16.3. Тропический дождевой лес
16.3.1. Осы, пчелы и термиты
Постоянное тепло, ежедневные дожди и 12-часовой световой день в течение всего года создают в тропиках необычайно продуктивные экосистемы. Общественные насекомые достигают в тропическом дождевом лесу максимального видового разнообразия, и хотя социальный паразитизм и рабовладельчество у них отсутствуют, здесь встречается много типов межвидового сотрудничества. Так, например, самки Mischocyttarus immarginatus в относительно сухом лесу в Коста-Рике конкурируют за места около строящегося гнезда Polybia occidentalis (Gorton, 1978). Эти самки в буквальном смысле слова устраивали драки, кусая и отталкивая одна другую, а после того как гнезда были построены, проводили время в набегах и столкновениях. Из 24 гнезд М. immarginatus 22 располагались около гнезда P. occidentalis; возможно, что его обитатели в какой-то мере защищали их от ящериц и птиц. Деревья, в которых обитают муравьи, нередко тоже служат "защитными зонтиками": число осиных и даже птичьих гнезд бывает здесь больше, чем можно было бы отнести на счет простой случайности (Evans, West-Eberhard, 1973). Рекордный пример описали Richards и Richards (1951): восемь гнезд Polybia rejecta, два - различных видов Mischocyttarus, одно - Protopolybia, одно - Metapolybia и еще гнездо иволги. Осы покладисты и удаляются в свои гнезда, тогда как муравьи защищают дерево от всех пришельцев. Хотя для тропиков характерно сотрудничество, там достаточно много и конкурентных отношений. Они могут принимать очень утонченные формы: "африканизированные" медоносные пчелы (в Гвиане) сокращают время, которое безжалые пчелы проводят на цветках, собирая пыльцу (Roubik, 1978); они оказывают влияние не на общее число фуражиров, а только на время, проводимое за сбором пыльцы. У Apis mellifera, интродуцированной несколько веков назад в Америку, хоботок примерно такой же длины, как у местных видов Bombus, тогда как в Евразии, откуда происходит род Apis и где он эволюционировал, у многих шмелей хоботки длиннее, и конкуренция с ними менее сильна (Pleasants, 1981; Prys-Jones, 1982). Может быть, в Евразии представителям Bombus пришлось удлинить свои хоботки, чтобы выжить? Не достаточно ли других различий, существующих между Apis и Bombus?
В западной части Малайзии обитает 57 видов термитов, относящихся к 23 родам из шести подсемейств (Matsumoto, Abe, 1979). На одном гектаре можно найти представителей 52 видов! Биомасса термитов достигает 3-4 кг/м2 (сырой вес), или 2,2-2,6 г/м2 (сухой вес). Большинство из них строит либо диффузные гнезда в земле, либо башни высотой до 2 м, иногда опирающиеся на основание дерева (рис. 16.7). На деревьях и кустах они либо выгрызают полости в отмерших ветвях, либо строят в живых ветвях или вокруг них картонные гнезда, сообщающиеся с поверхностью почвы крытыми галереями. Расстояния между гнездами варьируют от вида к виду, но в целом надземные гнезда распределены случайным образом. Тридцать семь видов питаются мертвой древесиной в лесной подстилке, 24 вида - остатками листьев, но есть также 17 гумусоядных видов, а два вида поедают лишайники, сдирая их со стволов деревьев, нередко на высоте до 40 м от земли. Два главных вида Macrotermes уже были рассмотрены. Longipeditermes longipes спускаются с деревьев вниз и под охраной солдат Отрезают кусочки от лежащих на земле листьев. Bulbitermes, гнезда которых находятся на высоте 1-9 м, тоже спускаются вниз по крытым галереям и подбирают с земли кусочки разлагающихся веточек и листьев. Некоторые живущие под землей гумусоядные формы поднимаются на поверхность земли, которая, очевидно, представляет собой зону очень активной жизни. Все это показывает, что стратификация гнезд мало связана с тем, в каком растительном ярусе питается данный вид, так как термиты готовы преодолевать расстояние между гнездом и источником пищи либо под эскортом солдат, либо по крытым галереям. В целом можно сказать, что в тропическом дождевом лесу обитают многочисленные виды термитов, образующие главный компонент почвенной микрофауны, что они устраивают гнезда в земле, на деревьях и в кустах, что все они питаются волокнистым растительным материалом, часто не из того яруса, где расположено гнездо, и что больше половины видов находятся в симбиозе с грибами. В девственном саванновом редколесье обитает 23 вида термитов: 33% относятся к роду Microcerotermes, 26% - к Ancistrotermes и 25% - к Microtermes. Представители последнего рода строят сухие диффузные гнезда, могут жить на возделываемых участках и проникать в корни кукурузы (Wood et al., 1977, 1980).
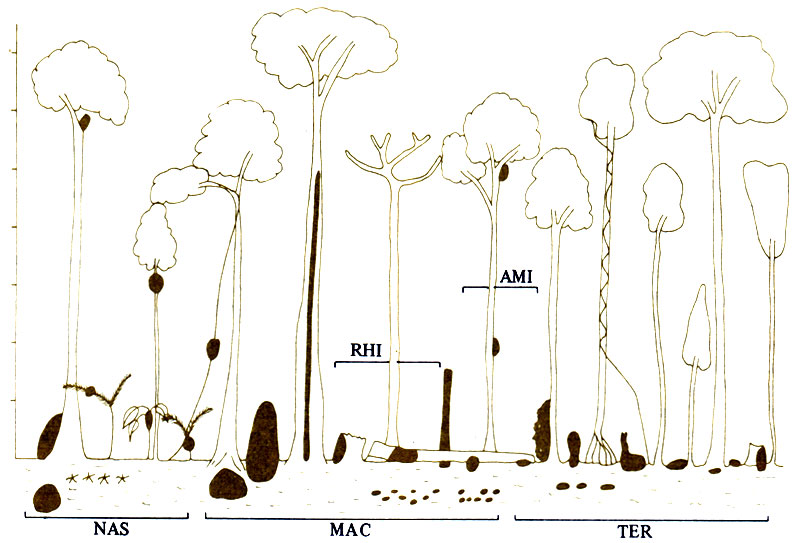
Рис. 16.7. Стратификация гнезд термитов в тропическом дождевом лесу Пасох-Форест в западной части Малайзии. RHI - Rhinotermitidae, три рода; AMI - Amitermitidae, два рода; TER-Termitinae, пять родов; MAC - Macrotermitinae, три рода; NAS - Nasutitermitinae, пять родов. (Abe, Matsumoto, 1979.)
Гнезда термитов привлекают многих других беспозвоночных. На одном из участков лесного резервата Доуала-Эдеа (3° с. ш.) на берегу озера 52% гнезд были построены термитом Cubitermes fungifaber и 37% из них были заняты вторичными хозяевами, но сожительство наблюдалось редко (Collins, 1980). Обычно появление нового хозяина не сказывается на внешнем виде гнезда и его форме, однако Noditermes покрывает гнездо свежим материалом. Collins (1981) перечисляет 31 вид, использующий гнезда пяти термитов, строящих холмики.
16.3.2. Муравьи в лесу
Число видов муравьев, населяющих дождевые леса в тропиках, огромно. Существует 29 родов, распространенных во всех тропических областях. Кроме того, 65 родов эндемичны для неотропической области, а для менее изолированной Африки - только 31 род. В Индо-Австралийской области, где имеется 42 эндемичных рода, некоторые роды несколько примитивнее африканских, и это позволяет думать, что африканские роды возникли позднее. Пантропические роды также представлены в Африке и в Южной Америке разными видами. Brown (1973) высказал предположение, что по крайней мере четыре рода распространились по земному шару в олигоцене - миоцене; это Tetramorium, Pheidole, Crematogaster (все они относятся к Mirmicinae) и Camponotus (Formicinae). Он привлекает внимание к тому факту, что Crematogaster, известный лишь начиная с миоцена, теперь распространен повсюду в тропиках и проник в "адаптивную зону" (нишу), которую прежде занимал Iridomyrmex, доминировавший в фауне муравьев олигоцена*. В настоящее время эти роды замещают друг друга: в Африке Crematogaster представлен 175 видами, а Iridomyrmex отсутствует; в индо-малайской области обитают 125 видов Crematogaster и 5 видов Iridomyrmex; в Австралии - 30 видов Crematogaster и 80 - Iridomyrmex. Brown (1973) указывает также, что в северном полушарии со времен миоцена роды Pheidole, Crematogaster, Tetramorium и Myrmica, принадлежащие к Myrmicinae, в значительной степени вытеснили роды, относящиеся к Dolichoderinae. Род Pheidole со времен олигоцена (балтийский янтарь), по-видимому, образовал сотни видов, расселившихся по областям земного шара с теплым климатом**. Tetramorium - тоже сложный род, очень обычный в южных районах палеарктической области, в Африке, в Индо-Малайской области и в северной части Австралийской области, но представленный в Америке всего одним видом.
*(Следует отметить, что муравьи эоцена и олигоцена известны только из Европы и Северной Америки, а о древней фауне нынешних тропических областей мы ничего не знаем. В миоценовом доминиканском янтаре найдены виды, близкие к современным.- Прим. ред.)
**(По современным представлениям, балтийский янтарь, в котором доминируют виды Iridomyrmex (более 50% всех муравьев), датируется поздним эоценом. - Прим. ред.)
По данным Lévieux (1975, 1977), в почве галерейного леса обитает 30 видов муравьев, тогда как рядом, в саванне, в почве встречается 120 видов, из которых лишь 20-общие для обоих местообитаний. Конечно, в лесу мало травы, и многие муравьи, которые здесь фуражируют, спускаются, подобно термитам, вниз из своих расположенных на деревьях гнезд. Lévieux указывает на контраст между видами, особи которых имеют крупные размеры и фуражируют на обширных территориях, как, например, Megaponera, и более мелкими, такими как Crematogaster, с локальными кормовыми участками. Хорошее описание тропических муравьев дает Bequaert (1922), рассматривающий муравьев бассейна Конго в Африке. Обеднение фауны хищных муравьев по мере удаления от экватора ярко продемонстрировал Jeanne (1979а), приманивавший их на личинок ос в низменных влажных местах в Америке на пяти широтах: 2° ю. ш., 10°, 19°, 31° и 43° с. ш. В первом из этих мест он поймал 74 вида, а в последнем - только 22. Леса обычно отличались большим разнообразием видов, чем поля, причем в лесах всех широт на земле встречалось больше видов, чем на деревьях. Во всех широтах наиболее обычным был род Pheidole, a Solenopsis geminata оказался наиболее широко распространенным видом в полях к югу от 43° с. ш., где он замещает Formica obscuriventris. Хищник Eciton, представленный здесь в изобилии, никогда не приходил к приманкам.
Муравьи-кочевники (Dorylinae) распространены по всему тропическому поясу и играют очень важную роль как хищники. Представители рода Neivamyrmex могут подниматься в горы и переносят короткие зимы, но большая их часть обитает в местах с жарким, хотя и не обязательно влажным климатом. Многие виды живут под землей, а другие - на земле и на деревьях. Aenictus - род мелких лишенных глаз мономорфных муравьев, представленных многочисленными видами в тропических районах Азии и Африки и живущих главным образом под землей, однако Schneirla (1971) специально изучал два наземных вида этого рода - A. gracilis и A. laeviceps. Эти муравьи отправляются в свои рейды в сумерки, однако освещенность не имеет для них значения, и они могут отдыхать на поверхности почвы. Каждый рейд продолжается час или два, причем муравьи могут покрывать расстояния до 25 м. На время стационарной фазы, когда расплод находится на стадии куколок и происходит откладка яиц, муравьи уходят под землю. Они поедают большое количество расплода других муравьев, термитов, ос и тараканов; очень быстро, проворно и эффективно действуя своими жалами, они обездвиживают жертву и утаскивают ее совместными усилиями. Эти виды справляются даже с такими крупными муравьями, как Polyrachls и Camponotus, и временно складывают их в кучи у разветвлений троп. Schneirla описывает также различия в распределении Eciton burchellt и Е. hamatum и в их стратегии при набегах. Е. vagans, который охотится на поверхности земли, возможно, поедает больше муравьев, чем другие виды. Как сообщает Schneirla, семьи Eciton и Neivamyrmex относятся друг к другу враждебно, и ни один из них не использует троп, проложенных другим видом. Подводя итоги, можно сказать, что у муравьев-кочевников рабочие особи имеют различные размеры, образуют семьи разной величины и совершают свои набеги на разных уровнях экосистемы, используя различные стратегии; все это расширяет диапазон их экологических возможностей, однако у нас пока еще очень мало сведений о том, каким образом они распределяются в пространстве, - известно лишь, что при разделении семей они перемещаются в противоположных направлениях.
Колонну муравьев-кочевников нередко сопровождают птицы, относящиеся к нескольким семействам, - дрозды, совы и муравьеловковые птицы из неотропических Formicariidae. Они поедают насекомых, которые идут сплошным потоком. На острове Барро-Колорадо (зона Панамского канала) примерно три семьи Eciton burchellt на 1 км2, движущиеся фронтом шириной 20 м со скоростью 15 м/ч, постоянно сопровождают птицы, добывающие таким образом не менее половины своей пищи (Willis, Oniki, 1978). Цитированные авторы перечисляют по меньшей мере 50 "профессионалов", среди которых 4 вида кукушек, 13 древолазов, 5 танагр и 28 муравьеловок. Они делят между собой зоны, причем доминантные виды охотятся ближе всего к центру колонны муравьев, а более мелкие подчиненные виды - на периферии. Некоторые из этих последних очень проворны, и их отнюдь не отпугивает агрессивность доминантных птиц. Промежутки между зонами, принадлежащими "профессиональным" спутникам бродячих муравьев, часто используют менее специализированные птицы. Следуя за колонной муравьев, эти птицы непрерывно кричат, привлекая внимание других птиц.
Weber (1972) описывает листорезов (Attini) как уживчивых муравьев. Он наблюдал в Гайане представителей Асrоmуrmех, срезавших травинки на той же кочке, что и термит Amitermes. Он указывает также, что гнезда этих насекомых частично вклиниваются друг в друга; "материала, необходимого для устройства гнезд, хватает на всех, и они фуражируют рядом без всякой враждебности", пишет Weber. При этом, однако, они обычно избегают контакта с другими видами. Weber наблюдал, как цепочка Atta cephalotes встретилась с колонной Eciton burchelli, как они перемешались, не проявляя враждебности, разделились и разошлись; это особенно удивительно, так как Dorylinae поедают многих Attini. Как и в холмиках термитов, в холмиках крупных Atta часто поселяются многие другие муравьи. Weber упоминает Trachymyrmex, Sericomyrmex и Myrmicocrypta, рабочие особи которых фуражируют друг над другом. Две семьи, содержавшиеся в лаборатории, использовали одни и те же цветки розы, но отрезали кусочки каждый вид по отдельности; Weber отмечает, однако, что Trachymyrmex "явно занимал доминирующее положение при отрезании субстрата", и если этого требовали обстоятельства, выступал как агрессор. Таким образом, у Attini определенно существует межвидовая враждебность, но в обычных условиях они избегают проявлять ее. Jutsum (1979) подтвердил, что представители разных семей или разных видов довольно часто кусают друг друга, однако по возможности предпочитают избегать этого. В Тринидаде Atta cephalotes обычно обитает в лесу, а Асrоmуrmех octospinosus - на открытых местах, причем создается впечатление, что самки выбирают эти местообитания после брачного лёта. Работа Jutsum не оставляет сомнений, что, как и у большинства других муравьев, после того как самка выбрала тип местообитания в целом, рабочие особи находят конкретное место для гнезда.
В тропиках рабовладельцы среди муравьев неизвестны, однако там существует интересная альтернатива, которая, возможно, встречается чаще, чем предполагают. Известны случаи, когда два (или более) вида муравьев живут в одном и том же гнезде, совместно пользуются галереями и тропами, ухаживают за одними и теми же сосущими растительноядными насекомыми, хорошо относятся друг к другу, но при этом содержат свой расплод по отдельности. В описанных случаях виды-сожители не связаны родством и могут принадлежать к разным подсемействам. Недавно Swain (1980) изучал в Бразилии муравья Crematogaster limata (Myrmicinae), который объединяется либо с Camponotus femoratus (Formicinae), либо с Monads debilis (Dolichoderinae). Cr. limata достигает всего 2,3 мм в длину, тогда как длина Camponotus femoratus 5,0 мм, а М. debilts 4,0 мм. Обитая вместе с М. debilis, Cr. limata находит сахарные приманки первым, но рабочие особи М. debilis, которые сами никогда не находят приманки, отнимают их, не прибегая к насилию. Хотя Cr. limata выставляет наружу свое жало и поднимает брюшко, по всей видимости выпуская репеллент, это не заставляет М. debilis отступить. Таким образом, М. debilis использует Сr. limata для нахождения пищи. Если же в качестве приманки используются личинки ос, то Cr. limata не уступает их, и муравьям М. debilis приходится искать приманки самим.
Взаимоотношения между Crematogaster limata и Camponotus femoratus яснее. Особи Cr. limata все еще находят приманку первыми, из чего бы она ни состояла, но если это сахарная приманка, то С. femoratus приходят одновременно с ними, нередко отстраняя Cr. limata; когда же приманкой служит личинка осы, то они перелезают через Cr. limata, но вместо того чтобы, подобно последним, разрезать ее на части, пытаются утащить ее целиком. Муравьи Camponotus femoratus обнаруживают и монополизируют приманку, не подпуская к ней Cr. limata, - для этого они подгибают брюшко; они могут просто поднять рабочую особь Cr. limata, ухватив ее своими челюстями, и унести в сторону, но никогда не раздавливают ее. Таким образом, С. femoratus полностью доминируют в данном сообществе, а Cr. limata не может воспользоваться добычей, даже если она перенесена в гнездо, так как камеры для расплода у каждого вида свои. Swain указывает, что хотя муравьи Cr. limata находят пищу для других видов и занимают подчиненное положение, они, возможно, получают взамен защиту и гнездо, поскольку именно С. femoratus строит гнездо, в котором поселяется Cr. limata, а не наоборот. Jeanne (1979а), описывая результаты своих наблюдений на трансекте, пересекавшей Америку с 2° ю. ш. до 43° с. ш., отмечает, что С. femoratus и Cr. limata пользуются общими тропами и общими гнездами и что добыча этих двух видов составляет 70% добычи всех хищников в подлеске бразильских лесов.
16.3.3. Муравьи на плантациях
Сообществу муравьев на плантациях кокосовой пальмы и шоколадного дерева уделяется большое внимание, и они уже были описаны автором этой книги (Brian, 1965); здесь мы рассмотрим некоторые из более новых исследований и проанализируем структуру сообществ. На Соломоновых островах на кокосовых пальмах совместно обитают четыре массовых вида и свыше 60 более редких (Brown, 1959; Greenslade, 1971). Два из массовых видов - Iridomyrmex cordatus (Dolichoderinae) и Oecophylla smaragdina (Formicinae) - местные, а два других - Pheidole megacephala (Myrmicinae) и Anoplolepis longipes (Formicinae) -завезенные. Более редкие виды лишь иногда обитают на одних и тех же деревьях с четырьмя главными, но есть существенные исключения. Любую попытку установить, каков порядок доминирования среди этих четырех видов, затрудняют необъяснимые флуктуации у A. longipes и Ph. megacephala. На одном обследованном участке, где были все виды, кроме Ph. megacephala, ежемесячные проверки изменений в составе обитателей показали, что О. smaragdina сменила место в 136 случаях, A. longipes - в 96, а I. cordatus - только в 6 случаях. Это позволяет думать, что I.cordatus, однажды заняв дерево, способен удерживать его за собой; после дальнейших наблюдений была установлена такая последовательность: I. cordatus > A. longipes = Ph. mega-cephala > О. smaragdina. Последний вид способен элиминировать все второстепенные виды, хотя он это делает не всегда. Когда по той или иной причине Ph. megacephala погибает, его место занимают субдоминанты.
Прямые способы борьбы занимают лишь небольшое место среди факторов, определяющих успех. Исследование, проведенное на кокосовой плантации на одном из океанических островов, позволяет предположить, что важную роль в этом играют шесть моментов: 1) Oecophylla smaragdina фуражирует в жаркий солнечный полдень, у Anoplolepis longipes фуражировка достигает пика в сумерки, a Iridomyrmex cordatus и Pheidole megacephala - ночные виды; 2) популяции видов-иммигрантов A. longipes и Ph. megacephala нестабильны; 3) по величине отдельных особей виды располагаются в следующем порядке (от мелких к крупным): I. cordatus, Ph. megacephala, A. longipes, О. smaragdina; мелкие муравьи могут прятаться в щелях и совместными усилиями справляются с крупным муравьем, но рабочие особи О. smaragdina, объединившись, могут поймать и убить жертву, слишком крупную для всех остальных; 4) О. smaragdina - единственный моногинный вид в данном сообществе; если самка в семье умирает, то муравьям приходится начинать все сначала, а для этого нужно отправиться на поиски новых мест; полигинный вид I. cordatus расселяется медленно и основательно, занимая все доступные территории при рациональной плотности; 5) полностью древесный образ жизни, как у О. smaragdina, не всегда дает преимущество, так как гнезда, вырытые в земле, служат убежищами, из которых затем можно расселяться; I. cordatus может строить гнезда в земле и вообще обладает более высокой приспособляемостью, чем О. smaragdina; 6) способность некоторое время прожить, питаясь одной только падью, дает преимущество в случае осады, и в этом отношении четыре вида располагаются в следующем порядке: I. cordatus, О. sniaragdina, Ph. megacephala, A. longipes; последний вид, будучи хищником, подвергается опасным лишениям, оказавшись отрезанным от гнезда на верхушке дерева. В целом создается картина флуктуирующей конкуренции между четырьмя массовыми и многочисленными более редкими видами, которые устраиваются там и тут, как только представляется удобный случай. Возможно, что до появления Ph. megacephala и A. longipes здесь существовало некое равновесие между двумя четко различающимися древесными видами - медленно расселяющимся полигинным видом I. cordatus с мелкими рабочими особями, активными ночью, и способным к быстрому расселению (летающие молодые самки!), но легко уязвимым моногинным видом О. smaragdina с крупными рабочими особями, активными днем.
В то время как распространение Pheidole megacephala и Anoplolepis longipes на Соломоновых островах сдерживается двумя сильными местными видами, на других тропических островах дело обстоит иначе (Haskins, Haskins, 1965). Гавайские острова в настоящее время заселяют "аргентинский" муравей Iridomyrmex humilis, который живет по обе стороны от экватора в полосе между 30 и 36°, и Ph. megacephala из экваториальной Африки. Эти два вида обитают в разных зонах: Ph. megacephala - в теплых низменных местах, a I. humilis - в более прохладной местности в горах, на высоте больше 1000 м; такое распределение свидетельствует о том, что окончательный выбор местообитания зависит от того, откуда происходит данный вид и к каким условиям он приспособлен. Как указывают Flucker и Beardsley (1970), Ph. megacephala исчезает из той зоны, куда распространяется I. humilis, но позднее может вернуться, причем нередко за ним следуют другие, субдоминантные муравьи, например виды Tetramorium или Solenopsis geminata - единственный вид, основывающий в этой зоне семьи; I. humilis нападает на его гнезда и иногда затрачивает две недели на то, чтобы истребить Solenopsis, причем продвигается примерно на 30 м в глубь их территории. На юге США 5. invicta задержал распространение I. humilis на север (Brown, 1973). Ph. megacephala, защищаясь от нападения, цепляется за ноги и усики врага, удерживая его до прибытия солдат, которые разрывают его на части. Рабочие особи Ph. megacephala хотя и мельче, но агрессивнее рабочих I. humilis - те становятся агрессивными лишь в том случае, если на них нападают. Anoplolepis longipes (Formicinae) на Гавайских островах строит гнезда только под большими камнями и в канавах; он менее агрессивен, чем I. humilis и A. longipes, хотя и способен выбрасывать направленную струю муравьиной кислоты.
На Бермудских островах дело обстоит сложнее (Lieberburg et al., 1975). Pheidole megacephala появился здесь в конце XIX века и распространился по всей территории; затем в середине XX века прибыл Iridomyrmex humilis, который начал в свою очередь быстро расселяться; ожидалось, что он через полвека вытеснит Ph. megacephala и займет его место, однако продвижение I. humilis к 1959 г. замедлилось (Crowell, 1968), и в некоторых районах Ph. megacephala теперь вновь завоевывает территорию. I. humilis вначале расселяется путем отделения групп, состоящих из рабочих и самки; это происходит периодически, и во время фазы экспансии муравей более агрессивен, чем обычно. Можно видеть, как колонны мигрирующих рабочих и самок вторгаются в области, где обитает Ph. megacephala. Солдаты Ph. megacephala поднимаются на поверхность гнезда, муравьи хватают друг друга челюстями, и борьба продолжается несколько недель. Помимо этих обычных для муравьев действий Lieberberg и его сотрудники описывают применяемую I. humilis химическую атаку: эти муравьи направляют кончик брюшка на головы рабочих особей Ph. megacephala и за несколько секунд ошеломляют их, несомненно с помощью знаменитого терпеноидного лактона иридомирмецина (Cavill, Locksley, 1957). Подобное химическое воздействие представляет собой, вероятно, лишь одну из особенностей фазы экспансии I. humilis. Таким способом I. humilis может вытеснить Ph. megacephala, но его распространение лимитируется ростом численности популяции, который в свою очередь зависит от адаптации к климату и местообитанию. Следует упомянуть интересный факт, который может помочь объяснить взаимоотношения между этими видами: в то время как Ph. megacephala отыскивает и охраняет крупные приманки, I. humilis этого не делает.
На восточном побережье Африки (в Танзании) в свое время Way (1953, 1954), а позднее Vanderplank (1960; обзор: Brian, 1965) изучали сообщество муравьев кокосовой плантации того же типа, что и на Соломоновых островах. Oecophylla longinoda и Pheidole (на этот раз Ph. punctulata) были здесь местными видами, a Anoplolepis longipes - завезенным видом. Здесь О. longinoda подвергается нападениям со стороны A. longipes и Ph. punctulata, а также другого вида Anoplolepis - A. custodiens, который проявляет также враждебность к A. longipes. Муравьи A. custodiens просто хватают О. longinoda по одному и убивают их, обрызгивая ядовитой струей, когда они скапливаются для обороны своего дерева у его основания; но там, где наземная растительность затеняет почву, A. custodiens, по-видимому, не могут даже выжить, не говоря уже о том, чтобы нападать на О. longinoda. Муравей Ph. punctulata, очевидно, не умеет справляться с О. longinoda так, как справляется Ph. megacephala с О. smaragdina на Соломоновых островах. Он строит гнезда у подножия деревьев в почве или в щелях и уничтожает самок О. longinoda и их небольшие зарождающиеся семьи. Рабочие особи просто вцепляются в этих муравьев, удерживая их, пока солдаты разрезают их на части (но только в сухую погоду!). Рабочие О. longinoda могут оказаться также жертвами некоторых видов Crematogaster. Таким образом, вид О. longinoda хорошо приспособлен к жизни на кокосовых пальмах, но подвергается давлению со стороны многих других муравьев. Room (1971) сравнивал распределение видов на разных кокосовых плантациях в Гане и обнаружил 108 видов, живущих на одних только кокосовых пальмах. Хотя это старые деревья, они образуют не вполне сомкнутый полог, и пятна солнечного света попадают на траву, покрывающую легкую суглинистую почву. Большинство видов устраивает гнезда в мертвой древесине на земле, но 10 видов обитают в пологе, в том числе Oecophylla longinoda, Crematogaster striatula, Cr. africana, Cr. clariventris и Cr. depressa. Для них характерно пятнистое распределение, причем их семьи взаимно исключают друг друга, и в разных местах доминирующее положение занимают разные виды. Ни один из них не элиминирует другие виды полностью, так как они слегка различаются по потребляемой пище, хотя все собирают падь и используют для постройки гнезда один и тот же материал (картон), за исключением муравьев О. longinoda, которые устраивают гнезда из листьев, соединяя их шелком. Солнечного тепла, доходящего до поверхности почвы, недостаточно для того, чтобы гнездящиеся в земле виды могли создать мощную популяцию, способную конкурировать с этими древесными муравьями. У каждого доминантного вида есть набор ассоциированных с ним видов, но только два вида действительно тесно связаны между собой. Самое удивительное из этих субсообществ то, в котором доминирует вид Oecophylla longinoda в ассоциации с Crematogaster clariventris; эти два вида ассоциированы на 90%. Их взаимоотношения истинно парабиотические: территория Cr. clariventris включена в территорию О. longinoda, и его особи так тесно связаны с семьей, в которой они живут, что О. longinoda не нападает на них, хотя нападает на особей этого вида, принадлежащих к другим семьям. Как полагают, присутствие О. longinoda обеспечивает Cr. clariventris защиту от других муравьев, однако не ясно, что получают муравьи О. longinoda; быть может, Cr. clariventris отыскивают для них пищу?
В одной очень неоднородной области сомкнутость крон сильно варьировала - от весьма разреженного полога, подходящего для Camponotus acvapimensis, до очень плотного, где нашел приют Tetramorium aculeatum. После того как Majer (1976а-с) удалил гнезда доминантных видов, Т. aculeatum - моногинный, хотя и полидомный вид - распространился в местах с более разреженным пологом; это означает, что обычно его сдерживает давление со стороны других муравьев. Подобным же образом полигинный вид Crematogaster striatula распространяется на территории, занятые муравьем Oecophylla longinoda, только после того, как гнезда последнего будут срезаны. Crematogaster depressa переселяется на большую часть шоколадных деревьев после удаления с них доминантных видов, т. е. требования, предъявляемые этим видом к местообитанию, не очень специфичны. Вид О. longinoda, так же как и подчиненные виды, к такому переселению неспособен. Как показывают эти эксперименты, каждый вид-это часть системы доминирования, обладающей гибкостью и чувствительной к состоянию данного местообитания.
При обилии пищи агрессивное поведение наблюдается очень редко, так как конфронтации муравьи обычно избегают. Драки начинаются лишь при недостатке пищи. Однажды муравьи Oecophylla longinoda захватили дерево, которое занимал один из видов Crematogaster, и стали облизывать щитовок, которых Crematogaster не использовал; последний отступил при этом в листву, а вид О. longinoda приобрел источник пади. Этот пример нельзя считать конкуренцией за пищу; это просто перераспределение территорий, которое улучшает использование ресурсов в целом и, вероятно, способствует стабилизации сообщества.
Можно привести и другой пример. Crematogaster clariventris зашел на территорию, принадлежащую Cr. depressa, и вступил с ним в схватку на стволе тенистого дерева (не шоколадного) на расстоянии не менее 14 м от гнезда О. depressa и 9 м от гнезда О. clariventris. Борьба продолжалась два или три месяца; в конце концов Cr. depressa разбил агрессора, вторгся на его территорию и захватил его гнездо. Majer тоже описывает весьма показательное событие: дерево, которое использовал вид О. depressa, упало на территорию, занятую муравьями Oecophylla longinoda, которые включили его в свои владения. Majer высказывает мнение, что эти сообщества метастабильны. Территории, принадлежащие семьям О. longinoda, пластичны, подобно территориям Formica polyctena и Tetramorium caespitum, и благодаря этому на них могут обосновываться также и подчиненные виды. Хорошим примером такой "подгонки", приводящей к ослаблению трений, служит сдвиг периода активности у Tetramorium aculeatum, позволяющий этому виду избегать столкновений с Crematogaster depressa. Сообщают, что Т. aculeatum избегает таким же образом встреч с Cr. clariventris (Aryeetey; см. Leston, 1973); этот вид, очевидно, обладает достаточной гибкостью.
Изучая другое сообщество, обитающее на шоколадных деревьях в Гане (6° с. ш.), Majer использовал радиоактивный фосфор, чтобы определять форму и размеры семей Crematogaster. Как правило, виды этого рода занимают здесь четко обособленные территории; случаи совместного обитания с Oecophylla longinoda редки. Если О. longinoda и Tetramorium aculeatum используют одно и то же дерево, то их территории строго разграничены. В этой области О. longinoda - самый многочисленный муравей, составляющий 71% всей биомассы муравьев, хотя он использует только 12% деревьев (конечно, этот вид отличается крупными размерами). Второе место по численности принадлежит Crematogaster striatula, хотя он составляет только 14% всей биомассы муравьев (по сырому весу). В другом месте соотношение этих двух видов по частоте было противоположным: 73% Cr. striatula и только 3% О. longinoda.
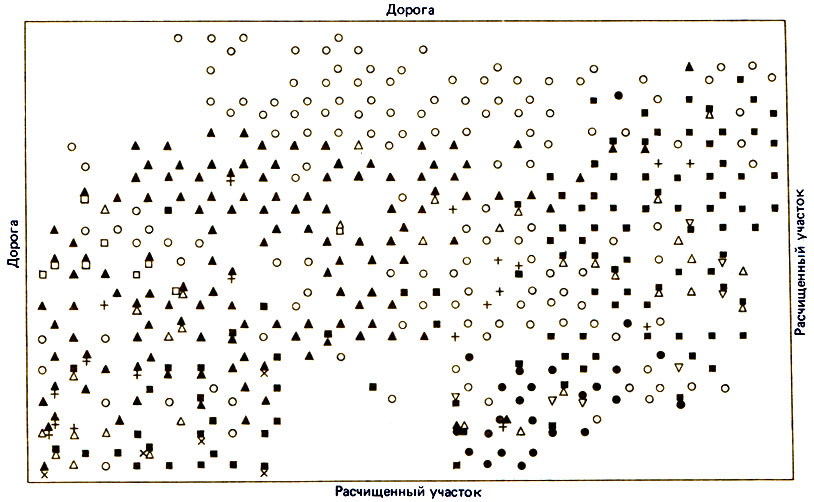
Рис. 16.8. Распространение муравьев на шоколадных деревьях на плантации недалеко от Ибадана (Нигерия). Белые кружочки - на дереве не обнаружено никаких муравьев; темные треугольники - Oecophylla longinoda; белые треугольники острым углом вверх - Pheidole megacephala; темные кружочки - Crematogaster clariventris; темные квадратики - Acantholepis capensis; белые квадратики - Crematogaster sp. А; белые треугольники острым углом вниз - Crematogaster acvapimensis. О распределении второстепенных видов см. Taylor, 1977
В Нигерии имеется сходный набор доминантных видов (рис. 16.8). При обследовании 3800 деревьев на 33% из них были найдены муравьи Oecophylla longinoda, на 14% - Tetramorium aculeatum, на 13% - Pheidole megacephala, на 12% - Crematogaster africana или Cr. depressa; привязанность видов к определенным деревьям была выражена слабо (Taylor, Adedoyin, 1978). Вид О. longinoda встречался чаще всех остальных видов на кофейных и шоколадных деревьях, но отсутствовал на деревьях с неподходящими листьями - слишком крупными, как у бананов, слишком узкими, как у гвинейской масличной пальмы, слишком мелкими и кожистыми, как у анакардии. Этот муравей очень плохо переносит дождь и ветер, а также, конечно, опадение листьев в сухой сезон. Crematogaster striatula часто встречался на кофейных деревьях и на деревьях кола, но редко - на шоколадных; возможно, что это было связано с присутствием на них Pheidole megacephala. Пазухи листьев масличной пальмы служат прекрасным местом для гнезд Ph. megacephala и Acantholepis capensis, хотя обычно эти два вида строят гнезда в земле. Оказалось также, что очень важную роль играет стадия развития плантации. Для некоторых видов сомкнутость полога не имеет значения, а для других она является важнейшим лимитирующим фактором. К стенотопным видам относятся: Tetramorium aculeatum и Crematogaster clariventris в густом пологе; Cr. gabonensis в старых деревьях с чешуйчатой корой; Acantholepis capensis и Camponotus acvapimensis в местах с разреженным пологом, где солнечные лучи доходят до земли. Эвритопный вид Pheidole megacephala широко распространен на земле и на деревьях. Taylor (1977) установил, что на 100 обследованных им местных дикорастущих лесных деревьях преобладали Oecophylla longinoda и Crematogaster africana (46%), Camponotus acvapimensis, Camponotus vividus и Cataglyphis guineensis (44%); оставшиеся 10% составляли другие виды. О. longinoda и Crematogaster подбирают мертвых муравьев и другие остатки, после того как пройдут Dorylinae (Leroux, 1979b).
Фауна, ассоциированная с доминантными видами муравьев, обитающих на шоколадных деревьях в Гане, включает 85 видов растительноядных сосущих насекомых, жуков, мух, бабочек и прямокрылых, из которых 54 встречались часто. Для каждого из этих видов были определены такие критерии, как сомкнутость крон, затененность, расстояние до опушки, расстояние до леса, сезон развития листьев и доминантный вид муравьев. Наибольшее число ассоциированных видов отмечено у следующих муравьев (в порядке убывания): Crematogaster striatula, Tetramorium aculeatum и Oecophylla longinoda. Это означает, что последний вид обитает, с точки зрения человека, на самых "чистых" деревьях, а первый - на самых заселенных. Муравей О. longinoda, будучи хищником, в то же время культивирует щитовок и потому не зависит от других насекомых и может истребить их, тогда как Т. aculeatum, в основном питающийся насекомыми, не может обходиться без них. Majer (1978) указывает, что при опрыскивании деревьев для борьбы с клопами-слепняками деревья, на которых обитают О. longinoda, обрабатывать не следует. Соблюдение интересов этого муравья не противоречит интересам человека.
На плантациях шоколадного дерева в Камеруне та же группа доминантных видов мозаично распределена в листве деревьев, но не доходит до уровня земли. В подстилке обитает около 80 видов, образующих иерархическую систему доминирования, в соответствии с которой между ними распределяются места для устройства гнезд и пища, но эти виды не могут образовать мозаику: эта возможность, несомненно, зависит не только от организации сообщества муравьев, но и от строения дерева. Кроме того, травам постоянно наносит ущерб человек, скашивая их или обрабатывая гербицидами (Jackson, 1982).
Сложность муравьиных сообществ показал Samways (1981), изучавший их на южноафриканских цитрусовых плантациях, где велась борьба с щитовкой. Когда прекратили обработку паратионом, разнообразие муравьев возросло и возникла мозаика доминантных форм. Спустя несколько лет это привело к доминированию одного муравья - Pheidole megacephala, который, хотя он и питается падью (в отличие от некоторых вытесненных субдоминантов), ограничил свои интересы злаками и другими травами, растущими между деревьями, и не наносил вреда цитрусовым. Samways приходит к выводу, что "очевидно, независимо ни от каких внешних влияний иерархия видов муравьев относительно постоянна и прочна... [и]... по-видимому, представляет собой одну из основных черт биологии сообществ муравьев".
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'