2.3. Сахара как источник энергии, уменьшающий потребность в животной пище
Многие осы охотятся на животных только тогда, когда у них есть личинки, поскольку трупы животных трудно запасать впрок. Виды рода Vespula охотятся, летая около листвы и цветков и схватывая самых разнообразных мелких мягкотелых насекомых при помощи одних только челюстей; осы посещают также раскрывшиеся цветки, из которых собирают сахара (Proctor, Yeo, 1973; Spradbery, 1973а).
Крупные австралийские муравьи-бульдоги (Myrmecia) охотятся даже на таких насекомых, как пчелы, убивают их с помощью жала, разрывают на части и кладут около своих крупных личинок, которые ими кормятся. После того как личинки окуклились, взрослые особи начинают кормиться сами, высасывая сахаристые выделения растений. В подсемействе Myrmicinae у примитивных форм жало редуцировано, а у эволюционно продвинутых -видоизменено. Даже у примитивных родов оно неспособно достаточно эффективно парализовать жертву, поэтому для того, чтобы схватить ее, раздавить, разорвать или проткнуть дырки в ее кутикуле, обычно используются челюсти; затем хищник высасывает из жертвы соки. Эти муравьи собирают также соки плодов и нектар, извлекая энергию из содержащихся в них Сахаров; тем самым они экономят белки, большую часть которых получают из мертвых беспозвоночных, а не в результате охоты. В лабораторных условиях Myrmica вполне довольствуются рационом, состоящим из насекомых и воды, но если добавить к этому раствор сахарозы, число метаморфизирующих личинок и откладываемых яиц возрастает. Муравьи при этом даже поедают меньше насекомых и выделяют прозрачную мочу вместо беловатой с зернышками мочевой кислоты. Очевидно, сахара стимулируют активность рабочих особей и используются ими в качестве источника энергии, тогда как белки расходуются на образование яиц и выкармливание личинок (Brian, 1973а).

Фото 5. Мандибулы рабочей особи Myrmica. Это молодая особь с острыми кутикулярными зубцами, вцепившимися в частицу пищи (х70)
![Фото 6. Мандибулы старой рабочей особи Myrmica. Зубцы изношены, а один из них полностью обломан (х 70). [Фото 5 и 6 - С. Hawkins, Univ. of Southampton.]](pic/000015.jpg)
Фото 6. Мандибулы старой рабочей особи Myrmica. Зубцы изношены, а один из них полностью обломан (х 70). [Фото 5 и 6 - С. Hawkins, Univ. of Southampton.]
Другим обычным источником Сахаров служит падь, которую выделяют различные насекомые, сосущие соки растений; это главным образом представители Aphidae, Coccidae и Aleurodidae. Три вида Camponotus (Formicinae), живущие на деревьях в Восточной Африке, активно используют для этого алейродид, а также щитовок и тлей, реже - насекомых, относящихся к трем другим группам (Lévieux, 1977). Эти муравьи собирают камеди, выделяемые растениями, и охотятся на личинок из семи и взрослых особей из девяти отрядов насекомых. Другая группа, Membracidae, питающаяся в восточной части Северной Америки на золотарнике, привлекает муравьев нескольких разных видов. Самый крупный из них - один из видов Formica - уничтожает листоядных насекомых, и если он присутствует в данной местности большую часть года, то это может способствовать росту золотарника и образованию его семян (Messina, 1981).
Тли выделяют через анальное отверстие жидкий секрет, попадающий обычно на листья, которые становятся липкими и покрываются плесенью, если муравьи, осы и пчелы не соберут этот секрет. Он содержит не только растительный сок (за вычетом того, что использовали для себя тли), но также выделения самих тлей (Way, 1963). Интересные данные были получены недавно в опытах с тлей Myzus persicae, которую культивировали на редьке (Raphanus sativus), выращиваемой в цветочных горшках. Эти растения содержат глюкозу и фруктозу, а падь содержала, кроме того, трегалозу, мелецитозу и сахарозу; растения содержат семь карбоновых кислот и пять кислот - производных фенола, а в пади их содержалось 8 и 18 соответственно. Другие соединения - растительные гормоны ауксин и гиббереллин, а также цитокинины и ингибиторы роста -попадают в падь из растения, но аммиак добавляют в нее тли, что ведет к повышению рН (Hussain et al., 1974). Падь содержит от 9 до 23 аминокислот и амидов, составляющих 2-3% ее сухого вещества; как и следовало ожидать, состав пади варьирует в зависимости от вида как насекомого, так и растения, а также от времени года (Auclair, 1963). Падь содержит также небольшие количества многих других веществ, полученных от растения или синтезированных тлей.
В умеренных областях осы и пчелы с короткими хоботками регулярно потребляют падь. Залетая на верхушки деревьев в тех районах, где муравьев мало, они могут без всякой конкуренции собирать ее в больших количествах; это особенно ценно для них в периоды, когда подходящих распустившихся цветков мало или они недостаточно активно выделяют нектар вследствие засухи. Из выделений растительноядных сосущих насекомых получается очень хороший мед.
Случайный сбор пади с листьев в процессе эволюции перерос в прямую связь донор - реципиент. В настоящее время существуют тли, которые не выделяют свой жидкий секрет до тех пор, пока к ним не прикоснется муравей, а после этого они сразу отдают его в виде капельки, которая держится на волосках, окружающих анальное отверстие. При этом тли подавляют свои обычные защитные реакции, такие как выбрасывание клейкого вещества из соковых трубочек. Поскольку муравьи отпугивают хищников и паразитов и часто строят укрытия от ветра и дождя, кооперирующиеся с ними насекомые имеют больше шансов выжить и оставить потомство, чем другие. Даже если их связь с муравьями носит еще факультативный характер, популяции этих насекомых получают от муравьев заметную выгоду; Уэй (Way, 1963) в своем обзоре отмечает, что тли, помимо защиты, могут получать, возможно, еще целый ряд прямых и косвенных преимуществ: они поглощают больше пищи и выделяют больше пади, образуют более многочисленные скопления, и плотность их популяций повышается. Это оказывает успокаивающее действие, в результате которого число бродячих особей снижается и каким-то еще не вполне понятным образом появление крылатых половых форм происходит позднее, чем это было бы в обычных условиях при данной плотности популяции. Быть может, муравьи производят выбраковку среди молодого потомства тлей?
Лесные муравьи (Formica rufa и близкие виды) собирают огромные количества пади с тлей, которые кормятся как на лиственных, так и на хвойных деревьях в лесах северной умеренной зоны. Большая полигинная семья F. lugubris, живущая в горах Юры (Швейцария/Франция), охотится на насекомых, почти равных по величине самим муравьям; среди их жертв много сосущих насекомых. По мнению Cherix (1980), муравьи уничтожают так много насекомых, что насекомоядные птицы не могут жить в этих местах. Многие их жертвы относятся к растительноядным сосущим насекомым, но, кроме того, начиная с июня лесные муравьи собирают также падь у нескольких видов тлей, особенно у Cinara pruinosa, которая кормится на пихте (Picea abies). Позднее Cinara тоже становится жертвой муравьев; Cherix (1980) описывает свои наблюдения над рабочими особями муравьев, напавшими на тлей, которые покинули свою группу и производили необычные движения; были ли они заражены паразитами или же просто одряхлели? Ясно, что граница между защитой и хищничеством весьма зыбкая.
Биология питания лесных муравьев усиленно изучается ввиду потенциальной возможности использовать их для биологической борьбы с гусеницами, поедающими листву; натуралисты указывают на зеленые островки деревьев с пышной листвой вблизи гнезд лесных муравьев. Formica rufa у северной границы своего ареала на Британских островах вылавливают весной тлей Drepanosiphum platanoides, причем пик уничтожения тлей совпадает с пиком их численности. Муравей никогда не заботится об этих тлях. К числу его жертв относятся также многие двукрылые, в том числе толстоножка (Bibio), и гусеницы, главным образом гусеницы зимней пяденицы (Operophtera brummata), которых он собирает с дубовых деревьев (Quercus), особенно в мае (Skinner, 1980b). Падь этот муравей берет от другой тли - Periphyllus testudinaceous - на платанах (Pseudoplatanus sp.); ее сбором занимается более 60% сборщиков, и она обеспечивает больше половины всего энергетического материала. Этот муравей хватает всех мелких насекомых, обитающих на земле и на деревьях, если их много и их нетрудно ловить. Интересным примером служит объедающая листву гусеница зеленой дубовой листовертки (Tortrix viridana): хотя, по данным Иноземцева (1974), муравьи сократили численность популяции листовертки на одну треть, Скиннер сообщает, что они уничтожили меньше особей, чем можно было ожидать, но указывает на то, что учет мог быть неточным, поскольку муравьи, возможно, разрывают гусеницу на части, а затем заглатывают ее прямо на месте. Если с помощью липких лент преградить муравьям доступ к Drepanosiphum platanoides, то численность тлей возрастает (Skinner, Whittaker, 1981). Муравьи снижают также численность популяции гусеницы, особенно на нижних ветках; по имеющимся оценкам, на тех деревьях, где охотятся муравьи, площадь листвы, съеденной гусеницами, составляет лишь 1,2%, а на деревьях, где муравьев нет, - 8,5%. Таким образом, Formica rufa уменьшает вред, наносимый гусеницами листьям, и несколько снижает численность популяций некоторых тлей, но общая численность тлей - главных продуцентов пади - возрастает и количество производимой пади довольно значительно (3,5·107 Дж на одно дерево). Соотношение ущерба и выгоды для дерева вычислить очень трудно (см. также Wellenstein, 1980).
Хороший пример сложности взаимоотношений в системах "муравей - тля-дефолиатор - дерево" рассмотрен в работе (Laine, Niemela, 1980), проведенной в северной Финляндии (70° с. ш.) на Formica aquilonia (лесном муравье, обитающем на севере Европы), березе пушистой (Betula pubescens) и поедающей ее листву гусенице Oporima autumnata. Этот муравей стабилизирует плотность своей популяции в лесах на высоте ниже 240 м на уровне примерно 5 гнезд на гектар, используя кормящуюся на березе тлю, выделяющую падь. Он способен при этом нападать на гусениц Oporima, когда они появляются, и препятствовать нанесению большого вреда березам. Однако муравьи действуют наиболее активно вблизи своих гнезд; например, в один обычный год повреждение листьев упало с 67 до 43% в 40 м от гнезда, а в год "взрыва" численности гусениц муравьи смогли предотвратить серьезную дефолиацию лишь вокруг своего муравейника, где сохранился "зелёный остров". Таким образом, березы, которые дают приют тлям, снабжающим муравьев пищей, и сбрасывают веточки, пригодные для постройки муравьиного гнезда, имеют больше шансов на процветание, чем те, которые этого не делают; но если они разрастутся чересчур пышно, то они могут затенить муравейник, что заставит муравьев уйти, - значит, вся эта система находится в состоянии очень тонкого метастабильного равновесия (Wielgolaski, 1975).
Некоторые лесные муравьи охотятся на одних тлей, а падь берут у других, а некоторые используют один и тот же вид тлей в обеих целях, но в разное время года. Обитающий на деревьях тропический муравей-портной Oecophylla longinoda (Formicinae) - жестокий хищник, но он выращивает несколько видов насекомых. Одно из них - щитовку Saissetia Zanzibarensis (Coccidae) - он защищает, обращается с ним мягко и собирает его падь; но как только рабочие насытятся сахаристой жидкостью, они становятся более агрессивными, убивают щитовок, поедают их сами ради белка и скармливают своим личинкам. Уэй (Way, 1954, 1963) обнаружил, что между популяциями щитовок и муравьев существует приблизительное равновесие. В Европе другой муравей, Lasius flavus, живет почти постоянно под землей, образуя облигатные ассоциации со многими видами корневых тлей. Эти тли могут обходиться без крылатых половых форм, так как муравьи не только защищают их, но и расселяют; муравьи даже собирают и прячут яйца тлей, а молодых особей переносят на листья кормовых растений. В одном сообществе, которое изучал Pontin (1978), семь видов тлей жили под землей на корнях различных злаков. Этот автор установил существование стабильной зависимости хищник - жертва, выражающейся в том, что часть молодых тлей первой возрастной стадии муравьи скармливают своим личинкам; он вычислил, что одна семья Lasius flavus, занимающая 5-7 м2 луговины, может существовать и расти благодаря одной только этой мутуалистической зависимости, хотя муравьи, несомненно, поедают и других почвенных насекомых.
При изучении сообщества муравьев в западной части Шотландии, в которое входило 6 видов, Muir (1959) обнаружил, что они связаны с 14 видами тлей, кормящихся на 21 виде травянистых растений. Большая часть этих тлей - облигатные сожители муравьев (мирмекофилы), принадлежащие к трем видам-полифагам. Например, Forda formicaria, которую выращивают муравьи пяти видов, кормится под землей на 12 видах растений (девять из них - злаки); Tetraneura ulmi, которую выращивают муравьи пяти видов, кормится под землей на 9 видах растений (все они - злаки). Все три мирмекофила предпочитают и чаще всего поедают Agrostis alba, A. tenuis и Festuca rubra. Тем не менее муравьи используют дополнительно и ряд других видов тлей, кормящихся на различных растениях над землей и под землей. Уэй (Way, 1963) приводит и много других примеров ассоциаций муравьев с сосущими насекомыми.
Пример с лесными муравьями позволяет думать, что присутствие муравьев может оказаться довольно выгодным для деревьев, что они могут даже приютить больше тлей, выделяющих падь, чем других тлей. Существует, однако, более прямой способ обеспечить это, зависящий от самого дерева: надо, чтобы нектарники, находящиеся на тех частях побега, которые нуждаются в защите, были активны только в период, когда эта защита необходима.
У североамериканской поздней черемухи (Prunus serotina) на молодых листьях есть нектарники, выделяющие нектар в течение примерно трех недель сразу после раскрытия почек, и именно в это время вылупляются питающиеся листьями гусеницы рода Malacosoma. Муравей Formica obscuripes (из группы лесных муравьев) посещает эти нектарники и, встретив там маленькую гусеницу, утаскивает ее в свое гнездо; более крупным гусеницам удается избежать этого. Tilman (1978) сумел показать, что этот муравей может так сильно понизить численность гусениц Malacosoma первых возрастов, что впоследствии больших гусениц окажется недостаточно для серьезной дефолиации. Другие примеры можно найти у Bentley (1977).
Многолетний подсолнечник Helianthella quinquenensis (Asteraceae) растет на высотах свыше 2500 м в Скалистых горах; его нектарники, расположенные по краям прицветников соцветия, выделяют секрет, не только богатый сахарозой, но и содержащий 19 аминокислот в концентрации, в 16 раз более высокой, чем в нектарниках цветков. Это привлекает к каждому соцветию до 40 различных видов муравьев, главным образом опять-таки Formica obscuripes. Они приходят из своих гнезд, находящихся в почве, и собирают нектар из прицветников, оставляя нектар цветков шмелям, опыляющим эти растения. Было показано (Inouye, Taylor, 1979), что на высотах свыше 3000 м муравьи отпугивают насекомых, пытающихся отложить яйца в цветки и тем самым воспрепятствовать завязыванию семян (рис. 2.4). Так как другие растения того же семейства используют для защиты от насекомых системные репелленты, возникает вопрос (на который пока нет ответа): какой из этих двух способов легче может выработаться в процессе эволюции, более эффективен и требует меньших затрат?
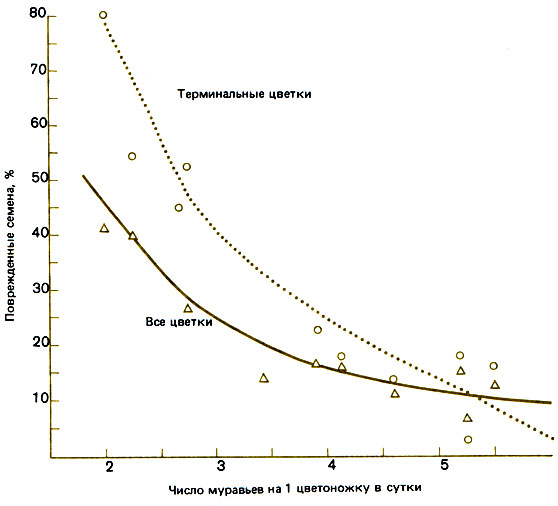
Рис. 2.4. Зависимость между числом муравьев, находящихся днем на одном соцветии Helianthella, и процентом поврежденных семян. (Inouye, Taylor, 1979.)
Тропическое растение Costus woodsonii секретирует из прицветников нектар, богатый сахарами и аминокислотами. Доминируют на нем, сменяя друг друга в зависимости от времени года, два вида муравьев: Camponotus platanus (Formicinae) и Wasmannia auropunctata (Myrmicinae) - первый в засушливый период, а второй в период дождей (рис. 2.5). Schemske (1980) показал, что они защищают плоды от мухи, которая откладывает яйца под прицветники, так что ее личинки уничтожают семена и присеменники. Camponotus прогоняет самок, откладывающих яйца, а более мелкие Wasmannia пролезают между прицветниками и поедают личинок. Семена распространяют птицы, если только маслянистые присеменники не были уничтожены (без них семена не созревают). Schemske указывает, что вид W. auropunctata - широко распространенный в тропиках вид, лишь недавно проникший в нарушенные тропические местообитания, и что вряд ли он мог играть какую-то роль в эволюции нынешней ассоциации муравей - растение. Хотя экстрафлоральным нектарникам удается избежать посягательств насекомых, высасывающих растительные соки, эта система привлечения на помощь муравьев может иметь успех лишь при достаточном числе муравьиных гнезд в непосредственной близости от деревьев.
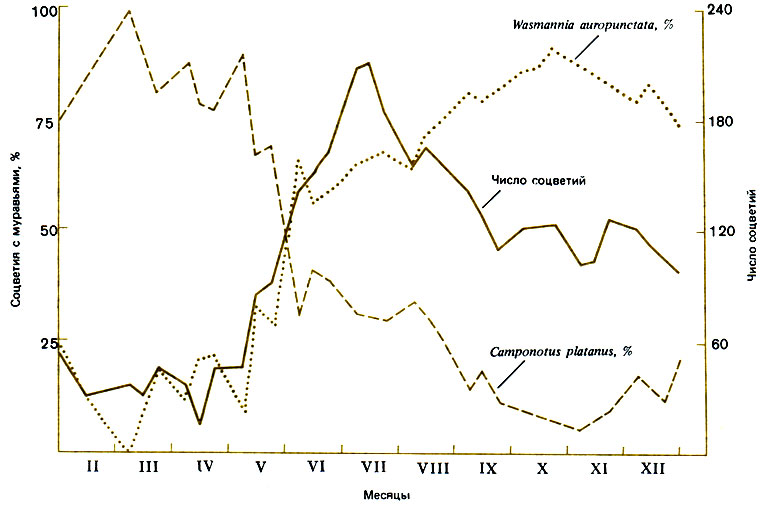
Рис. 2.5. Число соцветий на тропическом растении Costus woodsonii в разное время года и частота нахождения на них двух видов муравьев - Camponotus platanus и Wasmannia auropunctata. (Schemske, 1980.)
У некоторых тропических деревьев способы привлечения муравьев достигли своего логического завершения: у них имеются не только экстрафлоральные нектарники, содержащие сахара, но и подходящие места для устройства муравьиных гнезд и даже твердый белковый и жировой корм. Многие виды Acacia (Papilionaceae) привлекают муравьев с помощью нектарников, расположенных на стеблях, но некоторые виды предоставляют им также твердый корм, богатый белками и жирами, в виде "пищевых телец" (тельца Бельта), находящихся на кончиках листочков; они обеспечивают муравьям свыше 90% твердого корма. У этих видов Acacia стеблевые нектарники в изобилии выделяют нектар и могут продуцировать до 40 г глюкозы и фруктозы в день. Кроме того, полости в специализированных вздутых колючках служат местами для устройства гнезд, и в них живет один из видов Pseudomyrmex (Pseudomyrmecinae). У этих муравьев крепкие жала, и они кусают, жалят и убивают насекомых- фитофагов, прилетающих на растение; некоторых они затем съедают, а других просто бросают. Создается впечатление, что муравьи относятся к такому "гнездовому дереву как к своей территории и поэтому защищают его более эффективно. Они не только нападают на млекопитающих, но и очищают от травы некоторый участок вокруг дерева и уничтожают тянущиеся к нему ветви других деревьев. Эта защита от других растений предоставляет, конечно, дереву больше места и, кроме того, препятствует распространению огня (Jansen, 1966). Защитники деревьев есть и в другом подсемействе муравьев - Dolichoderinae: виды Azteca живут на деревьях рода Cecropia, которые предоставляют им пищу и жилье, а волоски, имеющиеся у основания каждого черешка, содержат помимо прочих питательных веществ гликоген (Stradling, 1978b; Wheeler, 1942).
Эти последние примеры демонстрируют высшую ступень мутуалистических взаимоотношений между муравьями и растениями: обеспечивая дереву защиту, муравьи в свою очередь получают территорию, удовлетворяющую все их потребности в пространстве и пище. В последующих разделах будут описаны менее тесные отношения, существующие между муравьями и семенами или зелеными листьями и между пчелами или осами и пыльцой.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'