12.2. Копуляция и расселение
Расселение имеет два аспекта -генетический и экологический. Первый отражает необходимость обеспечить аутбридинг (по крайней мере периодически), а второй - необходимость находить и занимать новые местообитания, если они свободны, или вторгаться в уже занятые местообитания, вытесняя другие виды. Обычная последовательность событий такова: скопление половых особей, образование пар и копуляция, расселение и выбор местообитания. Существует много вариантов, особенно у тех общественных насекомых, у которых отпочковываются небольшие, но полноценные новые семьи. Копуляция между сибсами в гнезде или на гнезде наблюдается редко, и существует много механизмов, препятствующих этому. Гибельные последствия инбридинга были продемонстрированы на медоносной пчеле при помощи искусственного осеменения. Инбредные рабочие особи плохо выживают, запасают меньше меда и менее эффективны в отношении терморегуляции. В свете гипотезы о том, что гапло-диплоидные системы менее устойчивы к инбредной депрессии, чем диплоидные, эти данные заслуживают внимания (Bruckner, 1980). Механизм определения пола тоже может быть нарушен. У Apis имеется только один половой локус, но несколько половых аллелей; гетерозиготность по этому локусу дает самок, а геми - или гомозиготность - самцов. При инбридинге вероятность появления гомозигот возрастает, а тем самым повышается и частота диплоидных самцов, которые в репродуктивном отношении уступают гаплоидным. Рабочие особи распознают их на стадии личинки и уничтожают в однодневном возрасте, точно так же как они распознают репродуктивных самок, оказавшихся не в своих ячейках (Woyke, 1980a,b).
Самцы Polistes пытаются копулировать в гнезде, но самки их отвергают, а рабочие особи прогоняют. Самцы собираются в тех местах, где самки фуражируют перед началом зимы, налетают на них, падают на землю и копулируют в течение короткого времени (например, P. fuscatus; Evans, West-Eberhard, 1973). В отличие от этого самцы тропических P. canadensis приходят в новые гнезда и копулируют с самками, которые уже начали строительство без участия рабочих особей. Так обстоит дело с роящимися Polybiini вообще; при этом обеспечивается аутбридинг и снижается риск для репродуктивных самок. Самцы Vespula летают по маршрутам между участками растительности, хотя они не оставляют ощутимого запаха (Calam, 1969); после копуляции молодые самки уходят на зимовку и только после этого разлетаются и ищут места для гнезд. У самцов Bombus тоже есть маршруты между рядами столбов или древесных стволов, определенно помеченных секретом их мандибулярных (Calam, 1969) или лабиальных желез (Bergström, 1975; Bergström et al, 1981). Эти секреты, несомненно, помогают привлекать самок, и они видоспецифичны; у В. pratorum это терпеноидный спирт с 15 атомами С, такой как фарнезол, у В. pascuorum - алифатический спирт 7-гексадецен-1-ол, у В. lapidarius - пальмитолеиловый спирт 9-гексадецен-1-ол, а у В. lucorum, представленного двумя формами, светлой и темной, - этиловые эфиры: этилмиристолеат у светлой и этилдодеканоат у темной формы. Самки посещают места, помеченные секретами самцов, копулируют, возвращаются в гнездо, а позднее уходят на зимовку. Помимо использования различных химических меток разные виды занимают различные ярусы растительности.
Meliponini роятся круглый год или, как африканская Hypotrigona, не делают этого только в середине засушливого сезона (Darchen, 1977). Процесс роения состоит из ряда этапов. Сначала темные старые сборщицы меда подготавливают и выстилают полость, затем приходят молодые, устраивают горшочки для хранения запасов и наполняют их медом и пыльцой, и наконец является самка с целой толпой молодых рабочих особей, которые строят ячейки для расплода. Самцы держатся вокруг новых гнезд, устраиваясь поблизости и готовые копулировать, когда спустя два дня самка выходит из гнезда; сразу после этого она начинает откладывать яйца. Эта система, видимо, уменьшает инбридинг, так как в копуляции могут участвовать самцы со всей округи; кроме того, она снижает риск, которому подвергается новая матка во время перемещений и во время копуляции. Вероятность того, что семье не удастся обосноваться, невелика, так как выбранное место хорошо обследуется, прежде чем туда перейдет главная часть новой семьи. В сущности, эту систему можно считать почти идеальной, так как она обеспечивает самке защиту все время, кроме короткого периода копуляции, и при этом создает возможность для конкуренции между чужими самцами.
Медоносная пчела подвергается гораздо более многочисленным опасностям, так как старая матка покидает гнездо с группой рабочих особей, переполненных медом, но ее не ожидает заранее подготовленное гнездо. В старом гнезде остаются ее враждебно настроенные друг к другу дочери, а прежде чем начинать восстановление старого сообщества, необходим рискованный брачный полет (Simpson, 1974). Приспособленность матки в эволюционном смысле зависит от максимизации числа и качества производимых ею дочерей. Чтобы добиться такой максимизации, она сначала создает весной как можно быстрее большое число рабочих особей (это наблюдается у рас умеренного пояса), а затем, пока рост еще продолжает оставаться на максимальном уровне, а скученность или неадекватное распределение корма еще не стали серьезной помехой для жизнедеятельности семьи, строит трутневые ячейки и маточники и откладывает в них соответствующие яйца. Это связано с увеличением длины дня, наступлением тепла и обилием цветов. Возможно, что непосредственной причиной строительства трутневых ячеек и маточников служит снижение концентрации регуляторных феромонов матки из-за высокой численности рабочих особей и общей ареактивности молодых рабочих, преобладающих в популяции после периода быстрого ее роста.
Один из факторов успеха состоит в выборе для размножения наилучшего времени года. Это дает возможность старой матке основать новую семью, подготовить гнездо, создать популяцию и запасти достаточно меда, чтобы пережить зиму. Роение происходит главным образом в мае или июне (рис. 12.9), так что на все это остается примерно столько времени, сколько может потребоваться в обычное лето, поскольку дни длинные и большая часть растений находится в цвету. Все же в некоторых случаях рои возникают и в августе, но тогда новую семью создать не удается. Возможно, что такие рои - рудимент прежнего тропического цикла, длившегося около трех месяцев.
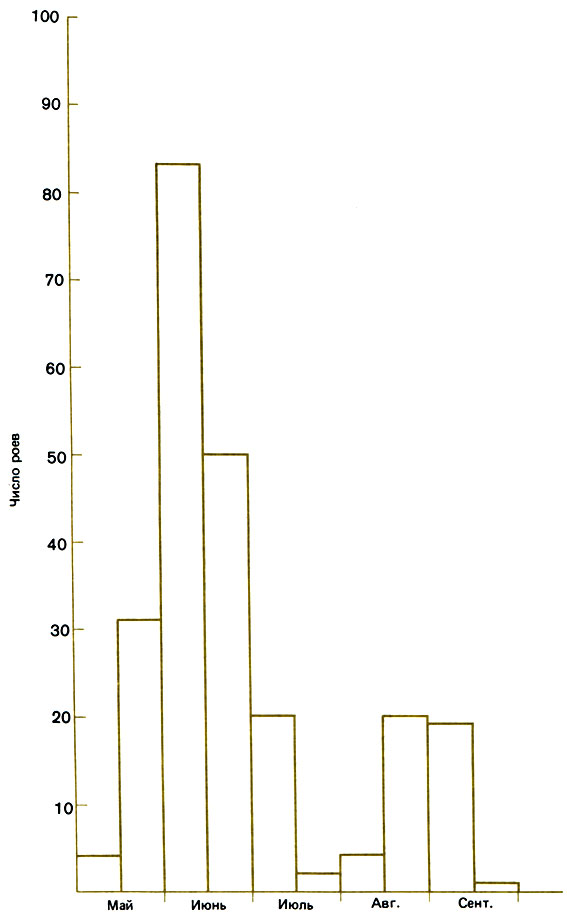
Рис. 12.9. Сроки вылета роев у медоносной пчелы, по суммарным данным за 5 лет в Нью-Йорке, США. (Fell et al., 1977.)
Другой фактор, от которого зависит успех, - это, вероятно, выведение оптимального числа новых репродуктивных самок. Их должно быть столько, чтобы им была обеспечена достаточная поддержка рабочих особей, т. е. чтобы образующийся рой имел оптимальные размеры. Чем больше рой, тем быстрее он может построить новое гнездо и обосноваться в нем, создав соты, яйца, расплод и запасы, но если рой слишком велик, то это будет за счет уменьшения других роев той части семьи, которая осталась в старом гнезде. По-видимому, одна семья может и должна отпочковывать от себя ежегодно по две или три новых семьи. Лишь одну из них основывает осемененная матка в возрасте более года, остальные закладываются молодыми самками.
Как же все это организовано? По крайней мере за 10 дней до роения рабочие пчелы начинают поглощать мед, готовясь покинуть улей вместе с маткой. Указывалось, что рабочие особи, уходящие с маткой, составляют более или менее случайную выборку из популяции (Butler, 1940), но наличие такой подготовительной фазы означает, что решение покинуть улей или остаться принимается каждой пчелой не спонтанно: некоторые рабочие особи по той или иной причине более преданы матке, чем другие. Быть может, у них существует какой-то способ распознавания генетического родства? Следует ожидать, что склонность к выработке воска для строительства ячеек и последующему развитию кормовых желез для кормления личинок будет сильнее выражена у молодых пчел; так обстоит дело у Apis mellifera adansonii, у которой более четырех пятых составляют особи моложе 8 дней (Winston, Otis, 1978).
Рассмотрим сначала, что происходит в улье, который зрелая матка оставляет в наследство своим дочерям. Молодые самки настроены друг к другу враждебно; самка, вышедшая из ячейки первой, старается ужалить более молодых и в случае успеха возглавляет новую семью. В других случаях вмешиваются рабочие особи, которые исполняют умиротворяющие танцы и добавляют к маточникам воск, чтобы помешать выходу молодых самок, особенно в перенаселенных семьях (Simpson, 1974); время от времени эти самки получают корм через дырку в воске и созревают внутри ячейки, а их кутикула твердеет. В период такого одиночного содержания репродуктивных самок они издают свое знаменитое прерывистое "чириканье" с частотой 650 Гц, создаваемое вибрацией сложенных крыльев, когда тело прижато к восковому субстрату. Это "чириканье" стимулирует роение и характерно для больших семей с высокой плотностью, которые способны образовать несколько жизнеспособных дополнительных роев. Если после выхода из ячеек молодых самок дополнительные рои не возникают, то эти самки уничтожают друг друга, пока не остается только одна из них, хотя сомнительно, чтобы это была наиболее приспособленная. Возможно, что происходят своего рода "предвыборные дебаты", в которых репродуктивные самки соревнуются, пытаясь склонить на свою сторону рабочих особей, как это делают молодая самка и матка у Trigona subnuda (Imperatriz-Fonseca, 1977).
Иногда матка не покидает гнездо вместе с роем, а остается в нем, и ее заменяет одна из ее дочерей. В таких случаях строится меньше маточников, и все выглядит так, как будто дело заранее планировалось как замещение матки, а не как роение - быть может, из-за того, что сезон подходит к концу или что матка плохо справляется с откладкой яиц (Butler, 1957). Наименее рискованный способ замещения для дочери состоит в том, чтобы сначала копулировать, а затем устранить старую матку, а для матери, безусловно, лучше передать гнездо осемененной дочери, чем рисковать полной катастрофой в результате несчастного случая при брачном полете (Simpson, 1974).
При наличии соответствующих условий - как внешних (длинные дни и ясная теплая погода), так и внутренних (запечатанные маточники) - некоторые пчелы, более активные и чувствительные, начинают исполнять в улье активизирующий танец (buzz-dance), который состоит из очень быстрых пробежек по прямым линиям при частично раскрытых крыльях, вибрирующих с частотой 250 Гц. Когда одна пчела соприкасается с другой, она сохраняет с ней контакт в течение 5 с и непрерывно (а не с перерывами, как матка) "чирикает" с частотой 500 Гц. Это побуждает другую пчелу к активизирующему танцу, но только в том случае, если она находится в состоянии готовности; если такой танец исполняется в нероящемся улье, то пчелы "замирают" (Esch, 1967). В готовом к роению улье реакция распространяется до тех пор, пока эмигрирующие пчелы не высыпают наружу все вместе, причем гораздо быстрее, чем обычно, и образуют плотное облако вблизи улья.
Пчелы не направляются прямо к новому гнезду, а собираются на расстоянии до 10 м от улья, оставаясь на месте до тех пор, пока там присутствует матка, а в противном случае возвращаются в улей. Распознавание матки - процесс сложный; Ferguson et al. (1979) стабилизировали безматочные рои при помощи синтетического феромона, сходного с секретом железы Насонова, который содержит цитраль, нерол, гераниол и фарнезол, а также 9-оксодекановую кислоту и другие компоненты из мандибулярной железы матки. Такой сложный состав имеет важное значение, поскольку Ambrose et al. (1979) показали, что роящиеся пчелы могут отличать свою собственную матку от других маток и выбирают матку, сходную по возрасту и репродуктивному состоянию со своей, не принимая в качестве заместительницы матку, которая отличается от прежней. Они даже способны идентифицировать собственную матку, если экспериментатор пометит ее шеллаком (Boch, Morse, 1979).
Найдя новое место для гнезда, пчелы-разведчицы (примерно 5% пчел роя) указывают своим товаркам направление к этому месту и расстояние до него, используя те же танцы, что и для сообщений о корме. Если имеется несколько возможных мест, то, по-видимому, разведчицы посещают каждое из них по отдельности и оценивают их. Только после того, как выбор будет одобрен всеми, роевой клуб готов отправиться в путь, но пчелам требуется еще некоторое время, чтобы взлететь в воздух. Seely et al. (1979) установили, что примерно за час до того, как рой взлетает, разведчицы начинают собираться на роевом клубе: некоторые сохраняют неподвижность, другие исполняют информационные танцы, третьи - активизирующий танец. Этот последний танец индуцирует высокие чирикающие звуки, и цепи висящих пчел распадаются, тогда как другие пчелы взлетают и начинают описывать круги в воздухе, и в конце концов все отправляются в нужную сторону. Пчелам, вероятно, помогают в этом разведчицы, летающие взад и вперед вдоль нужного направления, в то время как другие разведчицы летят к месту будущего гнезда, выделяя по пути феромон, секретируемый железой Насонова. Скорость их полета увеличивается, достигая 8-10 км/ч при высоте 3 м, и по мере движения рой уплощается.
Местообитание, выбираемое роями диких пчел, - это обычно редкий лиственный лес, в котором можно найти старые деревья с дуплами для гнезд и разнообразный корм. Место для нового гнезда редко бывает удалено от старого менее чем на 300 м, и иногда расстояние достигает километра (рис. 12.10); пчелы предпочитают устраивать гнезда в полостях объемом около 40 л на высоте 1-5 м, с расположенным у основания отверстием величиной 12-75 см2, обращенным на юг (Seely, Morse, 1977, 1978). Разведчицы обследуют найденное место в течение 40 мин и, возможно, определяют в темноте объем полости, сопоставляя расстояния, которые они проходят в ней в разных направлениях (Seeley, 1977). Avitable et al. (1978) находили большую часть диких семей на высоте более 3 м в толстых зрелых кленах (Acer) и дубах (Quercus) - B гнездах, обращенных на юго-запад.
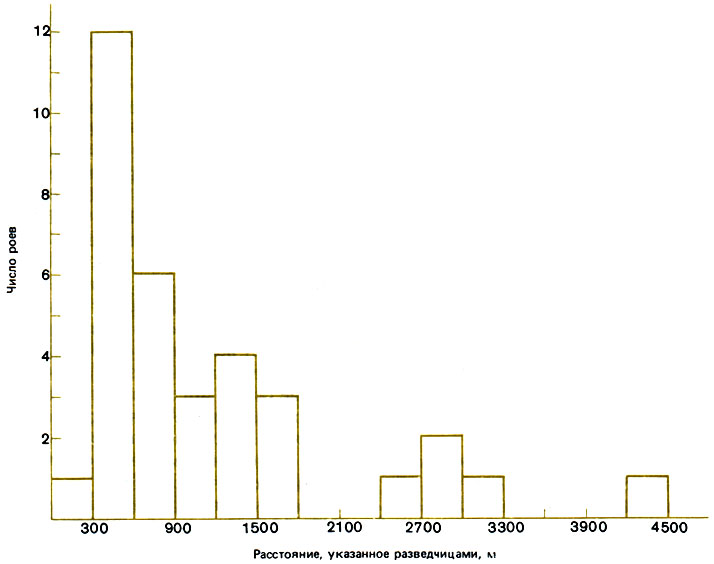
Рис. 12.10. Расстояние от скопления пчел до места нового гнезда, указанное роящимися пчелами: первые столбики в каждой паре - по Seeley, Morse (1977), вторые - по Lindauer (1955)
Поскольку самцы не прилетают к гнезду, как у безжалых пчел, молодым самкам приходится выходить из гнезда, чтобы отыскивать их и копулировать. Самцы летают и собираются в тех местах, где сходятся восходящие воздушные потоки, поднимающие их примерно на 10 м над землей (без подзорной трубы их увидеть нельзя, но можно слышать). У Apis mellifera adansonii молодые матки в хорошую погоду начинают 1-2-минутные ориентировочные полеты вокруг гнезда через 3-4 дня после выхода из ячейки. Они копулируют на 5-й день, а некоторые повторяют копуляцию и на следующий день; эти полеты продолжаются по 13 мин и происходят по большей части днем, около 15 часов (Fletcher, 1978). Во время полета матка выделяет из своей мандибулярной железы след из привлекающих веществ, которые воспринимаются особыми клетками на усиках самцов. Самец воспринимает 9-оксодекановую кислоту в очень сильном разведении, что дает ему возможность подлететь к самке против ветра с большого расстояния. Однако сейчас еще не выяснено, каким образом репродуктивная самка находит скопления самцов. После копуляции запас сперматозоидов в сперматеке достигает 5-6 млн., а после откладки 100000 яиц число их снижается до 2-3 млн., т. е. в среднем на каждое яйцо приходится 30 сперматозоидов. В каждое яйцо может проникать до 10 сперматозоидов (Harbo, 1979).
У муравьев существует много типов репродуктивного поведения. У некоторых Ponerinae и у всех Dorylinae самки бескрылые и проводят в семье всю свою жизнь. Крылатые самцы отыскивают самок, следуя по фуражировочным тропам, как, например, у Megaponera foetens (Longhurst, Howse, 1979). У Eciton (Dorylinae) в расплоде бывают тысячи самцов, но лишь несколько репродуктивных самок. У этих муравьев при разделении семьи образуются две части - одна со старой самкой, а другая с расплодом. Между этими двумя частями проявляется некоторый антагонизм. Молодую самку, пока она еще не потемнела, окружает группа рабочих особей, которые пробивают себе путь наружу и образуют скопление на расстоянии около метра от гнезда; рабочие из этой группы, увеличивающейся и достигающей 8-10 см в диаметре, перемещаются вместе с основной частью семьи. Спустя несколько дней выходят из своих коконов самцы, и, вероятно, начинается то состояние возбуждения, которое Schneirla (1971) назвал "стадией исхода самки": на этой стадии старая самка выходит наружу вместе с рабочими особями и отправляется в путь по одной из троп. Спустя примерно час первая молодая самка, все еще в сопровождении рабочих, уходит из гнезда по какой-нибудь другой тропе, образуя отводок. Другие репродуктивные самки выходят из коконов слишком поздно, и им трудно привлечь к себе рабочих особей; при неблагоприятных условиях рабочие особи изолируют самок, окружая их плотным кольцом и держа за ноги. Тем не менее некоторым молодым самкам удается вырваться из этих скоплений и в конце концов добиться того, что их принимает какая-нибудь кочующая колонна, которая в таком случае отказывается от собственной самки; такая замена может произойти и без разделения семьи, если старая самка очень непривлекательна (ср. Арis). У Eciton самцы обычно улетают и стараются присоединиться к какой-нибудь колонне муравьев своего вида. В течение первого дня они приобретают запах новой для них семьи, теряют крылья, обламывая их самостоятельно или позволяя сделать это другим особям, а затем достигают половой зрелости. Самцы не могут копулировать, пока не сбросят крылья, а после того, как произойдет копуляция, они гибнут.
В саванне Ламто муравьи Аnоmmа (по данным Leroux, 1979а) обычно меняют гнездо каждые 8 дней, но когда они начинают размножаться, производя много самцов и несколько самок, они остаются на одном месте 2-3 недели, непрерывно занимаясь охотой. Некоторые семьи многократно делятся, а другие не делятся никогда; в течение 134 дней одна семья разделилась трижды, но из 46 семей, находившихся под наблюдением, разделились только 14. Поскольку самцы Dorylinae выслеживают семьи, принадлежащие к их собственному виду, и копулируют с избранными ими самками, серьезному риску гибели подвергаются у них только самцы. Активность самцов ограничена; в сухой сезон они часто прилетают в световые ловушки, но сроки лёта у разных видов различны; самцы летают во влажном воздухе после дождя (Baldridge et al., 1980). У Dorylinae, хотя у них самка не способна к полету, вряд ли происходит инбридинг, так как ее братья улетают раньше, отправляясь на поиски семьи, которая их примет. Рабочие особи защищают самку в течение всей ее жизни, и по сравнению с этим риск, которому подвергается самка Apis во время брачного полета, кажется огромным:, если эту последнюю уничтожит хищник, то погибнет вся новая семья, так как теперь уже слишком поздно, чтобы рабочие особи могли вернуться в прежнее гнездо и начать все сначала; невольно возникает мысль, не объясняется ли такая беспечность медоносной пчелы ее ассоциацией с человеком.
Бескрылым репродуктивным самкам не всегда приходится ожидать, пока к ним прилетит самец. Самки Rhytidoponera metallica (Ponerinae) выходят из гнезда и привлекают к себе самцов с помощью феромона, выделяемого межсегментными железами (рис. 12.11). У Нуроропеrа eduardi самцы бывают как крылатые, так и бескрылые, причем последние копулируют с самками вскоре после своего "рождения" (LeMasne, 1953). У Myrmicinae в процессе эволюции возникла выраженная в различной степени зависимость от других свободноживущих муравьев, и этот социальный паразитизм часто ассоциируется с бескрылостью репродуктивных самок и самцов. У Formicoxenus nitidulus есть как крылатые, так и бескрылые репродуктивные самки; последние выходят из гнезда, взбираются на какое-нибудь растение поблизости и, приподнимая брюшко, выпускают яд из кончика выдвинутого жала. Бескрылые самцы направляются к жалу с расстояния по меньшей мере 15 см, и после некоторого "жеманничания" со стороны самки происходит копуляция (Büschinger, 1975). Крылатые самцы, по-видимому, никогда не летают и уже в первые два дня обламывают свои крылья. Вероятно, инбридинг происходит достаточно часто, однако отсутствие крыльев у социальных паразитов встречается только среди Leptothoracini. Самка Harpagoxenus sublaevis взбирается на растения около своего гнезда и часами сидит на них, выдвинув жало с выпущенной из него капелькой яда; это привлекает крылатых самцов, находящихся на расстоянии 3-4 м. У Н. sublaevis бывают и крылатые самки, а у H. canadensis все самки имеют крылья (Büschinger, 1978b), у Anergates atratulus - паразита Tetramorium caespitum - самцы очень похожи на куколок и копулируют со своими сестрами вскоре после появления на свет (Büschinger, 1974). Такой теснейший инбридинг, вероятно, крайне вреден в генетическом отношении, однако А. atratulus - чрезвычайно специализированный социальный паразит, у которого нет касты рабочих, а самки после копуляции летят к другому гнезду T. caespitum. Описано много подобных случаев, и, как отмечает Wilson (1975), эти кажущиеся неблагоприятными особенности на самом деле дают возможность их обладателям не терять контакта с популяцией хозяев.
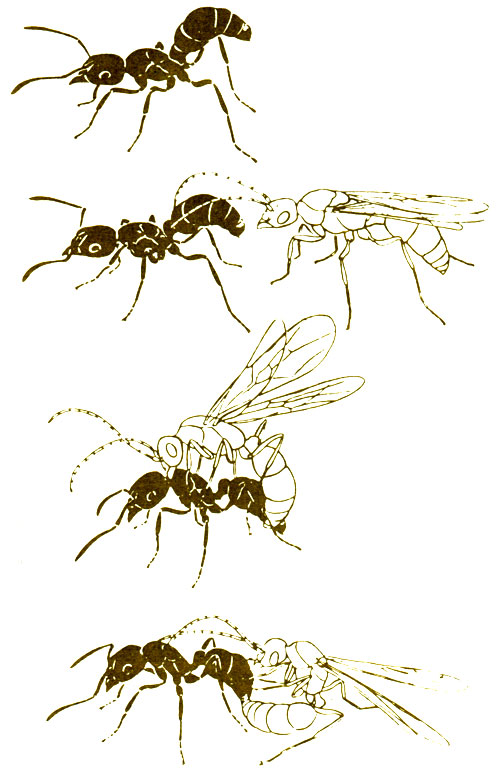
Рис. 12.11. Самка Rhytidoponera metallica привлекает самца, выделяя из тергальной железы, расположенной у кончика брюшка, особый феромон, после чего происходит копуляция. (Hölldobler, Haskins, 1977.)
В тех подсемействах муравьев, в которых репродуктивные самки большей частью летающие, оба пола обычно покидают гнездо и встречаются вне его. Расстояние, которое они при этом покрывают, варьирует в пределах от нескольких сантиметров до нескольких километров и может превосходно обеспечить расселение. У Myrmica, Pogonomyrmex и многих других муравьев самцы отправляются к какому-нибудь заметному объекту вроде скалы или дерева и летят тесно сбитым извивающимся роем на его подветренную сторону, где они падают на голую землю, чтобы копулировать. Они выделяют при этом феромоны, которые распространяются по ветру и помогают самкам найти это место. Разные виды могут использовать для этого из года в год одно и то же место, хотя и в разное время дня или года - вероятно, потому, что у него есть какие-то особенности, поскольку это не может быть связано у них с традицией, как постоянство токовищ у птиц. Быть может, прав Wynne-Edwards, предположивший, что скопления муравьев связаны с некоторым половым отбором. В результате отбора хорошо летающих самок преимущество получают особи с хорошо развитыми крыловыми мышцами, т. е. обладающие наилучшими запасами питательных веществ для одиночной жизни во время основания новой семьи. Самки Pogonomyrmex могут копулировать с несколькими самцами подряд, после чего они улетают, садятся, обламывают крылья и отправляются "пешком" на поиски места для гнезда (Hölldobler, 1976b).
Аттрактант самцов Pogonomyrmex близок к 4-метил-З-гептанону или соответствующему спирту (McGurk et al., 1966, цит. по Hölldobler, 1976). Ядовитая железа самок продуцирует феромон, который в сочетании с веществом, выделяемым покровами, а также со специфической морфологией и поведением самок помогает самцам, которых бывает по нескольку подряд, спариваться. Звуковые сигналы при спаривании не используются, кроме тех случаев, когда удовлетворенная самка не может уйти от самцов, - тогда она прибегает к стридуляции (Markl et al., 1977). Образование подобных скоплений самцов и самок зависит, во-первых, от примерно одновременного их созревания; во-вторых, от выхода из гнезда в подходящую погоду (обычно теплую, безветренную и влажную) и в определенное время дня, которое варьирует у разных видов ; в-третьих, муравьи должны лететь к какому-нибудь заметному ориентиру, выделяя характерный феромон; а после того как сформировался рой, он может привлекать к себе муравьев одним своим видом.
Многие муравьи, по-видимому, обходятся без каких-либо особых ориентиров, - они просто собираются на участках, отражающих свет или испускающих тепло; некоторые взлетают на определенную высоту и держатся на ней. Рабочие особи Solenopsis invicta открывают выходные отверстия гнезда, и самцы, вылетая из гнезд раньше самок, примерно в полдень, остаются на высоте 90-150 м в течение нескольких часов (Markin et al., 1971, цит. по Lofgren et al., 1975). Брачный лёт происходит несколько раз в году, чаще всего в июне и июле, иногда около гнезда, иногда в отдалении от него; обычно его наблюдают спустя 1-2 дня после дождя, особенно если дождю предшествовала сухая погода, при слабом ветре, относительной влажности 80% и температуре 24-32°С, но зимой муравьи могут летать при 20-26°С (Lofgren et al., 1975). Запоздалые брачные полеты могут пробить серьезную брешь в кормовых запасах семьи; поэтому многие муравьи уничтожают половых особей, не вылетевших из гнезда к концу сезона размножения. Кроме того, существует опасность, что самки, оставшись неосемененными, начнут все-таки откладывать яйца. Если эти яйца съедобны, то самки просто помогут семье; если же это будут репродуктивные яйца, в семье в неподходящее время появится слишком много самцов. У S. invicta, по-видимому, существует предохранительный механизм: яйцекладущая самка вырабатывает феромон, подавляющий сбрасывание крыльев молодыми самками в гнезде до копуляции. Если удалить старую самку, молодые самки сбрасывают крылья, пользуясь помощью рабочих особей, не позднее чем на следующий день. Несколько дней спустя рабочие начинают убивать множество таких только что сбросивших крылья самок, оставляя лишь нескольких, у которых хорошо развиты яичники. Новая "матка", хотя она не осеменена, способна прекратить дальнейшее сбрасывание крыльев другими самками, вероятно с помощью феромонов, вырабатываемых ею самой (Fletcher, Blum, 1981 a, b). Этот феромон покрывает тело самки (особенно брюшко), и его можно смыть ацетоном; он нелетучий, и рабочие особи либо получают его при непосредственном контакте с самкой, либо передают друг другу на ротовых частях. Ввиду наличия латентного периода в 1-2 дня между удалением самки и началом сбрасывания крыльев цитированные авторы высказали мысль, что это вещество служит "затравкой", проникающей в организм молодых самок и изменяющей их поведение путем прямого воздействия на нервную систему, а не на экстероцепторы. Эта ингибирующая система очень сходна с той, с помощью которой самки Myrmica останавливают превращение лабильных личинок в половых особей (Brian, 1970); достаточно поместить их на 30 с в эфир, чтобы смыть этот феромон.
Tetramorium caespitum, подобно Solenopsis, по-видимому, относится к видам, совершающим брачный полет на определенной высоте. В отличие от других видов, обитающих на верещатниках и в грасслендах, эти муравьи начинают летать, как только лучи всходящего солнца коснутся поверхности гнезда. Мандибулярная железа самца секретирует 4 метил-3-гексанон и соответствующий спирт; эти соединения имеются у рабочих особей, но их нет у репродуктивных самок (Pasteels et al., 1980).
Lasius и Acanthomyops при брачном полете тоже держатся в определенном слое воздуха и выделяют терпеноиды и индол в соотношении, различном у разных видов; возможно, что самцы сообщают о своем появлении при помощи этих веществ. Данных о высоте брачного лёта у разных видов пока нет, однако известно, что он происходит в разное время года: например, в Англии у L. niger - в июле и августе, а у L. alienus - в сентябре и октябре, часто даже слишком поздно, когда погода уже испортилась, и в таких случаях самцов поедают в гнезде собственные рабочие особи. У L. flavus брачный полет происходит в то же время года, что и у L. niger, но ближе к вечеру: в 17-19 вместо 13-17 часов (Boomsma, Leusink, 1981); эти авторы полагают, что каждый вид вылетает в то время, когда температура среды примерно равна температуре в гнезде, и избегают таким образом термического шока. Благодаря полетам на разной высоте при наличии или отсутствии ориентации по особенностям местности эти виды могут обитать на лишенных деревьев травянистых равнинах и в степях. Такие массовые полеты хорошо известны, так как они почти всегда привлекают всевозможных неспециализированных хищников: это не только птицы, хватающие муравьев в воздухе, но также рептилии и амфибии, млекопитающие и паукообразные, жуки, хищные клопы и другие насекомые, которые ловят муравьев среди растительности и на земле, во время копуляции или после нее. Высказывается мнение, что, хотя синхронизированные вылеты муравьев привлекают разнообразных хищников, такое массовое появление жертвы рассредоточивает атаку последних и позволяет немалой доле половых особей и значительному числу самок избежать гибели и приступить к закладке новых семей.
Другие муравьи тоже прибегают к уловкам бескрылых самок, чтобы завлекать самцов. Это относится ко всей группе Formica rufa; после копуляции, происходящей обычно около гнезда, самки возвращаются в него, а затем переходят в один из отпочковавшихся отводков, который временно сохраняет связь с родительским гнездом, но в конце концов либо резорбируется, либо отделяется* (Mabelis, 1979b; Pamilo, Varvio-Aho, 1979). Таким же образом репродуктивные самки Monomorium pharaonis используют секрет дюфуровой железы для привлечения самцов (Hölldobler, Wust, 1973).
*(Часто эта связь сохраняется постоянно, и образуются комплексы родственных гнезд, которые называют колониями (см. предисловие редактора перевода). - Прим. перев.)
Подводя итоги, следует сказать, что существуют два главных способа образования скоплений у муравьев: 1) самки летят к самцам, которые собираются около какого-нибудь хорошо заметного объекта или в зоне, находящейся на определенной высоте над землей; 2) самки привлекают к себе кочующих самцов с помощью "призывов", не удаляясь от собственного гнезда. Особый случай, наблюдаемый у Dorylinae, когда семья принимает чужого самца, после чего он осеменяет в ней самку, - это, возможно, вариант второго способа. Первый способ обеспечивает максимальную дисперсию генов и семей, а второй - достаточную дисперсию генов, однако ввиду того, что впоследствии семьи иногда разделяются или почкуются, расселение вида происходит при этом способе медленно, но зато надежно. В первом случае приспособленные самки - это самки с мощной летательной мускулатурой, что дает им возможность в одиночку основать новую семью. Во втором же случае это мелкие самки, возвращающиеся в гнездо и принимаемые собственной семьей или поселяющиеся в семьях других видов как социальные паразиты. Этим последним самкам не нужны ни дальние полеты, ни мощные летательные мышцы, и расселение во вновь появляющиеся незанятые местообитания не дает им никаких преимуществ. Лучше всего им оставаться около своих хозяев, чтобы их не сдувало, а наличие у них крылатых самцов может обеспечить дисперсию генов. Для тех видов, у которых даже самцы лишены крыльев и, появившись на свет, копулируют со своими сестрами, отсутствие риска потерять хозяина, возможно, перевешивает выгоду, связанную с аутбридингом.
Поспешность, с которой молодые самки обламывают свои крылья (если они у них есть) после копуляции, позволяет думать, что они не очень-то усердно ищут подходящее местообитание; во многих случаях они, по-видимому, уже нашли его раньше. В выборе местообитания иногда участвуют и самцы (как, например, у шмелей), но, вероятно, лишь в самой общей форме (Wynne-Edwards, 1962; см. Brian, 1965). Lasius, у которого самец так мал, что самка может нести его на себе, когда копуляция еще продолжается, служит примером поисков места для гнезда, производимом молодой самкой с воздуха. Судя по некоторым данным, самки действительно выбирают местообитание; например, у L. niger и L. flavus в Европе они предпочитают голые участки почвы на капустных полях (Pontin, 1960), а самки L. neoniger в Америке (луговой вид), пока они еще не сбросили крылья, избегают лесов, а случайно опустившись в лесистой местности, улетают прочь (Wilson, Hunt, 1966). Самки L. niger в Европе обосновываются в более влажных областях, чем самки L. alienus (Brian et al., 1966). Ha травянистых пустошах и в кустарниках самки Formica lemani, Myrmica ruginodis и M. scabrinodis выбирают открытые поляны или недавно расчищенные места в лесу и строят гнезда на хорошо прогреваемых участках и на южной стороне старых пней (Brian, 1965).
Брачные полеты термитов сходны с таковыми у летающих муравьев с той разницей, что они заканчиваются не копуляцией, а образованием пары; копуляция же происходит позднее, уже в гнезде, и время от времени повторяется. Очень немногие термиты активно летают, и не известно ни одного вида, который бы улетал за пределы относительно спокойных нижних слоев воздуха. Крылатые самки и самцы Allognathotermes hypogeus собираются на верхушках деревьев, разбросанных с плотностью примерно по одному на гектар; пары образуются в листве деревьев, причем самец прикрепляется к нижней поверхности тела самки, которая затем слетает на землю, быстро отламывает свои крылья и начинает бродить в поисках места для гнезда. К числу термитов, образующих брачные скопления, относится также Pseudacanthotermes militaris (Noirot, Bodot, 1964).
Самки Allodontotermes giffardi не летают, хотя у них и есть крылья. Они взбираются на травинки, подняв брюшко, и машут крыльями; этим они привлекают самцов, визуально или акустически, а к концу, несомненно, химическим путем (Bodot, 1967).
В отличие от муравьев, у которых рабочие особи просто открывают выходные отверстия, термиты строят специальные "взлетные" платформы и укрытия, охраняемые солдатами. Синхронизация достигается, во-первых, благодаря созреванию самцов и самок примерно в одно и то же время и, во-вторых, благодаря выбору определенной погоды и определенного времени суток. В обзоре на эту тему Nutting (1969) пишет, что обычно в областях с умеренным климатом брачный лёт у термитов происходит летом, а в пустыне - зимой, после дождя*. В тропиках не у всех видов лёт приходится на сезон дождей, у некоторых он происходит в засушливый период; это, по-видимому, частично зависит от того, в какого рода местообитании им предстоит рыть норку после брачного полета, потому что дождевая вода размягчает древесину и почву, но вместе с тем заполняет щели в почве. У некоторых видов и даже отдельных семей брачный лёт происходит несколько раз в год, что довольно выгодно для них (у Pterotermes до 40 раз, по данным Nutting). У некоторых видов время брачного лёта непредсказуемо, но у других его можно предсказать; например, у пяти видов Amitermes он происходит на следующий день после дождя, а у одного вида Heterotermes начинается через 10 мин после захода солнца и продолжается всего час! Пролетаемое термитами расстояние тоже сильно варьирует: так, у Macrotermes из нескольких тысяч вылетающих половых особей некоторые могут пролететь расстояние в несколько километров, а у Calotermes flavicollis, у которого в каждом лёте участвует по 30-60 половых особей из каждой семьи, они пролетают лишь по нескольку десятков метров.
*(У среднеазиатских термитов лёт происходит весной (Тутиков, 1979). - Прим. ред.)
Термиты способны в общих чертах распознавать тип местообитания. Nutting (1969) полагает, что виды, обитающие в пустыне, узнают по очертаниям объекты, которые могут предоставить им пищу и убежище. Некоторые термиты летят вдоль рядов деревьев или по улицам, как бы рассматривая их, например Amitermes в Аризоне. На территории Берега Слоновой Кости Amitermes часто поселяется вблизи холмиков Trinervitermes geminatus (Josens, 1972), и возможно, что он распознает их прежде всего зрительно. Формирование пар происходит на земле, причем самец следует за самкой, поддерживая с ней тесный тактильный и химический контакт. Например, самка Trinervitermes bettonianus привлекает самца, находящегося на расстоянии 10 см от нее, выделяя летучее вещество, а самец ощупывает тергальную область самки, помещая голову на область от 6-го до 10-го тергита. При утрате контакта тергальные и стернальные железы прокладывают эфемерные следы (Leuthold, 1977). У Zootermopsis nevadensis и Z. angusticollis, использующих для этого стернальные железы (Pasteels, 1972), самок привлекают вытяжки из особей обоих полов, однако самцов привлекают только вытяжки из самок; после образования пары термиты обламывают свои крылья. У примитивных термитов это достигается путем трения о какие-нибудь предметы, при помощи задних лапок или при участии брачного партнера, отгрызающего крылья у их основания. Высшие термиты ставят крыло перпендикулярно груди и приподнимают крыловые пластинки до тех пор, пока они не отламываются по слабому шву около основания (Nutting, 1969). У Microtermes самка переносит самца на нижней поверхности своего тела, как у Allognathotermes hypogeus. Самец сбрасывает крылья, а самка обследует местность с воздуха и, выбрав подходящее место, опускается и начинает устраивать гнездо.
Термиты, живущие в древесине, обычно проникают в нее через отверстия, проделанные жуками, или через различные трещины; чрезвычайно большое значение для них имеет состояние древесины - ее мягкость и степень разложения. Термиты, строящие гнезда под землей, часто используют временные укрытия, предоставляемые опавшей листвой и растительными остатками, но очень скоро начинают рыть вертикальный ствол; большую часть работы выполняет самка, а самец охраняет ее от самцов, не нашедших пары. В конце ствола термиты выкапывают камеру, которая иногда бывает обмазана экскрементами и так мала, что самка едва может в ней повернуться. Отверстие ствола закрывается перегородкой из экскрементов и разного рода обломков, различающейся по форме у разных видов. Macrotermes natalensis роет с помощью мандибул и отгребает вырытую землю ногами. Microtermes havilandi строит над входом небольшой купол из уплотненной почвы, причем вся эта операция занимает лишь около 10 мин, в течение которых самец и самка работают поочередно (Nutting, 1969).
Во время брачного лёта на термитов, так же как и на муравьев, нападают многие неспециализированные хищники. Миграции перелетных птиц могут быть приурочены к лёту термитов в тропиках весной и к лёту муравьев в умеренном поясе осенью. Парам половых особей необходимо построить как можно скорее надежные убежища, а не оставаться просто под слоем листьев, где они могут стать добычей жуков, пауков и птиц, роющихся в лесной подстилке. Хотя они должны держаться вместе, поскольку у них нет приспособления для хранения спермы, они все же не подвергаются такой опасности, как муравьи, копулирующие на открытых местах; кроме того, оставаясь вместе, они строят гнездо общими усилиями.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'