемейству Formicinae, а два из трех мелких - к подсемейству Myrmicinae, мы видим здесь явное сходство с картиной сосуществования, наблюдаемой в травянисто-верещатниковой микростации в Дорсете.
В сообществе муравьев на одном пастбище в штате Луизиана (31° с. ш.) Baroni Urbani и Kannowski (1974) обнаружили преобладание холмиков и территорий Solenopsis invicta, однако там обычны также Monomorium minimum и Paratrechina arenivaga, тогда как Cyphomyrmex rimosus встречается реже и распределен независимо от этих видов, а Solenopsis xyloni обитает только в тени деревьев. М. minimum, многие семьи которого живут в холмиках S. invicta, нападает на последних, выбрасывая на них струю репеллента, что заставляет рабочих особей S. invicta заняться чисткой тела, вместо того чтобы мобилизовать на помощь своих товарок; эта тактика оказывается успешной, даже если М. minimum и не обитает в холмиках S. invicta. С S. xyloni тот же вид М. minimum справляется далеко не так успешно; ему удается создать популяцию, но затем рабочие S. xyloni убивают этих муравьев, раздавливая их своими челюстями. Таким образом, в затененных местах М. minimum оказывается в особенно невыгодном положении. На открытых местах при дневном свете малая величина муравьев М. minimum компенсируется наличием у них распыляемого репеллента; к тому же они не часто встречаются с S. invicta, который более активен в ночное время.
Несколько севернее (39° с. ш.) во влажном климате штата Мэриленд в листопадных лесах (Quercus, Fagus, Саrуа) с сильно развитым подлеском обитает 24 вида муравьев; четыре главных вида - это Aphaenogaster rudis (Myrmicinae) (27%), Prenolepis imparls (Formicinae) (23%), Paratrechina melanderi (Formicinae) (20%) и Camponotus ferrugineus (Formicinae) (6%). Lynch et al. (1980) установили, что Prenolepis imparls живет большими стабильными семьями в глубоких постоянных гнездах, а в его наземной активности наблюдаются два пика - в мае и в ноябре. Он избегает температуры 26°С и выше, но достаточно активен при таких низких температурах, как 10°С (рис. 16.2). В период с июля по октябрь A. rudis и Paratrechina melanderi активны на поверхности земли днем, тогда как С. ferrugineus активен ночью. Такое распределение фуражировки во времени предотвращает столкновения между тремя из этих четырех видов, так что периоды активности перекрываются только у A. rudis и Paratrechina melanderi. Последний вид с его мелкими рабочими особями и небольшими семьями быстро находит пищу, но рабочим приходится привлекать помощников, чтобы унести ее в гнездо. Prenolepis imparls сильно превосходит другие виды по способности мобилизовать помощников. Кормовые ресурсы лучше всех защищает, конечно, С. ferrugineus, который может убить P. imparts, но P. imparls очень агрессивен и занимает второе место в иерархии доминирования. Только схватки между этими двумя видами могут иметь серьезные последствия; у других видов дело ограничивается преследованием, укусами и приставаниями. P. imparis может занять половину сетки с приманками за 15 мин, а затем медленно распространиться на остальные приманки; если муравьям предлагают прямокрылых, то 66% забирает Prenolepis imparts, 26% - Aphaenogaster rudis и только 4% - Paratrechina melanderi. Prenolepis imparls не может помешать тому, чтобы A. rudis забрал себе часть пищи, если она дается в виде мелких приманок, распределенных диффузно, но может монополизировать крупные приманки, расположенные отдельными пятнами. Доминирующий вид (Camponotus ferrugineus) ведет скрытный образ жизни и фуражирует по ночам; следующий за ним вид (Prenolepis imparis) живет глубоко под землей и избегает летней жары, предоставляя в это время поверхность земли эвритопному неспециализированному A. rudis (Myrmicinae) с его спутником Paratrechina melanderi (Formicinae). Подобное распределение видов по времени активности наблюдается на всем земном шаре. В сообществах южной Австралии виды Iridomyrmex занимают доминирующее положение, активны днем и не перекрываются в пространстве (рис. 16.3); между их территориями остаются промежутки, в которых могут обитать виды, принадлежащие к другим родам, таким как Camponotus и Melophorus (Greenslade, 1979); представители первого из этих родов - крупные, трудноуловимые, неспециализированные и могут кормиться ночью, а представители другого фуражируют в жаркие часы дня. Iridomyrmex purpureus на юге Австралии обитает только на открытых местах; ему нужно солнце, и он плохо передвигается по траве. Этот муравей обходит на вытянутых ногах границы своей территории и благодаря этой демонстрации избегает смертельных схваток (Greenslade, 1975); в этом он сходен с Myrmecocystus mimicus (рис. 16.4; Hdlldobler, 1976а).
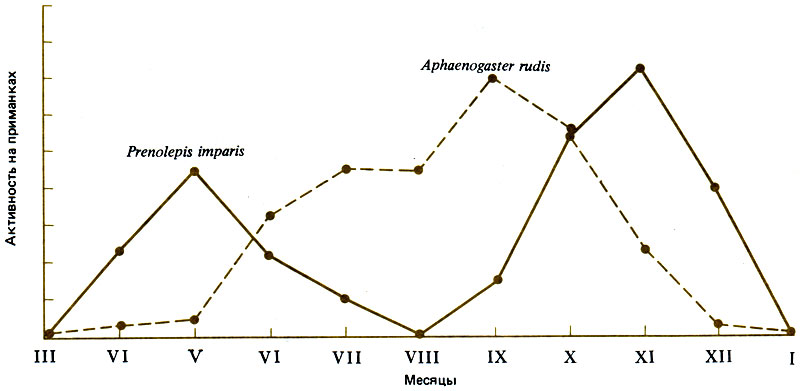
Рис. 16.2. Сезонная комплементарность фуражировочной активности двух видов муравьев на лесной подстилке в листопадном лесу в штате Мэриленд. (Lynch et al., 1980.)
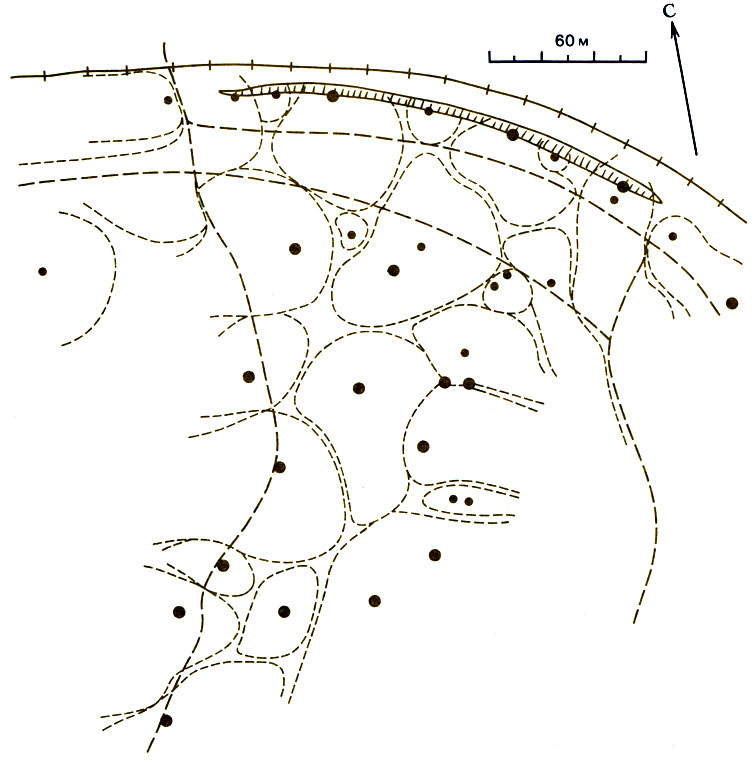
Рис. 16.3. Распределение территорий (границы очерчены прерывистыми линиями) и гнезд (черные кружки) Iridomyrmex purpureus (Greenslade, 1975а.)
Рабовладение известно только в трех трибах муравьев: Tetramorini, Leptothoracini (Myrmicinae) и Formicini (Formicinae) (Büschinger et al., 1980). Эта крайне сильная взаимозависимость возникла только в травянисто-кустарниковых сообществах северного умеренного пояса с высокой плотностью популяций видов - "рабов": 2% видов являются социальными паразитами и 17,5% - "рабовладельцами". Все это моногинные виды, использующие в качестве хозяев также моногинные виды, хотя некоторые из них факультативно полигинные, а один - Polyergus lucidus - может даже быть в течение какого-то времени полигинным, однако его добавочные самки совершают набеги и вместе с несколькими рабочими особями основывают новое гнездо. Polyergus и Strongylognathus сильными заостренными челюстями прокалывают головы непокорных рабов. Можно представить себе много гипотетических стадий, в результате которых возникает рабовладение: сначала самки после брачного полета могут попасть в гнезда одного из близко родственных видов; часто такой социальный паразитизм бывает лишь временным, а затем пришлая самка строит собственное гнездо. Но если ее собственные рабочие особи специализированы для набегов, то ей придется "послать" их в набег на соседние семьи "хозяина" за куколками.
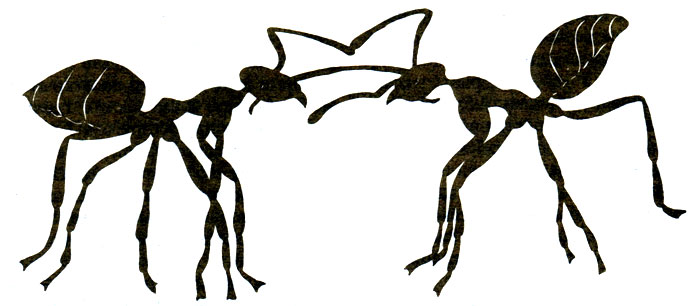
Рис. 16.4. Демонстративное поведение рабочих особей Myrmecocystus mimicus на границе территории. (Hölldobler, 1976а.)
Муравей Formica sanguinea, по-видимому, занимает доминирующее положение в травянисто-кустарниковых местообитаниях в лесах Евразии. Он ведет себя агрессивно по отношению к F. rufa и F. pratensis, сначала унося из их гнезд куколок, а затем уничтожая рабочих особей; из некоторых куколок, унесенных в гнездо F. sanguinea, выходят взрослые особи, которые становятся членами новой семьи и даже отказываются признавать самок своего вида. Рабочие F. fusca и F. rufibarbis настроены к F. sanguinea менее враждебно, и семьи этих видов могут избежать уничтожения. Вид F. sanguinea агрессивен также по отношению к Lasius niger, L.flavus и Camponotus herculeanus и может уничтожать их гнезда, но он может также существовать самостоятельно, питаясь падью и насекомыми, например лесными муравьями (Marikovsky, 1963). Более специализированный социальный паразит (рабовладелец) Polyergus rufescens совершает набеги на гнезда F. fusca и F. cinerea. Проведение набега зависит от различий в реакции со стороны хозяев (Dobrzánska, 1978): муравьи F. fusca, у которых входы в гнездо закамуфлированы, выносят свой расплод и спасаются бегством, взбираясь на растения; поликалический вид F. cinerea, которому свойствен обмен рабочими между гнездами, строит баррикады. Эти способы защиты эффективны при нападениях F. sanguinea, но атаки P. rufescens бывают так внезапны и сокрушительны, что на оборону просто не хватает времени. "Рабы", находящиеся в гнезде, тоже ведут себя во время набега по-разному: муравьи F. fusca расширяют свои входные отверстия и уменьшают заторы, создаваемые возвращающимися Polyergus rufescens; что касается F. cinerea, то они просто подбирают своих куколок, которых P. rufescens бросает снаружи, и даже могут отнимать их силой. Частично такое поведение развивается в результате научения, постепенно достигая совершенства. У P. rufescens, рабы которых принадлежат к виду F. rufibarbis, сезон активности начинается раньше. Судя по имеющимся данным, вид P. rufescens не делится на расы, различающиеся по своим "жертвам"; Dobrzánska даже сообщает о нескольких случаях, когда в одной семье P. rufescens рабы относились к нескольким разным видам.
Расселяющиеся полигинные формы Formica rufa по мере своего распространения вытесняют другие виды и могут даже уничтожить F. fusca, в гнездах которого молодые одиночные самки F. rufa основывают новые семьи как временные социальные паразиты. Некоторые виды используют лесных муравьев: Formicoxenus nitidulus (Mirmicinae) устраивает гнезда в их холмиках, Diplorhoptrum fugax уничтожает их расплод, a Leptothorax обитает в оболочке их холмика. Распространяющийся на острове Хоккайдо подвид Formica truncorum yessensis, вероятно, вытесняет многие более мелкие виды Lasius, Aphaenogaster и Myrmica, но проворный F. japonica живет на открытых местах, недоступных F. truncorum yessensis, тогда как и Diplorhoptrum, и Leptothorax сосуществуют с F. t. yessensis (Higashi, Yamauchi, 1979). Расселяющаяся популяция F. lugubris в горах Юры не уничтожает относящихся к Myrmicinae Tetramorium caespitum, Leptothorax acervorum, Monica rubida и три вида Myrmica (Cherix, 1980). F. opaciventris (из группы F. exsectoides) встречается только в западной части Северной Америки, и Scherba (1964) отмечает, что в штате Вайоминг на высоте 2065 м на полынном лугу, окруженном тополями, ивами и соснами, его гнезда находятся на расстоянии примерно 6 м одно от другого; этот муравей, по-видимому, не допускает на луга муравьев F. fusca, которые живут только на открытых местах в лесу. В тех местах, где эти виды встречаются друг с другом, F. opaciventris осаждает гнезда F. fusca, постепенно уничтожает его рабочих особей и уносит расплод, но он может делать это только на лугах; в лесу F. fusca эффективно сопротивляется, часто временно покидая свое гнездо.
Еще один пример относится к Японии, где Yasuno (1964а, b, 1965а-с) на высоте до 600 м (41° с. ш) изучал сообщество грассленда, окруженного буковым лесом. В одних районах доминирует Formica exsecta*, а в других - F. truncorum. Последний поселяется на хорошо прогреваемых лесных опушках, а если удалить его, то его место занимает F. fusca**, приходящий с открытых грасслендов, или Camponotus herculeanus - с травянисто-кустарниковых участков. F. truncorum - поликалический вид, создающий новые семьи, но он может также использовать F. fusca в качестве временного хозяина. Polyergus rufescens избегает Formica truncorum и даже не трогает гнезд F. fusca, если они находятся на территории F. truncorum, так что последний фактически защищает F. fusca там, где может. Непосредственно вокруг своего гнезда P. rufescens искореняет муравьев F. fusca, которые, однако, обладают очень высокой приспособляемостью и быстро возвращаются назад, если удалить P. rufescens. Таким образом, F. fusca составляет основу сообщества муравьев; он пользуется защитой со стороны доминантного вида (F. truncorum), который использует его в качестве временного хозяина, и сам используется подчиненным видом (P. rufescens) в качестве хозяина (раба). F. truncorum удерживает также Myrmica ruginodis в прохладных местах в глубине леса.
*(По современной номенклатуре F. japonica. - Прим. ред.)
**(По современной номенклатуре F. fukaii. - Прим. ред.)
Рабовладельцы подчиняют себе самку вида-хозяина (например, Strongylognathus karawajewi), препятствуя развитию половых особей из ее расплода; по-видимому, все яйца мужского пола (гаплоидные), которые она откладывает, уничтожаются, а развитие диплоидных яиц подавляется, если у личинок заметны признаки развития в самок. Такая семья может существовать лишь до тех пор, пока жива самка вида-хозяина. В случае ее уничтожения семье, чтобы продолжать свое существование после гибели рабочих вида-хозяина, приходится время от времени добывать замещающих рабочих (в виде куколок) из разных гнезд. Таким образом, рабовладение возникает из конкуренции или неэффективного хищничества, при котором собранные при набегах куколки не поедаются. Развивающиеся из них рабочие особи способны при этом научиться заниматься тем, с чем они сталкиваются в своем непосредственном окружении. Только виды, проявляющие способность к научению в раннем возрасте, могут стать рабами; по всей вероятности, виды с полностью врожденным поведением (как, например, Lasius niger) не пригодны для этого (Le Moli, 1980). Однако L. niger нередко служит хозяином для самок L. umbratus, которые оказываются в роли временных социальных паразитов.
Рабовладение ведет к быстрой утрате нормального независимого поведения видом, использующим рабов. Североамериканский Leptothorax duloticus совершенно не способен собирать и перерабатывать твердую пищу, хотя он все еще умеет строить гнезда и заботиться о расплоде. Если удалить рабов (L. curvispinosus), его забота о расплоде возрастает, но не в такой степени, чтобы он мог самостоятельно выжить (Wilson, 1975). L. duloticus, добывающий личинок и куколок во время набегов, обладает рядом мелких морфологических особенностей, облегчающих переноску крупных объектов (эти же особенности имеются у рабовладельческих видов Formica). Wilson указывает также на черты поведения, которые могут быть выгодными для рабов; например, их рабочие особи откладывают яйца и добавляют их к кучке яиц, отложенных самкой L. duloticus. Хотя некоторые из этих яиц бывают съедены, рабы, возможно, производят нескольких самцов, которые, по-видимому, копулируют с молодыми самками из гнезд без паразитов и распространяют гены рабства. Благодаря этому они могли бы пользоваться покровительством рабовладельцев в обмен на выкармливание их молоди. Такого рода мутуализм напоминает "парабиоз", при котором фуражиры предоставляют свою способность отыскивать пищу в обмен на обеспечиваемую им защиту.
Этот беглый обзор сообществ муравьев в областях с умеренным климатом показывает, что межвидовая иерархия доминирования построена по некоему общему плану. Доминантный вид удерживает за собой первоклассные кормовые зоны с помощью силы и жестокости отдельных особей, своей организованности и "экономической мощи", но при этом вокруг остается множество мест с субоптимальными условиями, где могут существовать виды, приспособленные к более низким температурам, обладающие особыми навыками в добывании корма или устройстве гнезд или же просто достаточно скрытные. В стабильных условиях развивается полигиния, ведущая к социальному паразитизму и рабовладельчеству; но если рабы способны размножаться партеногенетическим путем, то такие отношения не обязательно выгодны лишь одной стороне, так как рабы при этом пользуются защитой со стороны доминирующего вида.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'