12.1. Морфогенез каст
12.1.1. Термиты и муравьи
У термитов (за исключением сем. Termitidae) рабочие особи мало чем отличаются от личинок, если не считать того, что их покровы несколько жестче и слегка пигментированы, а сами они более активны и более общительны. Развитие их протекает постепенно и не сопровождается какими-либо изменениями в строении зачатков крыльев или гонад. Линька создает возможность для роста и развития, хотя эти процессы имеют место не всегда, а "стационарная" линька означает, что число стадий развития может варьировать. Тем не менее существует ряд определенных типов развития. Например, у Mastotermes darwiniensis, самого примитивного из ныне существующих термитов, имеются пять личиночных стадий и одна стадия рабочего; на этой последней стадии особи окрашены темнее и занимаются фуражировкой. Но это не завершающая стадия, так как рабочая особь может перелинять и превратиться в нимфу с явно выраженными зачатками крыльев, а затем, после трех нимфальных стадий, стать крылатой половозрелой формой с функционирующей половой системой (мужской или женской). Таким образом, всего здесь 10 стадий. Даже нимфа не детерминирована к тому, чтобы непременно превратиться в имаго, так как она может встать на путь развития, приводящий к солдату. Для этого она должна сначала перелинять в промежуточную форму, называемую просолдатом; на стадии просолдата происходит перестройка, а затем, после новой линьки, получается взрослый солдат; у Mastotermes это просто более крупная бескрылая особь с массивными челюстями и недоразвитыми гонадами. Это превращение, подобно превращению в имаго, сопровождается утратой морфогенетических потенций: состояние солдата резко отличается от метастабильной стадии рабочего, который может оставаться рабочим, проходя "стационарные" линьки, или же, пройдя через нимфальные стадии, превратиться в половую особь или в солдата и закончить на этом свой жизненный цикл. В небольших семьях могут формироваться узкоголовые солдаты, возможно в результате преждевременного метаморфоза, стимулируемого царской парой (Watson, 1974). Для представителей семейства Calotermitidae (например, Calotermes, Zootermopsis) характерен такой же однонаправленный морфогенез; личинки старших возрастов выполняют различные работы, ожидая, когда настанет их очередь превратиться в имаго или солдат; при этом они проходят через несколько нимфальных стадий или через одну стадию просолдата (рис. 12.1). В этом семействе нимфы могут в случае необходимости превращаться в половых особей, бескрылых или с недоразвитыми крыльями, но вполне эффективных в смысле размножения. Нимфы могут также превращаться в солдат.
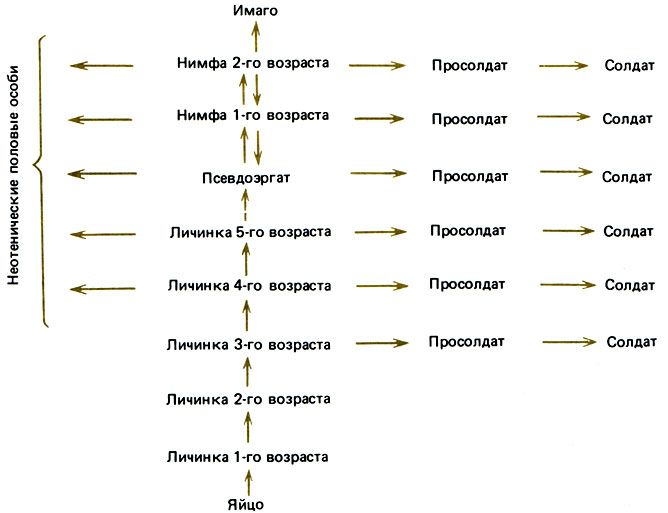
Рис. 12.1. Возможные пути онтогенетического развития у Calotermes flavicollis. Каждая стрелка соответствует одной линьке; пунктирная стрелка означает варьирующее число линек. (Ltischer, 1976.)
У Rhinotermitidae (например, у Reticulitermes) еще сохраняются две общие для всех особей личиночные стадии. В дальнейшем единый ствол лабильных в отношении касты особей расщепляется на два. Новой особенностью является ветвь, ведущая к развитию почти исключительно стерильных каст, т. е. рабочих и солдат. Этот путь развития состоит из пяти личиночных стадий; личинки выполняют различные работы, однако они могут превращаться в просолдат или бескрылых замещающих половых особей, но никогда не превращаются в нимф или имаго. Они явно утратили способность к развитию крыльев, но развитие гонад у них еще возможно. Другая, лабильная, ветвь состоит из шести нимфальных стадий, которые могут превращаться в замещающих половых особей или же полностью завершать цикл - до крылатых имаго; это не рабочие особи, и они не могут развиться в солдат. Однако эта ветвь лабильна в том смысле, что нимфы способны регрессировать в личинок, т. е. утрачивать свои зачаточные крылья; после этого они могут участвовать в работах, а в дальнейшем превращаться в солдат, пройдя переходную стадию просолдата. У большинства Rhinotermitidae переход к развитию по стерильной ветви зависит от времени года и от питания, а также от размеров семьи, но у представителей рода Schedorhinotermes эта ветвь для самцов закрыта, так что все работы в семье и ее защита лежат на самках.
У Termitidae бескрылых или почти бескрылых замещающих половых особей нет - они, видимо, исчезли в процессе эволюции. Очевидно, крылатые имаго могут обеспечить достаточное замещение, несмотря на то, что они появляются только в определенное время года, как у муравьев. Рабочими особями здесь также служат не личинки с затвердевшими покровами, а особи, развивающиеся из личиночных стадий в результате одной-единственной линьки - обычно окончательной, хотя в отличие от имагинальных стадий у этих рабочих сохраняются проторакальные железы. Несмотря на то что гонады у них недоразвиты, особей разного пола можно различить на личиночной стадии, и теперь можно считать установленным, что вероятность развития по стерильному пути каким-то образом связана с полом. После того как этот путь выбран, вероятность превращения в солдат тоже зависит от пола; например, у Nasutitermitinae солдатами обычно становятся самцы, а у Macrotermitinae и Termitinae - самки. У тех видов, у которых представители данной касты бывают двух разных размеров, это чаще обусловлено различием в стадии созревания, чем половыми различиями (Brian, 1979а; Noirot, 1969, 1974).
У Termitidae репродуктивная и стерильная ветви расходятся после всего лишь одной стадии, возможно даже на стадии яйца (Noirot, 1969). На пути развития половых особей имагинальной стадии предшествуют семь стадий (рис. 12.2). У такого примитивного вида, как Amitermes evuncifer, между яйцом и первой личинкой, выполняющей функции рабочего, имеются две личиночные стадии, а затем три или четыре возраста рабочих. Солдаты, развивающиеся из личинок того и другого пола, образуются большей частью из рабочих первого возраста. У Microtermes (Amitermitinae) рабочие мужского пола после двух личиночных стадий проходят еще три стадии, а рабочие женского пола - четыре стадии и в результате бывают крупнее рабочих - самцов; просолдаты тоже дивергируют рано, от рабочих первого возраста. Другие примеры можно найти у Noirot (1974) и Brian (1979а). В общем можно сказать, что у Termitidae дивергенция репродуктивной и стерильной линий происходит рано, возможно на стадии яйца; что личиночных стадий у них меньше, а нимфальных больше; что солдаты развиваются обычно из личинок определенного пола, а рабочие - одинаково из мужских и женских личинок; что особи разного пола и находящиеся на разных стадиях созревания различаются по размерам.
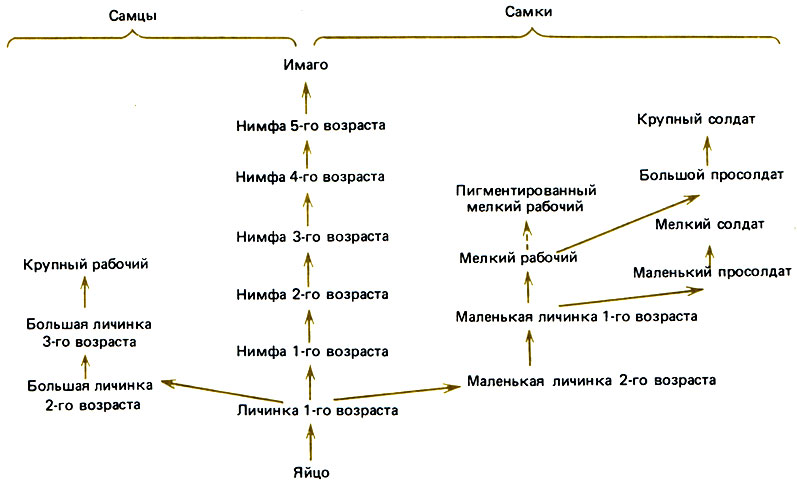
Рис. 12.2. Возможные пути онтогенетического развития у Macrotermes bellicosus. Каждая стрелка соответствует одной линьке, за исключением пунктирной, которая означает пигментацию без линьки. (Lüscher, 1976.)
Люшер (Lüscher, 1976) обнаружил у Termitidae (Macrotermes subhyalinus) материнский эффект, обусловленный различным содержанием ювенильного гормона (ЮГ) в яйце. В тот период, когда плодущая царица откладывает яйца, дающие нимф, уровень ЮГ в ее крови низок. В отличие от этого у молодых цариц уровень ЮГ в крови высокий, и все отложенные ими яйца развиваются по стерильной линии. Люшер разработал на основе этих данных общую теорию о роли ЮГ в регуляции каст у термитов; согласно этой теории, ЮГ-подобное вещество, выделяемое в пищевой поток парой половых особей, может приостановить образование замещающих особей во время чувствительной стадии цикла линек. Их способность синтезировать ЮГ, несомненно, очень велика; им нужно большое количество этого гормона, чтобы производить яйца, но остается ли его достаточно для циркуляции в семье (рис. 12.3)? Солдаты, по мнению Люшера, выделяют какое-то анти-ЮГ-вещество, прекращающее дальнейшее образование солдат; убедительно показано, что добавление аналогов ЮГ к культурам термитов, содержащим личинок в чувствительной стадии (первые 6 дней третьей личиночной стадии у М. michaelseni, согласно Okot-Kotber, 1980), повышает формирование просолдат (Brian, 1979а; Nijhout, Wheeler, 1982). Можно думать, что ЮГ делает возможным дальнейший аллометрический рост, ведущий к характерной гипертрофии головы. Высказано предположение (Nijhout, Wheeler, 1982), что сроки выбросов ЮГ в кровоток важнее, чем общее его содержание в крови.
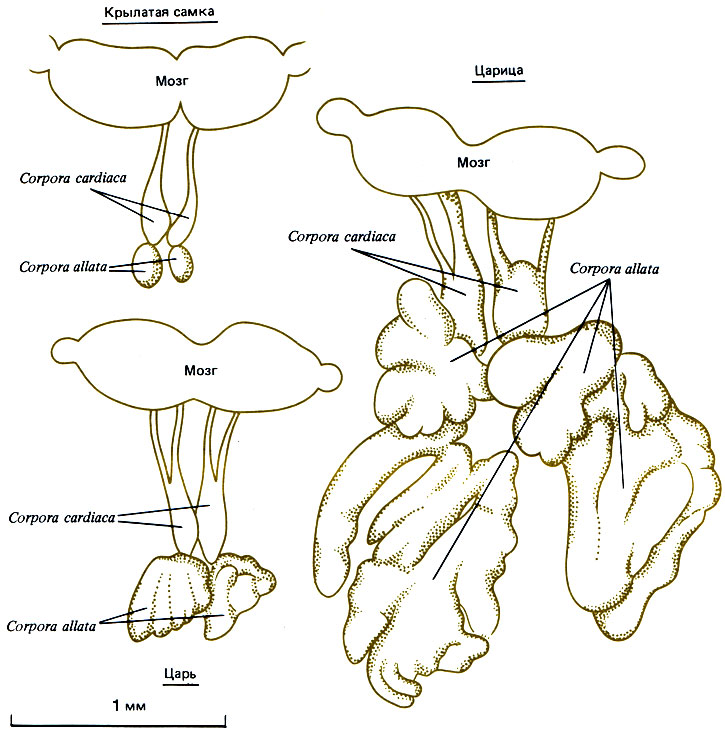
Рис. 12.3. Эндокринные системы взрослых особей Macrotermes subhyalinus. (Lüscher, 1976.)
Данных о морфогенезе каст у примитивных муравьев пока еще мало. У Odontomachus (Ponerinae) количество пищи может до очень поздних стадий влиять на кастовую дифференциацию; здесь существует один главный лабильный путь развития, но мы не знаем, не отходит ли от него ветвь на более ранней стадии или даже не бывает ли определенного кастового "уклона", заложенного матерью в яйцо. У Myrmica действуют те же регулирующие факторы, что и у Ponerinae, и, кроме того, существует несколько способов, при помощи которых самки могут управлять этим процессом. Одна, а может быть, и две линьки заменены пластификацией кутикулы, и в регулировании морфогенеза каст участвуют железы. Можно выделить два дискретных и взаимоисключающих физиологических состояния, различающихся по соотношению скоростей роста "зародыша" имаго и роста личинки в целом (под "зародышем" здесь имеется в виду совокупность имагинальных дисков, имеющихся в личинке). При формировании репродуктивной самки, соответствующем обычному процессу развития насекомого, "зародыш" растет медленно и большая часть пищи, поглощаемой личинкой, откладывается в жировом теле. Это продолжается до тех пор, пока головной мозг личинки не переместится в грудной отдел, где вокруг него формируется голова взрослой особи. После этого "зародыш" растет изометрически, у него развиваются крылья, разделенные яичники, сегментированные усики и ноги; начинается видимая дифференцировка всех частей организма, необходимых взрослой репродуктивной самке. Эти процессы становятся возможными в период прохладной погоды (обычно зимой), когда возобновляется активность corpora allata. Развитие "зародыша" метастабильно и может быть нарушено голоданием при температуре выше 23°С. "Зародыш" увеличивается в размерах и изменяет форму: увеличиваются почки ног, но не почки крыльев и не зачатки яичников, что нетрудно увидеть сквозь кутикулу личинки. В целом это приводит к увеличению относительной массы тканей "зародыша" в личинке. Происходящие изменения необратимы и сопровождаются утратой морфогенетических потенций: детерминируется развитие рабочей особи. Быстрый переход "зародыша" из одной формы в другую - это переход между двумя стабильными состояниями, и промежуточных форм в естественных условиях не образуется, если не считать тех случаев, когда под влиянием паразитов нарушается питание. В процессе роста "зародыша" рабочей особи крылья остаются небольшими, а во время метаморфоза вообще исчезают, включаясь в наружный скелет; маленький гермарий не распадается на трубочки, как у репродуктивной самки, а соединяется с яйцеводом, образуя яичник с одной яйцевой трубочкой (рис. 12.4).
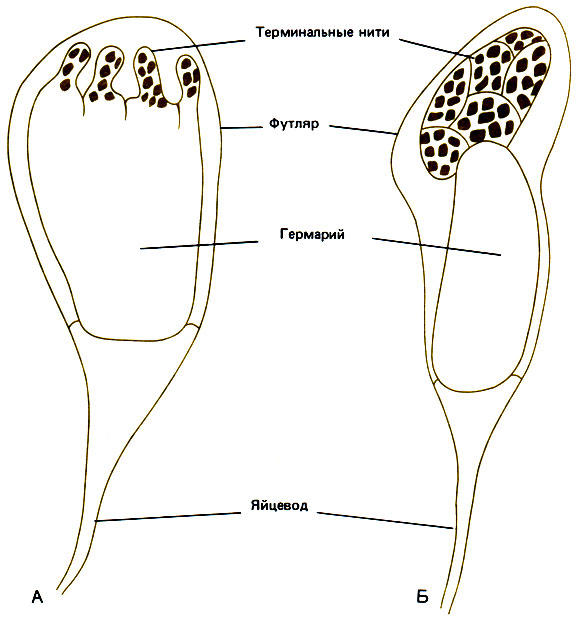
Рис. 12.4. Яичники личинок Myrmica последнего возраста, развивающихся в репродуктивных самок (А) и в рабочих особей (Б)
![Фото 30. Касты Neotermes jouteli. А. Крылатые репродуктивные формы, рабочие особи и яйца. [J. В. Free.]](pic/000106.jpg)
Фото 30. Касты Neotermes jouteli. А. Крылатые репродуктивные формы, рабочие особи и яйца. [J. В. Free.]
![Фото 30. Касты Neotermes jouteli. Б. Крупный солдат с сильно развитыми челюстями и челюстными мышцами. [J. В. Free.]](pic/000107.jpg)
Фото 30. Касты Neotermes jouteli. Б. Крупный солдат с сильно развитыми челюстями и челюстными мышцами. [J. В. Free.]
А чем объясняется меньшая величина рабочих особей? Освободившись от крыльев и яичников и увеличившись в размерах, "зародыш" продолжает свое развитие, и питание личинки прекращается на 2-3 дня раньше, чем это бывает при развитии репродуктивной самки; в результате рабочие особи оказываются мельче. Детерминация развития в направлении рабочей особи может быть сравнительно поздним отклонением от лабильной линии, ведущей к развитию репродуктивной самки (рис. 12.5); оно может произойти уже после того, как личинка накопила питательные вещества, диапаузировала, перезимовала и вновь обрела способность к росту; таким образом, здесь проявляется известное сходство с нимфой Schedorhinotermes, которая регрессирует в рабочую особь, хотя, конечно, при этом не бывает линьки и все описанные изменения происходят внутри личиночной кутикулы. Между тем многие рабочие особи (возможно, большинство их) образуются из "зародышей", дивергировавших от лабильного ствола задолго до фазы накопления запасов перед диапаузой; в их число входят все рабочие особи, происходящие из яиц, детерминированных к развитию рабочих. После такой детерминации дифференцировка обычно замедляется, личинки перестают кормиться и внезапно метаморфизируют; дифференцировка "зародыша" и его превращение в куколку внутри личиночных покровов после прекращения питания - вполне определенный процесс, и он может занимать при развитии рабочей особи столько же времени, что и при развитии репродуктивной самки. Тем не менее детерминацию к развитию рабочей особи можно рассматривать как шаг к метаморфозу, поскольку она вызывает относительный рост "зародыша" внутри личинки, сокращает время роста и тем самым уменьшает окончательные размеры рабочей особи. Это безусловно преждевременный, но незавершенный метаморфоз, необратимо искажающий развитие "зародыша" (Brian, 1974а); его формирование, по-видимому, запускается каким-то сигналом, исходящим от головного мозга, который подавляет активность corpora allata, не включая функцию проторакальных желез.
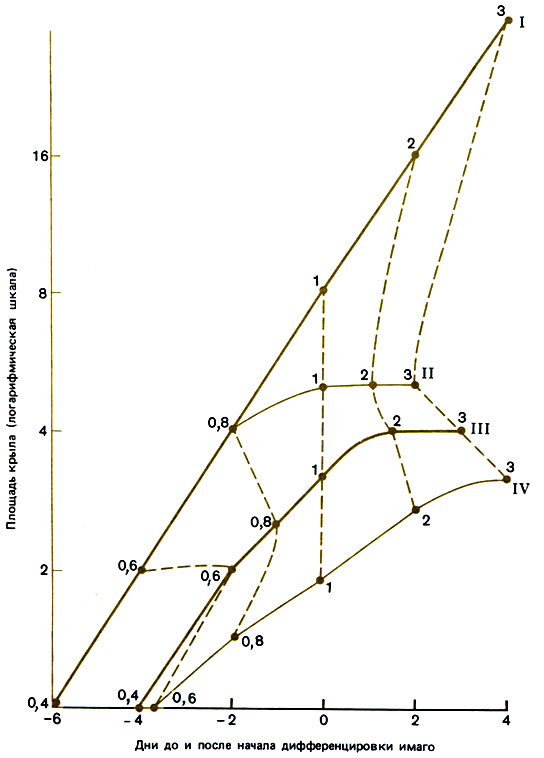
Рис. 12.5. Рост крыловых имагинальных дисков у личинок Myrmiса, I - репродуктивные самки; II - крупные рабочие особи; III - средние рабочие особи; IV - мелкие рабочие особи. Стадии дифференцировки взрослых особей обозначены цифрами 0,6; 0,8; 1; 2; 3. (Brian, 1974а.)
У Myrmica, как и у всех муравьев, только самки делятся на два типа, различающиеся по размерам, морфологии, поведению и долговечности. У самцов такого деления нет, и мужские "зародыши" не подвержены такому нарушению развития, как женские; зачатки всех частей имаго морфогенетически связаны между собой, и рост "зародыша" протекает изометрически. Роль ювенильного гормона у обоих полов состоит в том, чтобы ограничивать рост "зародыша" и способствовать запасанию питательных веществ в жировом теле личинки. Перевязка corpora allata у женской личинки, лабильной в отношении касты, ведет к детерминации развития рабочей особи, и ее можно приостановить только с помощью аналогов ювенильного гормона (Brian, 1974b). У Plagiolepis pygmaea активность corpora allata зависит не от числа, а от величины их клеток (Suzzoni, Grimal, 1980). Итак, у муравьев рабочие особи образуются из "зародыша", который заключен в червеобразной личинке, но созревает и отделяется от нее, еще не достигнув полного развития. Этот процесс обособления от личинки прекращает ее дальнейшее развитие, точно так же как личинки высших термитов навсегда становятся рабочими. В этом смысле рабочие особи муравьев и термитов гомологичны. Различия между ними обусловлены лишь тем, что у термитов ювенильные особи соприкасаются с окружающей средой с самого начала, тогда как у муравьев они вначале заключены во временный "футляр", совершенно отличный от них, освобождение из которого кладет конец пластичности их формы.
Процесс формирования солдат у муравьев все еще остается неясным. У Pheidole линия репродуктивных самок отделена от бесполой линии уже на стадии яйца, на развитие которого влияет (см. выше) концентрация ювенильного гормона в гемолимфе матери (Passera, Suzzoni, 1979; Wheeler, Nijhout, 1981). Личинки, дающие самок, толстые и лишены волосков, а личинки бесполых особей вытянутые и покрыты волосками. Отклонение развития в сторону солдат происходит на последней (третьей) личиночной стадии, на которой кишечник имеет золотистую окраску; личинок солдат можно отличить по их более крупным размерам, а также по тому, что у них имеются почки передней пары крыльев, но нет видимых почек задней пары; у личинок рабочих особей совсем нет крыловых почек (или, в редких случаях, они очень малы). У всех этих каст крыловые почки невелики по сравнению с почками ног, но у куколок, дающих солдат, на месте переднего крыла можно видеть палочковидный выступ (как у "крупных рабочих" и интеркастов Myrmica, рис. 12.6).
Рис. 12.6. А-В. Интеркасты Myrmica с палочковидными крыловыми выступами; эти особи развиваются из лабильных личинок, которые не смогли дорасти до настоящих репродуктивных самок и близки к касте, известной как солдаты (например, у Pheidole). Г. Солдат Zacryptoceros со сходными палочковидными крыльями. (Wheeler, 1910.)
Размеры солдат и рабочих не перекрываются. Путем местного введения ювенильного гормона во время чувствительной фазы на последней личиночной стадии можно получить гигантского солдата; такое воздействие приостанавливает начало метаморфоза и создает возможности для дополнительного роста, который носит аллометрический характер и приводит к характерной морфологии солдат. Оно приводит также к увеличению размеров рабочих особей, но лишь незначительному, и создает ситуацию, подобную естественному различию между двумя видами Solenopsis - S. invicta (эквивалент необработанных Pheidole) и S. geminata (эквивалент Pheidole после обработки ювенильным гормоном).
В естественных условиях солдаты формируются только в присутствии самки, при обилии пищи, высокой температуре (26°С и выше) и большом числе рабочих особей. Наличие взрослых солдат подавляет дальнейшее их образование, хотя этот эффект к концу сезона размножения уменьшается. Дело не в том, что солдаты - плохие кормилицы и не могут снабжать необходимой пищей личинок, дающих солдат; наблюдаемый эффект зависит от частоты контактов между особями и, по-видимому, связан со способностью разрушать ювенильный гормон (Edwards, 1982; Wheeler, 1982), как и у термитов. Когда удастся разработать схемы онтогенеза промежуточных типов, интересно будет выяснить, каким образом произошел переход от онтогенеза, свойственного Myrmica, с общим лабильным стволом и двумя главными путями развития (дающими мелких и крупных рабочих), к такой картине, как у Pheidole, у которого пути развития репродуктивных самок и рабочих разделены со стадии яйца, а от линии рабочих ответвляется еще линия сверхкрупных солдат (рис. 12.7) (Brian, 1979а).
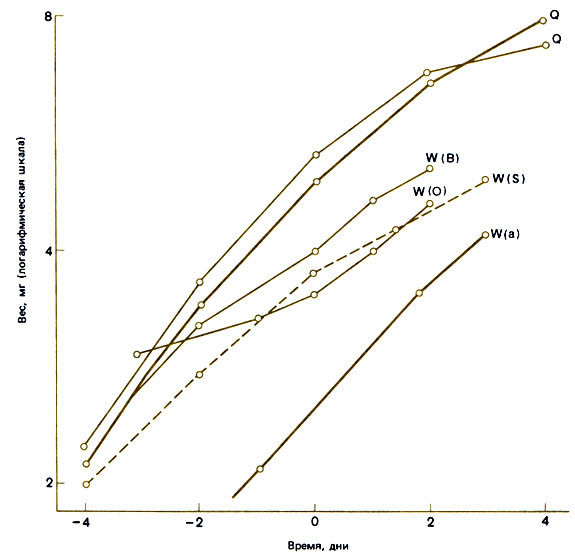
Рис. 12.7. Зависимость веса растущих личинок Myrmica от времени после начала сегментации ног. Q - самки; W (а) - нормальные рабочие особи средних размеров; W (О) и W (В) - лабильные личинки, которых выращивали в культуре перед началом зимы, когда ростовой потенциал низок; W (S) - лабильные личинки, которых выращивали в культуре после зимнего восстановления ростового потенциала, но при этом сразу после периода голодания при высокой температуре (25°С). W (О, В и S) - прототипы 'солдат' (т. е. крупные рабочие особи). (Brian, 1974а.)
12.1.2. Осы и пчелы
У ос и пчел обе касты имеют крылья, и возможно, что именно необходимость крыльев для рабочих особей была причиной, задержавшей эволюцию сильно дифференцированной касты рабочих. Маточники ос снабжаются более обильной и, возможно, лучшей по качеству пищей, содержащей больше белка, вырабатываемого слюнными железами (Montagner, 1966). Яйца не предрасположены к развитию особей той или другой касты, и большие различия в размерах между личинками появляются только после прохождения ими трех из пяти стадий. Личинки, находившиеся в маточниках, если перенести их в маленькие ячейки, регрессируют, давая рабочих особей, т. е. они лабильны в отношении касты (Spradbery, 1978а), но личинки из маленьких ячеек при переносе в маточники дают интеркастов. Они не детерминированы к развитию в рабочих особей, и, быть может, в обычных условиях каста определяется уже после того, как личинка перестает принимать пищу. После этого происходит внутренняя оценка ее трофического состояния, и метаморфоз идет в направлении той или другой морфы, приводя к образованию либо крупной репродуктивной самки в состоянии репродуктивной диапаузы, либо маленькой недиапаузирующей рабочей особи, у которой функция размножения блокирована социальными факторами.
У Bombus ячейки располагаются беспорядочно, но их содержимое часто обследуется (Pendrel, Plowright, 1981). Яйца не детерминированы. Некоторые личинки утрачивают лабильность в возрасте 3-4 дней, к концу первой стадии, что примерно соответствует четверти личиночной жизни, и их развитие направляется в сторону рабочих особей. Рабочие умеют распознавать личинок, обладающих потенциями к развитию в репродуктивных самок, и дают им больше корма, так что они проводят три лишних дня в личиночном состоянии и четыре лишних дня в виде куколок. В группах, где нет матки, число личинок, развивающихся в самок, регулируется отношением рабочие/личинки, но в присутствии матки это отношение может измениться на обратное. Концентрация ювенильного гормона у личинок последнего возраста, дающих половых самок, выше, чем у личинок рабочих особей, поэтому возможно, что задержку метаморфоза обусловливают corpora allata, как и у Myrmica. Röseler (1977) высказал предположение, что матки инактивируют corpora allata личинок, но поскольку им для этого не нужно непосредственно прикасаться к личинкам, то, вероятно, инактивирующий фактор передается через рабочих особей. Быть может, матка подает какой-то сигнал, на который рабочие особи реагируют определенным поведением, как у Myrmica.
У Meliponini кастовые различия тоже зависят от количества корма: репродуктивные самки развиваются в больших ячейках и могут при этом получать тот же корм, что и все остальные личинки, если количество этого корма будет достаточным (Darchen, Delage-Darchen, 1974, 1975). Даже у Melipona количество корма может иметь очень важное значение, реализуя или усиливая латентные генетические различия, связанные с гетерозиготностью по двум аллелям. По-видимому, детерминация осуществляется на довольно поздних этапах личиночного развития, перед самым метаморфозом, так как обработка ювенильным гормоном в этот период приводит к увеличению числа самок и уменьшению числа рабочих особей. Таким образом, внутренняя оценка кастовых потенций происходит после прекращения кормления, сопровождаясь гормональными изменениями, необходимыми для соответствующего морфогенеза.
У медоносной пчелы различия между кастами самок выражены гораздо сильнее, чем у всех других пчел. Хотя у рабочих особей сохраняются крылья, у них редуцируется половая система и развиваются специфические системы желез и особые формы поведения. Яйца не детерминированы, и кастовые различия развиваются постепенно, под влиянием различных режимов кормления. Личиночная жизнь матки продолжается всего 5 дней, а рабочей пчелы - 6 дней. Если личинку в возрасте до трех дней перенести из обычной ячейки в маточник, то из нее развивается матка, но если сделать это позднее, то разовьется интеркаст с меньшим числом яйцевых трубочек и небольшой сперматекой. Такие особи часто гибнут сами по себе или же изгоняются рабочими, которые распознают их неполноценность.
Личинки, дающие рабочих особей, в первые три дня отнюдь не голодают; они получают богатую белком пищу, синтезируемую в гипофарингеальных железах молодых рабочих пчел (молочко), и растут так же или даже быстрее, чем личинки такой же величины, дающие маток такого же размера (рис. 12.8). Однако молочко (см. разд. 9.3) не обеспечивает личинкам возможность метаморфизировать, и в течение последних трех дней им дают смесь молочка и меда. В результате у медоносной пчелы, в отличие от Bombus, выкармливание личинок рабочих особей и их метаморфоз занимают больше времени, чем выращивание личинок репродуктивных самок. Личинки, находящиеся в маточниках, получают все время один и тот же корм - маточное молочко; этот хорошо сбалансированный корм содержит 12% сахара вместо 4% в обычном молочке и богат витаминами (хотя они, по-видимому, не имеют решающего значения). Эти и другие качественные различия в снабжении кормом (здесь были перечислены лишь некоторые из них) играют важную роль, поскольку диплоидные личинки, выращиваемые в трутневых ячейках и получающие большое количество обычного молочка, превращаются в крупных рабочих особей, а не в маток. Маточное молочко при хранении теряет свои свойства, и из личинок, которые его получают, развиваются рабочие пчелы; его порча, возможно, обусловлена кристаллизацией гексоз, потому что, если добавить к обычному молочку гексозы, оно становится почти таким же хорошим кормом, как маточное молочко. Если же добавить еще и один из аналогов ювенильного гормона, оно не будет уступать ему ни в чем. Личинок, находящихся в маточниках, кормят секретом не только гипофарингеальных, но и мандибулярных желез, и притом в 10 раз чаще, чем личинок в рабочих ячейках, и они, несомненно, все время получают хорошо сбалансированный корм. Это имеет важнейшее значение (Beetsma, 1979; Brian, 1979а, 1980).
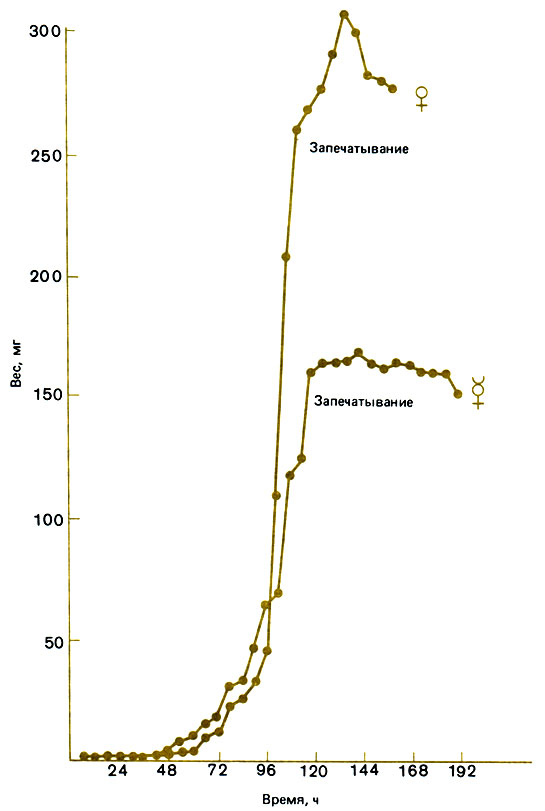
Рис. 12.8. Рост личинок самок и рабочих особей у медоносной пчелы. (Wirtz, 1973.)
Здесь можно провести интересное сравнение с Myrmica. У личинок рабочих особей в возрасте 1-3 дней не откладывается в жировом теле гликоген, тогда как у личинок, развивающихся в маточниках, это происходит. Подобное же различие можно обнаружить у личинок последнего возраста Myrmica; оно означает, что для морфогенеза самок характерны высокий уровень запасания питательных веществ и медленный рост "зародыша" имаго, тогда как при формировании рабочих особей вся потребляемая пища сразу же идет на развитие физиологически доминантного и активного "зародыша". Это, как и у Myrmica, вероятно, обусловлено высокой концентрацией ювенильного гормона у личинок, из которых развиваются самки, что весьма интересно, так как было показано, что у развивающихся в репродуктивную самку личинок Apis в возрасте трех дней концентрация этого гормона в крови в 10 раз выше, чем у личинок рабочих особей (Beetsma, 1979). Как показали эксперименты, ювенильный гормон вызывает "включение" сразу всего комплекса особенностей репродуктивных самок. Прекоцен - фактор, разрушающий corpora allata, но не снижающий потребления корма, может иногда препятствовать развитию репродуктивных самок. Таким образом, синдром запасания корма и формирования репродуктивных самок, вызываемый высоким уровнем ювенильного гормона в крови, наблюдается и у муравьев, и у пчел.
Эти различия в концентрации ювенильного гормона продолжают влиять на метаморфоз, когда на личиночной стадии впервые появляются кастовые отличия будущей матки медоносной пчелы, у которой яичники необычайно велики и содержат около 300 яйцевых трубочек; все эти трубочки сохраняются, а у рабочих особей число их, превышающее к концу личиночной стадии сотню, сокращается примерно до 12. Итак, самка, потенциально (генетически) способная иметь 300 яйцевых трубочек, при образовании рабочих особей вынуждена эпигенетическим путем утратить из них 288. Материал, освобождающийся в результате их дегенерации, во время метаморфоза, несомненно, включается в восковые железы, в приспособления для сбора пыльцы и в другие специальные структуры рабочих. Стабилизация яичников у личинок рабочих особей может быть достигнута за 1-2 дня в маточнике, где потребление маточного молочка ведет к увеличению corpora allata и повышает уровень ювенильного гормона в крови примерно в 10 раз. Следует отметить еще один интересный момент: даже в стеклянных ячейках в лаборатории личинки, дающие репродуктивных самок, после того как они сплели кокон, ориентированы вертикально головой вниз, тогда как личинки рабочих особей лежат горизонтально. Такое поведение первых соответствует поведению всех Apidae, и его можно индуцировать и у вторых с помощью ювенильного гормона (Beetsma, 1979). Личинки рабочих особей, очевидно, адаптированы к жизни в вертикальных сотах с почти горизонтальными ячейками; некоторые гены, определяющие терминальные этапы развития, никогда не вступают у них в действие.
Из всех данных, собранных многими учеными за много лет (здесь упомянута лишь небольшая их часть), становится ясно, что различные пищевые режимы приводят к различиям в развитии желез, влияющих на план и процесс перестройки и реорганизации развития на стадии куколки. Пока не известно, каким образом организм оценивает различия в корме. Оказывает ли пища влияние на нейроэндокринную систему после того, как она всосалась из кишечника, т. е. через трофику, или же дело в том, что пища содержит какие-то вкусовые вещества, стимулирующие рецепторы ротовой области, и воздействует на нейроэндокринную систему через них? Иными словами, трофическое это воздействие или сенсорное?
Beetsma (1979) и его сотрудники придерживаются второй точки зрения; они обнаружили в лабиальной области личинки рецепторы, чувствительные к сахарам. По их мнению, содержание сахара в молочке в первые три дня не настолько высоко, чтобы увеличить потребление пищи. Тем не менее в эти дни личинки в рабочих ячейках растут так же быстро, как личинки в маточниках, а значит, поедают столько же пищи; лишь позднее, когда молочко разбавляется медом, скорость роста падает. О влиянии питания личинок на эпигенетическую регуляцию морфогенеза известно слишком мало; главную роль сейчас приписывают corpora allata, однако известно, что в регуляции участвуют и проторакальные железы, причем не только сами по себе, но и во взаимодействии со всей системой эндокринных желез (Brian, 1980).
Кастовые различия у всех общественных насекомых обусловлены различиями в питании, направляющими развитие по разным генетически детерминированным каналам, т. е. в конечном счете различиями в поведении рабочих особей. У эусоциальных видов это поведение зависит от матки. У высших муравьев и термитов, но не у пчел и ос этот фактор сочетается с глубоким влиянием, исходящим от внутреннего состояния матери во время формирования яиц. В совокупности все эти факторы дают возможность "зародышу имаго", живущему и развивающемуся в футляре, которым служит личинка, доминировать в смысле поглощения пищи. "Зародыш", конечно, зависит от личиночных механизмов получения и переваривания пищи, однако по мере своего роста он затрудняет кормление личинки, что в конечном итоге лишает пищи его самого, и в результате он никогда не достигает таких размеров, как "зародыши" репродуктивных самок. Возможно, что в организме матки фактором, влияющим на детерминирование касты, служит ювенильный гормон - так же, как в личинке и куколке.
Судя по имеющимся данным, гены вряд ли оказывают какое-либо действие помимо создания ограниченного полом диморфизма у самок, отсутствующего у самцов (Hymenoptera). У одного муравья (Harpagoxenus) и у одной пчелы (Melipona) было показано участие генов в определении каст. Известно также несколько случаев различий между кастами муравьев по изозимам: при скрещивании Solenopsis geminata с S. xyloni все рабочие обладают гибридными свойствами, но 95% самок относятся к материнскому типу. Это означает, что отцовские гены подавляются только тогда, когда гибриды развиваются в репродуктивных самок (Hung, Vinson, 1977). У видов Myrmecia рабочие особи тоже отличаются от самок по генным частотам, но во время написания обзора Крозье (Crozier, 1980) это был единственный известный случай. Полиморфизм имеет генетическую основу, но морфогенез касты зависит от материнской гемолимфы и от режима питания личинок.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'