Глава VIII. Хвое- и листогрызущие вредители и меры борьбы с ними
Хвое- и листогрызущие насекомые питаются тканями листьев (хвои) и в активных фазах развития ведут открытый образ жизни; только сравнительно небольшая часть их в фазе личинки живет внутри листьев, минируя их. В эту группу входят представители разных отрядов лесных насекомых. Бабочки, пилильщики и ткачи характеризуются большими калебаниями численности, и по этому признаку их обычно объединяют в отдельную группу массовых хвое- и листогрызущих, или первичных, вредителей. Остальные, в основном жуки (листоеды, слоники, нарывники и др.), не дают столь ярко выраженных вспышек массового размножения, характеризуются более умеренными колебаниями численности и образуют очаги преимущественно в молодых насаждениях, парках и полезащитных полосах.
Описание образа жизни листоедов и других жуков, повреждающих листву на деревьях, было дано в гл. VII. Поэтому дальнейшее изложение будет относиться только к группе массовых хвое- и листогрызущих насекомых.
Биологические особенности
Массовые хвое- и листогрызущие насекомые характеризуются рядом биологических черт, определяющих резкие колебания численности и возникновение вспышек массового размножения в насаждениях, где условия наиболее благоприятны для их жизни.
Благодаря открытому образу жизни насекомые в активных фазах развития подвергаются непосредственному воздействию климатических факторов, благоприятствующих их питанию, росту, развитию, расселению и размножению или вызывающих большую смертность. Кроме того, они легко уязвимы для врагов и часто в большом количестве уничтожаются птицами, хищниками, паразитами, а также погибают от болезней.
В фазе личинки (гусеницы) насекомые этой группы питаются хвоей или листвой, а в фазе взрослого насекомого используют нектар цветков или не питаются совершенно. В связи с этим все процессы жизнедеятельности у взрослого насекомого протекают в основном за счет тех резервных питательных веществ, которые были накоплены в жировом теле личинки. Накопление этих веществ у личинок (гусениц) зависит от времени их отрождения и сроков питания, в период которого листья могут иметь различный биохимический состав тканей. Особенности питания личинок обусловливают изменчивость плодовитости и выживаемости потомства. Для всех массовых хвое- и листогрызущих насекомых характерна очень высокая потенциальная плодовитость и кучность откладки яиц. Большинство хвое- и листогрызущих насекомых обладает хорошо развитыми локомоторными функциями, способно к дальнейшим и быстрым перелетам, приспособлено к пассивному переносу ветром (гусеницы монашенки, непарного шелкопряда и краснохвоста разносятся ветром) и т. п. Наконец, для ряда представителей этой группы, в частности чешуекрылых, характерна способность к концентрации расселяющейся фазы.
Преобладающее большинство хвое- и листогрызущих насекомых имеет одногодичный жизненный цикл. Только некоторые из них в более южных районах дают две генерации в год (обыкновенный сосновый пилильщик), а в районах Сибири развиваются в течение двух лет (сибирский шелкопряд).
При определенных экологических условиях большинство представителей этой группы впадает в диапаузу, которая возникает на разных фазах развития. Так, у монашенки, непарного и кольчатого шелкопряда имеется устойчивая эмбриональная диапауза, а у ряда пилильщиков в диапаузу впадают личинки во время коконирования, и диапауза длится 3 - 4 года и более. У пушистого шелкопряда диапауза на фазе куколки может продолжаться от 4 до 8 лет, а у лунки серебристой и дубовой хохлатки, как правило, перележивает 1 - 2 года 10 - 15% всех зимующих куколок.
По характеру питания хвое- и листогрызущих насекомых можно разделить на две группы.
Одна группа видов специализирована на питании растениями весной, когда листья наиболее богаты белком и, следовательно, очень питательны, хотя и имеют неустойчивый, изменяющийся химический состав. По циклу развития насекомые этой группы делятся на несколько групп:
а) зимующие в фазе яйца (непарный и кольчатый шелкопряды, монашенка, дубовая зеленая, боярышниковая и ряд других листовертов, зимняя пяденица, рыжий сосновый пилильщик и др.);
б) зимующие в фазе гусеницы (златогузка, ивовая волнянка, сосновый и сибирский шелкопряды и др.);
в) зимующие в фазе куколки (сосновая совка, пяденицы-шелкопряды, ранневесенние совки, кленовая стрельчатка и др.).
Другая группа видов специализирована на использовании растений в летний период вегетации, когда листья меньше содержат белка, не так питательны, но имеют более устойчивый химический состав. Чаще всего насекомые этой группы зимуют в фазе куколки (лунка серебристая, краснохвост, сосновая пяденица и др.).
Большинство листогрызущих насекомых многоядны, однако при питании на различных древесных породах имеют разную выживаемость, плодовитость и сроки развития. Оптимум развития у этих видов наблюдается при питании на ограниченном круге растений; в разных географических районах предпочтение отдается различным древесным породам. Так, непарный шелкопряд в лесостепи испытывает угнетение при питании листьями липы, клена остролистного и березы, а севернее Москвы березу предпочитает дубу.
Есть среди листогрызов и монофаги, но число их ограничено (зеленая дубовая листовертка, дубовая хохлатка).
Хвоегрызущие насекомые обычно предпочитают какую-либо одну породу и плохо развиваются на остальных хвойных (сосновый шелкопряд и сосновая пяденица очень неохотно питаются елью). Даже в пределах p. Pinus сосновый обыкновенный пилильщик и сосновая совка предпочитают сосну обыкновенную сосне банкса и почти не трогают сосны черную и веймутовую.
Выживаемость многих представителей хвое- и листогрызущих насекомых тесно связана с совпадением сроков развития личинок и отдельных фаз вегетации древесных пород. Так, минимальная смертность гусениц зеленой дубовой листовертки первого возраста бывает в том случае, когда массовый выход гусениц совпадает с фазой открытой почки, куда они забираются после отрождения. В связи с этим многие виды предпочитают ранние формы дуба и вяза поздним формам (зимняя пяденица, непарный шелкопряд, зеленая дубовая листовертка). Монашенка больше повреждает рано распускающуюся красношишечную форму ели, а, развиваясь на сосне, гусеницы первого возраста выживают только при питании мужскими цветками и самыми нежными иглами молодых побегов.
Отдельные виды хвое- и листогрызущих насекомых различна реагируют на окружающие гигротермические условия. Одни из них исключительно свето- и теплолюбивые, заселяют хорошо прогреваемые изреженные насаждения и опушки; предпочитают типы леса, произрастающие по повышенным элементам рельефа, на сухих и бедных почвах (златогузка, непарный шелкопряд, сосновая совка, сосновые пилильщики), другие предпочитают более влажные местообитания. Они преобладают в хорошо сомкнутых насаждениях, в глубине лесных массивов, тяготеют к более увлажненным типам леса и пойменным лесам, более гигрофильны и менее светолюбивы (зимняя пяденица, сосновая пяденица, монашенка и др.).
Вспышки массового размножения
В гл. III была изложена общая схема динамики численности насекомых. Она распространяется и на группу хвое- и листогрызущих насекомых, у которых особенно резко проявляются подъемы и падения численности. Это в значительной мере обусловлено биологическими особенностями популяций данной группы, о которых говорилось выше. Выход популяции из депрессии связан с целым рядом прямых и косвенных проявлений климатических факторов.
Вспышки массового размножения обусловлены крайними отклонениями отдельных метеорологических элементов от нормы и их аномальным развитием в течение нескольких лет. Только крайние отклонения могут вывести популяцию из нормы, изменить соотношение в развитии хозяина с комплексом энтомофагов, вызвать разрыв или усилить совпадение в фенологии кормовых растений и питающейся фазы вредителя, способствовать развитию эпизоотий и массовой гибели популяции насекомого.
Кривая изменения численности популяции для каждого вида на протяжении одного периода массового размножения специфична. Однако в практических целях можно считать, что виды хвое- и листогрызущих насекомых с одногодичной генерацией при нормальном течении вспышки имеют примерно одинаковую изменчивость численности во времени в период массового размножения (рис. 66).
К первой фазе (начальной) вспышки относится только одно поколение вредителя, которое выкармливается в наступивших для него оптимальных условиях. Численность вредителя в насаждениях в этой фазе по сравнению с численностью предшествующего вспышке поколения увеличивается незначительно, чаще в два - четыре раза.
Во второй фазе (роста численности), охватывающей два - три поколения, численность вредителя продолжает возрастать, однако она еще невелика, наносимые вредителем повреждения в кронах не бросаются в глаза, и их можно обнаружить лишь при специальном осмотре. Личинки вредителя в данной фазе массового размножения отличаются повышенным содержанием жировых и белковых веществ и упитанностью, а куколки и яйца - крупными размерами.
В период второй и в начале третьей фазы происходит не только размножение и увеличение численности вредителя, но и его рассеивание по насаждениям.
При переходе в третью фазу {кульминации вспышки) численность вредителя резко увеличивается. Личинки (гусеницы) сильно объедают кроны деревьев, так что повреждение бросается в глаза и легко обнаруживается. Третья фаза длится два-три года. Постепенно личинки начинают испытывать недостаток в корме, что ведет к ослаблению их, снижению плодовитости у особей, заканчивающих питание и развитие, к распространению эпидемических заболеваний и гибели. В этой фазе заметно нарастает число энтомофагов, уничтожающих вредителя.
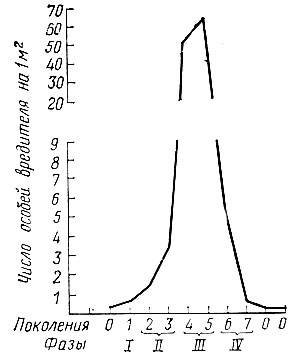
Рис. 66. Схема развития вспышки массового размножения хвое-листогрызущих насекомых по фазам (по Ильинскому, 1952)
В четвертой фазе (фаза кризиса) численность вредителя начинает быстро идти на убыль. Высокая плодовитость вредителя сменяется низкой и даже полным бесплодием. У соответствующих видов вредителей начинают преобладать самцы. Процент особей, пораженных энтомофагами и болезнями, все больше увеличивается. Численность вредителя падает до минимума, и вспышка заканчивается. Период четвертой фазы вспышки длится два-три года.
В период между вспышками численность вредителя держится на низком, но непрерывно колеблющемся уровне, а плодовитость близка к средней, но тоже колеблется.
Вспышка в целом в одном и том же насаждении развивается чаще всего в течение шести-семи лет. У вредителя с двойной генерацией (у обыкновенного соснового пилильщика) она протекает несколько быстрее, а при двухгодовом цикле - медленнее.
В природе могут наблюдаться значительные отклонения от приведенной схемы, обусловленные различными воздействиями внешних условий и спецификой реагирования на них каждого вида. Так, например, у зеленой дубовой листовертки вспышки массового размножения длятся 10 лет и более, повторяясь иногда одна за другой (рис. 67).
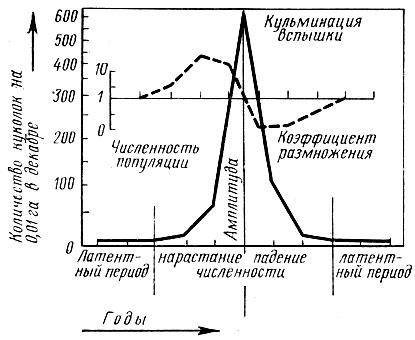
Рис. 67. Изменение численности сосновой совки в период массового размножения (по Швердтфегеру, 1957)
Об изменениях численности и ее уровне судят по плотности популяции (средняя или абсолютная заселенность), коэффициенту размножения, зараженности вредителя энтомофагами и болезнями, а также по внутреннему состоянию организма (плодовитость, масса куколок, число яиц в кладке и др.).
Коэффициентом размножения называют отношение плотности популяции в данном году к прошлому году или отношение двух смежных поколений. Чем выше коэффициент размножения, тем быстрее идет нарастание численности вредителя (рис. 68).
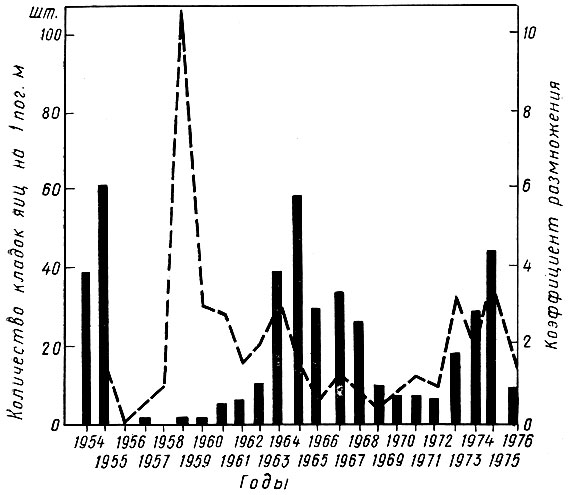
Рис. 68. Изменение численности дубовой зеленой листовертки за период 1954-1976 гг.
В местах роста численности популяций образуются очаги. Под очагом понимают заселенный вредителями участок леса, где их численность угрожает насаждению потерей 30% хвои и более (или 50% листвы и более) и где требуется решение о назначении борьбы. Различают очаги первичные, вторичные и миграционные.
Первичные очаги возникают в наиболее благоприятных условиях для данного вида вредителя. Чаще они появляются в чистых насаждениях, произрастающих на бедных почвах, вытоптанных скотом, нарушенных чрезмерным вмешательством человека, обеднённых видами животных и растений. Здесь рост численности идет очень быстро и она достигает максимальных размеров.
Вторичные очаги возникают в насаждениях, менее подходящих для размножения вредителя. В этих насаждениях больше насекомоядных птиц и энтомофагов, бонитет выше, чаще всего имеется подлесок и развитый травяной покров. Целостность таких насаждений нарушена меньше, обычно это бывают насаждения естественного происхождения. Численность популяции растет медленнее, уровень ее ниже. Наиболее сильные повреждения крон запаздывают по сравнению с первичными очагами на один-два года.
Миграционные очаги формируются вследствие разлета взрослых насекомых или переползания личинок. Судьба их зависит от условий, в которые попадают мигранты.
Географическое распространение
Большинство представителей хвое- и листогрызущих насекомых имеет очень широкие ареалы и встречается на огромных территориях, занятых их кормовыми породами.
В пределах европейской части СССР очаги хвое- и листогрызущих насекомых обычно наблюдаются в лесах степной и лесостепной зон. Эти зоны называются областью наиболее частых массовых размножений или зоной первичных вспышек (Ильинский, 1952). Вспышки массового размножения чаще всего приурочены к определенным лесорастительным условиям и многократно повторяются в одних и тех же лесных массивах. Наряду с этим есть лесные массивы, где ряд видов почти никогда не дает вспышек массового размножения, хотя они и лежат в центре ареала. Вероятнее всего это связано с устойчивостью насаждений, их составом и возрастом. Наконец, очень часто встречаются комплексные (сопряженные) очаги, в которых сразу развиваются вспышки целого ряда видов. Например, в лиственничных лесах Красноярского края наблюдаются сопряженные очаги сибирского шелкопряда, лиственничной пяденицы и непарного шелкопряда (Кондаков, 1965). В дубравах Поволжья часто возникают сопряженные очаги целого комплекса питающихся дубом видов - пядениц-шелкопрядов, листоверток, огневок и т. д.
Бывают и такие случаи, когда наблюдается известная закономерная смена видов во времени. Так, например, в одних и тех же районах Украины сначала появляется сосновый шелкопряд, а затем сосновые пилильщики, в затухающих очагах непарного шелкопряда возникают очаги дубовой зеленой листовертки и т. д.
Анализ статистического материала и наблюдения за развитием вспышек в природе показали, что в одних случаях они носят локальный характер и ограничиваются небольшой площадью, а в других - захватывают сразу очень большие пространства, часто распространяясь в пределах целой ландшафтно-географической зоны или даже в нескольких зонах. Такие вспышки массового размножения были названы пандемическими. Они характерны для наиболее распространенных, процветающих видов хвое- и листогрызущих насекомых.
Пандемическая вспышка может развиваться по территории не одновременно и не везде, так как в различных географических районах складываются местные типы погоды и различны лесорастительные условия. Такие вспышки повторяются довольно редко и в случае своего естественного затухания депрессия бывает очень глубокой.
Наряду с пандемическими встречаются локальные вспышки массового размножения, присущие почти всем хвое- и листогрызущим насекомым. Они обычно связаны с местными типами погоды, возникают на небольших площадях, в лесных массивах с наиболее благоприятными экологическими условиями. Эти вспышки часто бывают обусловлены хозяйственной деятельностью человека, истреблением полезных организмов, систематическими химическими обработками и другими местными причинами, ведущими к ослаблению насаждений и физиологическим сдвигам у кормовых пород.
При организации надзора и составлении прогнозов очень важно знать историю вспышек каждого вида, их приуроченность к определенным массивам, повторяемость и связь со вспышками других видов.
Некоторые хвое- и листогрызущие насекомые дают вспышки очень редко и они ограничиваются небольшими территориями. Объяснение этому пока не найдено. Наконец, есть виды с очень узкими ареалами, приуроченными к строго определенному физико-географическому району.
Влияние насекомых на состояние и прирост насаждений
Хвое- и листогрызущие насекомые, объедая хвою и листву на деревья, нарушают их нормальный водообмен и ассимиляцию что ведет к потере прироста и устойчивости.
Хвойные насаждения обычно резко снижают прирост под влиянием потери хвои, при повторных объединениях начинают усыхать и подвергаться нападению стволовых вредителей.
По данным Н. С. Грезе и В. Л. Циопкало (1936), объедание хвои гусеницами сосновой совки независимо от возраста насаждения (I - IYклассы) вызывает потерю прироста по диаметру до 50% в год повреждения и до 100% (в среднем 88%) в следующий после повреждения год. Если насекомые объедают майские побеги на деревьях в возрасте до 25 лет, то в том же году происходит их усыхание, а следующей весной из запасных почек образуются короткие боковые побеги. О. В. Трофимова, В. Н. Трофимов, А. Д. Орлинский (1979) указывают, что объедание сосновой совкой до 30% хвои практически не отражается на изменении категорий состояния деревьев в чистых сосновых насаждениях III - IV классов возраста. Повреждение до 60% хвои через один год становится визуально незаметным. Однократное сплошное объедание ослабляет насаждение, но не приводит к гибели. Охвоенность крон восстанавливается за два-три года, а отпад немного превышает естественный. При двукратном объедании насаждений с уничтожением 30 - 60% хвои в первый год и 80 - 100% во второй начинается процесс распада насаждения.
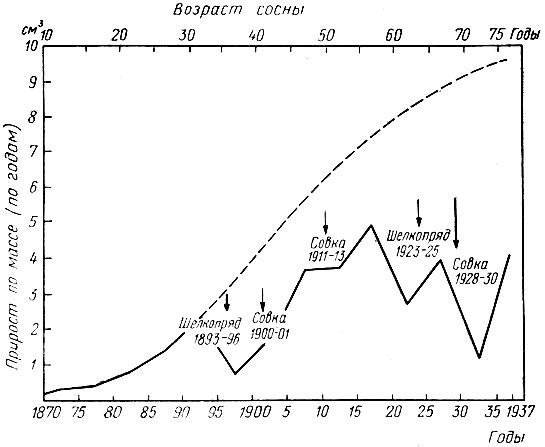
Рис. 69. Прирост сосны, поврежденной сосновой совкой и сосновым шелкопрядом, за период с 1870 по 1937 г. (по Маркусу, 1942)
Влияние на прирост сосны повреждений гусеницами сосновой совки и соснового шелкопряда в Германии за период с 1870 по 1937 г. показано на рис. 69 и 70 (Маркус, 1942). Характерно, что объедание хвои особенно сильно сказывается на прирост в последующие годы после размножения хвоегрызущих вредителей. Так, в годы вспышки сосновой пяденицы в Воронежской области (1955 - 1956) при 100%-ном объедании хвои потери прироста па диаметру составили 60%, а в последующие за вспышкой два года - 62 и 40%. Такая же картина наблюдалась и после повреждения сосновым шелкопрядом чистых сосновых культур 30-летнего возраста в Ленинском лесхозе Липецкой области, где в год повреждения (дефолиация была 80%) потеря прироста по диаметру составила 28%, а в последующий год 42% (Голосова, 1971).

Рис. 70. Влияние повреждений сосновой совкой и сосновым шелкопрядом на прирост сосны (по Маркусу). На торцевом срезе через ствол сосны видны годичные кольца. Начиная с 1931 г., эти кольца становятся узкими, что связано с повреждением хвои указанными вредителями
Потери прироста в сосняках-зеленомошниках (возраст 60 - 70 лет, полнота 0,7), поврежденных сосновым обыкновенным и желтоватым пилильщиками, определялись Ф. П. Моисеенко и А. М. Кожевниковым (1976) в Гомельской области. Ими установлено, что прирост деревьев, потерявших 40 - 50% хвои, в первый год после объедания в два раза меньше, чем у поврежденных. На второй год у этих деревьев он увеличился по сравнению с предыдущим годом на 20%. При потере хвои у деревьев более 90% прирост в первый год после объедания не образовался, а на второй год 20% деревьев усохло, у 65% деревьев прирост стал появляться и только 15% к концу вегетации и достигли прироста деревьев, поврежденных на 40 - 50%. В результате наблюдений (12 лет) авторы пришли к выводу, что потеря прироста по объему у средне поврежденных (в годы размножения пилильщиков) деревьев составила 20%, а у сильно поврежденных - около 43%.
После повреждения гусеницами монашенки чистые сосновые насаждения обычно не усыхают, но теряют во второй год сильного объедания и последующие два года 32-38% прироста по диаметру.
Повреждения звездчатого ткача обычно сильно сказываются на приросте сосновых культур и часто вызывают их частичное отмирание. Красноголовый ткач поедает преимущественно старую хвою и поэтому наносит меньший вред. Исследования прироста в поврежденных рыжим сосновым пилильщиком средневозрастных насаждениях Приморского лесхоза Ленинградской области показали, что за период массового размножения у сильно поврежденных деревьев прирост по диаметру был на 25% ниже, чем у слабо поврежденных в том же древостое, и на 26,7% ниже, чем до периода повреждения. Усыхания деревьев не наблюдалось (Стадницкий, 1976).
Устойчивость сосны к повреждениям хвоегрызущими вредителями зависит от условий местопроизрастания, состава, возраста, полноты, санитарного состояния насаждений, погодных условий и географического местоположения. После двукратного повреждения сосновой совкой и сосновым шелкопрядом насаждения обычно интенсивно усыхают.
Хорошо переносит повреждения лиственница. Она обладает высокой регенеративной способностью хвои. Ее брахибласты живут от 2 - 40 лет, ежегодно возобновляя хвою. Зачатки хвои будущего года развиваются на конусе нарастания брахибласта одновременно с хвоей текущего года, и при утрате ее вторичная хвоя отрастает в то же лето. Способность ежегодно сменять хвою обусловила высокую устойчивость лиственницы к дефолиации. При многократной потере хвои запасающиеся ткани обедняются пластическими веществами, в первую очередь углеводами, и деревья снижают защитные реакции, подвергаясь нападению стволовых вредителей. В связи с этим неоднократно наблюдалась гибель лиственничных насаждений в очагах сибирского шелкопряда. После объедания деревьев серой лиственничной листоверткой первичная хвоя на следующий год появляется позднее и ее биохимический состав неблагоприятен для питания гусениц. Поэтому деревья подвергаются лишь однократной дефолиации, бабочки непрерывно мигрируют по территории массового размножения и очаги ежегодно смещаются. По исследованиям А. С. Плешанова (1972) в результате однократной полной дефолиации прирост на деревьях сократился в год повреждения на 10 - 35%, и на следующий год на 6 - 21,7%. Заселение стволовыми вредителями деревьев после их однократной дефолиации не происходит.
Ель и пихта сильнее, чем сосна и лиственница, реагируют на потерю хвои, скорее подвергаются нападению короедов и усыхают. Заселение короедами и хвойными черными усачами часто начинается даже после однократного сильного повреждения хвои, а после двукратного насаждения быстро отмирают. В настоящее время накопилось много данных о влиянии дефолиации хвойных пород на их прирост и состояние, которые обобщены в книге А. И. Воронцова. Патология леса. М., 1978.
Лиственные насаждения переносят повреждения листвы насекомыми лучше, чем хвойные, но потери прироста тоже достигают больших размеров, особенно при весеннем питании гусениц. Повреждения листвы дуба вредителями летне-осенней группы почти не затрагивают прирост текущего года и лишь незначительно сказываются на приросте следующего года.
Однократная сильная дефолиация, как правило, не оказывает влияния на состояние насаждений, они легко оправляются и на следующий год нормально функционируют. При двукратной сильной дефолиации и полной потере листвы ухудшается общее состояние древостоя. У многих деревьев в кронах появляются сухие ветви, на стволах возникают водяные побеги, иногда происходит даже частичное отмирание кроны. После трехкратной дефолиации начинается отмирание деревьев в насаждении и заселение их стволовыми вредителями. Здесь, как и в хвойных насаждениях, большую роль играют погодные условия, географическое местоположение насаждений, их состав и возраст.
Уничтожение насекомыми листвы деревьев приводит не только к потере прироста и ослаблению. Оно вызывает последовательные, но большей частью временные изменения в жизни экосистемы. Увеличение прозрачности верхнего полога в необычный фенологический срок вызывает резкие изменения освещенности, температуры и влажности, приближающие условия под пологом леса к условиям открытого места. В этих насаждениях резко увеличивается мощность травяного покрова, улучшается состояние подроста и самосева древесных пород, усиливается плодоношение (Воронцов, Иерусалимов, Мозолевская, 1967).
При дефолиации в насаждениях наряду с изменением фитоклимата меняется и поступление питательных веществ в почву за счет экскрементов и трупов гусениц. По данным А. А. Молчанова (1971), содержание минеральных веществ и азота энтомогенного происхождения от количества в общем опаде листвы экскрементов и трупов гусениц в 228-летних дубовых насаждениях Телермановского лесничества, поврежденных непарным шелкопрядом, составляло (%): азота - 39,6, фосфора - 33,2, калия - 38,7, кальция - 2,12, магния - 39,1. По данным Р. И. Злотина и К. С. Худашевой (1974), опад листьев дуба за вспышку дубовой зеленой листовертки уменьшился всего в 1,4 раза, а суммарный запас мертвых напочвенных остатков - более чем в четыре раза. За вегетационный период до вспышки в среднем разлагалось 28% суммарного запаса листовой подстилки и опада. За то же время в годы массового размножения листовертки убыль массы этого запаса составляла в среднем 75%. Таким образом, в очагах листовертки разложение мертвых растительных остатков протекало в три раза быстрее.
Ускоренное разложение подстилки и опада в период вспышки определяется несколькими причинами. Одна из них заключается в усилении абиотической минерализации за счет значительного количества солнечной радиации, проникающей под полог осветленных (в силу потери листвы, съеденной листоверткой) насаждений. Солнечная радиация изменяет гидротермический режим в слое подстилки, что способствует выносу органических и химических соединений.
Очень большое значение имеет зоогенный опад (экскременты). Продукция экскрементов листовертки за период ее личиночного развития достигает 6 ц/га, а в среднем составляет 3 ц/га (воздушно-сухая масса). Экскременты являются водопрочными образованиями и по сравнению с одинаковой массой листового опада имеют примерно в 50 раз большую поверхность, которая служит сферой активного развития сапрофильных микроорганизмов. Особый химический состав экскрементов листовертки, а именно узкое отношение углерода к азоту, также увеличивает активность микробной плазмы. Это отношение в экскрементах составляет 20, а в подстилке оно увеличивается и достигает 42. В падающих на почву огрызках зеленых листьев отношение углерода к азоту уменьшается до 11. Известно, что для развития микроорганизмов наиболее благоприятны органические соединения, в которых отношение C:N не превышает 20.
Таким образом, массовое размножение листогрызущих насекомых стимулирует процессы минерализации растительного опада и способствует более интенсивному протеканию биологического круговорота в результате быстрого освобождения значительного количества вещества и энергии, заключенных в лесной подстилке.
Защита насаждений
Надзор
Необходимость защиты насаждений, сроки и объем борьбы определяют по данным специального надзора и лесопатологических обследований.
Специальный надзор за хвое- и листогрызущими насекомыми проводят в насаждениях, типичных для возникновения вспышек массового размножения вредителей этой группы. (Сущность и организация надзора описаны в гл. IV.) Для определения плотности популяции и прогноза численности вредителей особенно важен детальный надзор. В насаждениях, выбранных под надзор за тем или другим видом вредителя, ежегодно проводят учет численности. Для этого определяют плотность популяции, встречаемость, плодовитость, соотношение полов, основные факторы смертности и общую жизнеспособность особей по их массе, размерам, окраске, анализу гемолимфы.
Методы определения плотности популяции при надзоре и лесопатологических обследованиях одинаковы. Они зависят от биологии и экологии вида вредителя, не следует вести учет по подвижным, активно перемещающимся фазам имаго и личинки во время питания. Основной учет проводят преимущественно осенью по зимующей фазе и дополняют промежуточными на других фазах развития. Учет численности вредителей проводят в лесной подстилке и почве, в кроне и на стволах деревьев.
Учет вредителей, окукливающихся или зимующих в лесной подстилке и почве, проводят на прямоугольных площадках размером 0,25; 0,5; 1,0 м2.
Учет вредителей в кроне ведут на модельных деревьях. Крону дерева условно делят на три слоя (нижний, средний, верхний) и из каждого берут по одной модельной ветви определенной длины. На них подсчитывают количество кладок яиц, личинок или зимующих гнезд. Иногда ведут учет на 100 (или 1000) листьев или почек, на площадь или массу листвы (хвои), на всю крону. В последнем случае ветви постепенно спускают на расчищенную площадь или мешковину и подсчитывают всех личинок (гусениц).
Учет вредителей на стволах деревьев ведут обычно по кладкам яиц (непарный шелкопряд, монашенка). Если они сосредоточены в комлевой части ствола, то подсчитывают их на всем дереве без валки моделей. В случае размещения кладок яиц по всему стволу валят модельные деревья и считают на нескольких метровых отрезках.
Учет численности хвое- и листогрызущих насекомых трудоемок. Между тем от его точности зависит прогноз ожидаемой дефолиации, а значит, и правильность решения о назначении борьбы. Поэтому в течение последних лет проводились многочисленные исследования методов учета с широким привлечением биологической статистики1. Подробнее с методами учета студенты знакомятся на учебной практике.
1 (В списке литературы к этой главе приводятся основные работы, отражающие достижения в области совершенствования методов учета.)
По данным надзора осенью составляется прогноз о размножении вредителей на следующий год. Если численность быстро увеличивается, а зараженность энтомофагами и болезнями незначительна (менее 30%), проектируется метод борьбы. Обоснованием для метода борьбы служит также показатель степени ожидаемой потери хвои или листвы насаждением при повреждении на следующий год. Он вычисляется по таблицам, составленным А. И. Ильинским (1965). В них для каждого класса возраста насаждения приводится критическое число яиц, гусениц или куколок на единицу площади или одно дерево, при котором ожидается 100%-ное объедание насаждения. В зависимости от характера насаждения (возраста, состава, полноты), степени его устойчивости и вида вредителя борьбу назначают при разных степенях ожидаемого объедания. В хвойных насаждениях активные меры борьбы чаще всего проводят уже при 30 - 50%-ной потере хвои, в лиственных - только в случае угрозы 100%-ного объедания и высокого коэффициента размножения.
Долгосрочное прогнозирование динамики численности хвое- и листогрызущих насекомых еще далеко от совершенства. В настоящее время усиленно развивается моделирование процессов динамики численности популяций и строятся прогнозирующие модели (А. И. Воронцов, 1978; А. В. Голубев и др., 1980).
Лесохозяйственные мероприятия. Применение истребительных мер борьбы не может гарантировать длительное снижение численности популяции вредителя ниже порога вредности. Предотвращение образования очагов хвое- и листогрызущих насекомых возможно только при сочетании химического метода борьбы с лесохозяйственными мероприятиями, направленными на создание неблагоприятных условий для их размножения. Эти мероприятия нужно предусматривать заранее, в процессе выращивания насаждений и ухода за ними.
Необходимо стремиться к созданию смешанных сложных равномерно сомкнутых насаждений. Такие насаждения менее всего благоприятны для развития свето- и теплолюбивых насекомых. Они наиболее гармонично и полно используют окружающие условия внешней среды, концентрируют наибольшее количество полезных организмов и поэтому обладают необходимой биологической устойчивостью.
При подборе древесных пород необходимо вводить в насаждения: породы, наименее повреждаемые самыми распространенными в данном географическом районе хвое- и листогрызущими насекомыми; породы, повреждаемые самыми распространенными хвое- и листогрызущими насекомыми, но запаздывающие по сравнению с ними в развитии весной (поздно распускающаяся форма летнего дуба и др.); породы, питание которыми неблагоприятно отражается на развитии ряда хвое- и листогрызущих насекомых, вызывает у них нарушение обмена веществ и снижает выживаемость популяции (например, при питании непарного шелкопряда липой и кленом остролистным).
На бедных песчаных почвах, где не хватает азота, нужно повышать устойчивость насаждений культурой люпина многолистного путем высева в междурядьях. Люпин обогащает почву азотом, влияет на физиологические процессы в хвое, усиливает смоловыделение, затеняет почву и во время цветения привлекает энтомофагов.
Положительное влияние на устойчивость насаждений оказывает кустарниковый ярус, отеняющий почву и часто мешающий полету бабочек и откладке ими яиц в нижней части деревьев. Кустарниковый ярус и густые опушки привлекают насекомоядных птиц на гнездование, служат для них и полезных насекомых источником дополнительного питания.
В чистых сосновых лесах можно создавать небольшие участки из лиственных пород, кустарников и травянистой растительности и насыщать их энтомофагами и насекомоядными птицами. Такие участки площадью 100 - 200 м2 делают по одному на 3 - 5 га. В ПНР они получили название ремизов. Опыт создания подобных участков проведен Челябинской станцией защиты леса и в Белоруссии.
В сосновых культурах с мертвым слоем лесной подстилки запрещается ее сгребание, так как при этом ухудшаются лесорастительные условия и уничтожается ряд эффективных паразитов хвое-и листогрызущих насекомых. Необходимо соблюдать предосторожность при проведении любых лесохозяйственных мероприятий в лесу, с тем чтобы сохранить удобные места гнездования и гнезда птиц. При уходе за лесом и санитарных рубках нужно оставлять дуплистые деревья и сохранять подлесок. Для гнездования птиц развешивают также искусственные гнездовья (см. гл. IV).
Использование энтомофагов
Для ликвидации небольших очагов хвое- и листогрызущих вредителей используют энтомофагов путем их сезонной колонизации или внутриареального расселения (см. гл. IV).
Для борьбы с сосновым шелкопрядом используется яйцеед-теленомус (Telenomus verticillatus), которого размножают на яйцах соснового шелкопряда в лабораторных условиях. Для этого заблаговременно накапливают яйца соснового шелкопряда, затем их заражают теленомусом в специальных вивариях. Зараженные яйца размещают в развивающихся очагах соснового шелкопряда перед лётом бабочек. Накопление яиц соснового шелкопряда и разведение теленомуса весьма трудоемко, и без соответственной механизации получить большое количество паразитов трудно.
Для борьбы с сибирским шелкопрядом используется телономус стройный (Telenomus gracilis). Зараженные теленомусом яйца сибирского шелкопряда собирают в затухающих очагах и переносят в возникающие микроочаги. При недостатке зараженных яиц теленомуса разводят в лаборатории, где выкармливается популяция гусениц сибирского шелкопряда. В этом случае выпускают прямо из банок имаго теленомуса во время лёта бабочек шелкопряда.
Для борьбы с кольчатым коконопрядом используют яйцеедов-теленомусов. Их несколько видов. Наиболее специализирован и эффективен Telenomus laeviusculus. Его цикл развития в основном синхронен с циклом хозяина. Кладки яиц кольчатого коконопряда собирают рано весной путем обрезки веток, на которых они находятся. Их помещают в изолированные помещения и ждут выхода гусениц. После этого оставшиеся кладки, из которых не вышли гусеницы, готовят к развешиванию на деревьях, связывая по две ветки. Через две-три недели после выхода гусениц кладки, заряженные теленомусом, развешивают в возникающих очагах вредителя. При хранении веточек с кладками в помещениях необходимо их периодически увлажнять, чтобы не произошло высыхания яиц. В дополнение к описанному способу использования теленомусов для борьбы с кольчатым коконопрядом рекомендуется подготовленные к развешиванию кладки яиц, зараженных теленомусом, в апреле на 10 дней подвергнуть воздействию более высоких температур (около 22° С). При этом выход теленомуса из зараженных яиц начинается раньше массового лёта хозяина. На 1 га нужно иметь около 10 тыс. теленомусов, для чего следует развесить 200 - 500 кладок яиц, зараженных теленомусом. При этом на 2 га тратится один рабочий день.
Для борьбы со златогузкой рекомендуется использовать эффективного специализированного паразита гусениц птеромалида - Eupteromalus nidulans. Для этого ранней весной в очагах златогузки роют однометровые ямы на глубину 50 см (4 - 5 на 1 га). На дно ям помещают для дренажа тонкие ветки, а затем зимние гнезда златогузки. В одну яму закладывают около 200 гнезд. На 1 га должно быть не менее 30 тыс. шт. паразитов, которые разлетаются из гнезд и заражают гусениц. Края ям обрабатывают хлорофосом, чтобы не расползлись незараженные гусеницы. Гнезда, в которых гусеницы сильно заражены паразитами, собирают перед закладкой их в ямы или с осени. В этом случае они хранятся всю зиму при температуре около 4° С. Гнезда златогузки собирают в затухающих очагах и переносят в возникающие.
О большой пользе муравьев рода формика как истребителей вредных лесных насекомых уже говорилось выше (см. гл. IV). Наибольшее значение они имеют в чистых сосновых и дубовых насаждениях, где отсутствует подлесок, мало развит травяной покров и поэтому мало энтомофагов. В этих условиях особенно важно организовать охрану муравьев и расселение их в те насаждения, где их недостаточно. Особенно эффективно муравьи уничтожают гусениц младших возрастов сосновой совки, сосновой пяденицы, соснового шелкопряда, монашенки, пилильщиков, дубовой зеленой листовертки, серой лиственничной листовертки и зимней пяденицы.
Использование биопрепаратов. Большинство видов хвое- и листогрызущих насекомых восприимчиво к бактериальным препаратам (описанным в гл. IV). Поэтому рекомендуют их использовать для борьбы с этой группой вредителей авиационным методом или с помощью наземных машин для борьбы с вредителями.
Для борьбы с сибирским шелкопрядом нужно использовать дендробациллин и инсектин; в борьбе с сосновым шелкопрядом эффективнее гомелин. Можно использовать любые бактериальные препараты для борьбы с сосновой пяденицей, кольчатым коконопрядом, дубовой зеленой листоверткой.
Технология борьбы не отличается от таковой при использовании пестицидов. Наиболее эффективен авиационный метод.
Недостаточно эффективны бактериальные препараты в борьбе с непарным шелкопрядом и сосновым пилильщиком. Зато эти насекомые восприимчивы к вирусному заболеванию - полиэдрозу. В связи с этим для борьбы с непарным шелкопрядом создан специальный вирусный препарат вирин-ЭНШ. Он применяется в виде водной суспензии для инфицирования корма гусениц (при авиационной обработке насаждений) или кладок яиц. Препарат разводят в 0,02 - 0,04%-ном растворе вспомогательного вещества ОП-7. При авиационной обработке расход препарата составляет 0,1 кг на 1 га. Для обработки кладок яиц готовят 0,1%-ную суспензию по препарату. Опрыскивание кладок яиц делают перед выходом гусениц весной. Для этого в очаге выбирают несколько участков, составляющих 2 - 3% общей площади. Их обрабатывают и таким образом создают источники инфекции. В распространении инфекции большую роль играют тахины - паразиты непарного шелкопряда.
В борьбе с сосновыми пилильщиками применяют вирин-диприон. Расход препарата при авиаопрыскивании 20 - 40 мл/га, рабочей жидкости 50 л/га.
Использование инсектицидов
В настоящее время с помощью биопрепаратов еще не всегда можно вовремя подавить очаги хвое-и листогрызущих насекомых. Поэтому в случае возникновения крупных очагов вредителей (сосновый и сибирский коконопряды, сосновая совка и др.) необходимо проводить химические меры борьбы. Метод борьбы зависит от характера насаждения, его расположения и площади, на которой распространился вредитель. На больших площадях и в малодоступных насаждениях проводится авиахимическая борьба. В небольших очагах, полезащитных полосах, зеленых насаждениях городов предпочитают обработку насаждений с помощью наземной моторной аппаратуры.
Авиахимическую борьбу с хвое- и листогрызущими вредителями проводят против личинок (гусениц) младших возрастов, приурочивая начало работ к их отрождению из яиц. Для установления календарных сроков борьбы проводят фенологические наблюдения.
Для борьбы с хвое- и листогрызущими вредителями применяют преимущественно фосфорорганические инсектициды и препараты гексахлорана (его гамма-изомера). Их используют в форме масляных и водных растворов, эмульсий, суспензий и изредка дустов. Оптимальный расход препаратов гамма-изомера гексахлорана по действующему веществу колеблется в пределах 0,2 - 0,3 кг на 1 га. Нормы расхода масляных растворов составляют 5-8 л на 1 га, масляных эмульсий - 12 - 20 л на 1 га. Оптимальный расход хлорофоса (по действующему веществу) - 0,8 - 1,2 кг на 1 га, норма расхода 3 - 5%-ных суспензий и водных растворов - 20 - 30 л на 1 га.
Соответственно концентрация их будет для эмульсий гамма-изомера 1 - 2%-ная, а для водных растворов и суспензий хлорофоса 2 - 5%-ная. Самые низкие концентрации (2 - 4%-ные) используются против сосновых пилильщиков, сосновой и пихтовой пядениц, а самые высокие (4 - 5%-ные) - против непарного и кольчатого шелкопрядов.
В Списке разрешенных Гослесхозом СССР химических средств борьбы нормы расхода инсектицидов указаны по препарату. В этом случае на 1 га расходуется 1,5 - 3 кг 16%-ной минерально-масляной эмульсии. В этом же Списке рекомендованы для борьбы с рядом хвое- и листогрызущих насекомых бензофосфат (1 - 3 кг на 1 га), карбофос (1,2 - 4 кг на 1 га), метафос (1 - 4 кг на 1 га). Нормы расхода по препарату.
Рабочие жидкости, особенно суспензии и эмульсии, нужно приготавливать непосредственно перед применением. Для этого на загрузочной площадке аэродрома необходимо иметь соответствующие емкости (баки, чаны, цистерны и др.), мотопомпы с запасом горючего и воду, которую подвозят на автоцистернах.
Наиболее перспективно авиационное ультрамалообъемное опрыскивание (УМО), при котором жидкий пестицид без разбавления в тонкодисперсном состоянии наносят на обрабатываемую поверхность. Для УМО используют 30%-ный раствор рицифона в этилцеллозольве (2 - 2,6 кг/га) и 40%-ный карбофос в том же растворителе (1,5 - 2 кг/га).
При организации борьбы с хвое- и листогрызущими насекомыми необходимо стремиться к комплексности и проводить интегрированную защиту. Не следует увлекаться химическими обработками насаждений, а проводить их только в самых необходимых случаях, опираясь на точные данные прогноза. Для этого следует совершенствовать технику надзора и учета численности хвое- и листогрызущих насекомых. Применять инсектициды нужно в сроки, когда меньше всего уничтожаются энтомофаги. Для этого надо хорошо знать их фенологию. Так, обработки инсектицидами для защиты дубрав от зеленой дубовой листовертки, непарного шелкопряда и других гусениц, появляющихся рано весной, нужно проводить как можно раньше, когда еще не начался лёт энтомофагов.
Интегрированные системы лесозащитных мероприятий строят на экологической основе и разрабатывают для каждого вида вредителя и соответствующего региона. При квалифицированной помощи станций защиты леса в системы можно включить инсектициды и биопрепараты, проходящие еще опытно-производственную проверку, а также феромоны.
При химической и интегрированной борьбе с хвое- и листогрызущими вредителями нужно строго соблюдать все правила техники безопасности при работе с пестицидами и биопрепаратами.
Обзор отдельных видов
Чешуекрылые (Lepidoptera)
Листовертки (Tortricidae)
Небольшие бабочки, в размахе крыльев не больше 25 мм. Передние крылья широкие, почти прямоугольные, их задний и наружный края образуют угол. В покое крылья складываются кровлеобразно вдоль спины. Гусеницы длиной 10 - 20 мм, 16-ногие, с твердыми хитинизированными переднегрудным и анальным щитками; на теле небольшие бородавки, несущие короткие волоски. Гусеницы очень подвижны; при прикосновении характерны резкие, извивающиеся движения тела, передвижение назад, быстрое опускание вниз на паутинной нити. Куколки перед вылетом бабочек при помощи шипиков на брюшке выдвигаются из паутинных коконов.
В лесах встречается очень много видов листоверток, повреждающих шишки и плоды' (гл. V), почки и побеги молодняков хвойных пород (гл. VII), листья и побеги лиственных пород в насаждениях разного возраста. Наиболее опасные вредители - серая лиственничная листовертка, и дубовая зеленая листовертка. Кроме того, в лесопарках и дубравах степной зоны широко распространены следующие виды листоверток, численное соотношение которых обычно меняется по годам: розанная листовертка (Cacoecia rosana L.), боярышниковая (С. crataegana Hb.), пестро-золотистая (С. xylosiea-па L.), сетчатая (С. reticulana Hb.), заморозковая (Exapate com gelatella CI.).
Серая лиственничная листовертка (Zeiraphera diniana Gn.). Бабочка в размахе крыльев 20 - 22 мм. Передние крылья желтовато-серого цвета в неясных штрихах и темных пятнах, с двумя перевязями и неправильным подвершинным пятном темно-бурого цвета. Задние крылья буровато-серые.
Лёт начинается в середине июля и продолжается до конца августа. Самка откладывает яйца под чешуйки коры укороченных побегов и тонких веточек группами по 2 - 12 шт. Плодовитость одной самки до 200 яиц. Яйца бледные, лимонно-желтые, с морщинистой поверхностью, диаметр около 0,5 мм. Яйца зимуют.
Гусеницы выходят в период распускания хвои лиственницы, обычно во второй половине мая. В первом возрасте они питаются на цветках и почках, располагаясь в рыхлом паутинном мешочке среди молодых хвоинок. По мере дальнейшего роста гусеница соединяет хвоинки вместе в пучок в виде трубочки и там живет, минируя и выскабливая их. Более взрослые гусеницы живут открыто и поедают хвою полностью. Они зеленовато-серые с черной головой.
При массовом скоплении гусениц в кроне выделенные ими паутинные нити опутывают ветви и задерживают остатки поврежденной хвои и кал. Кроме лиственницы в Западной Европе повреждается ель. Гусеницы проходят пять возрастов. Окукливание в конце июня. Куколка блестящая, коричневая, с редкими желтоватыми волосками, 10 - 12 мм. Фаза куколки 15 - 17 дней. Генерация одногодовая.
Вспышки массового размножения листовертки возникают преимущественно в горных районах с большими массивами чистых лиственничных насаждений. Первичные очаги приурочены к изреженным старым, хорошо прогреваемым насаждениям. Наряду с локальными периодически возникают вспышки массового размножения на огромных площадях. При этом наблюдаются массовые миграции бабочек на большой высоте из насаждений поврежденных в неповрежденные. Очаги носят пятнистый характер. Сильное объедание хвои гусеницами бывает один-два года, затем очаги перемещаются. В целом же пандемические вспышки длятся семь-восемь лет. Гибели насаждений в местах повреждений их гусеницами не наблюдалось, встречалось лишь куртинное усыхание и снижение прироста.
Надзор проводится в июне-июле по повреждениям крон гусеницами. Для учета берут модельные ветви и подсчитывают количество гусениц на 1 м ветви. Полное объедание можно ожидать при 300 гусениц на 1 м ветви, а переход вспышки в фазу кульминации - при 15 - 20 (Ильинский, 1965).
Повреждения хорошо обнаруживаются с самолетов и вертолетов при воздушном патрулировании. В июле надзор ведется по бабочкам и помощью светоловушек.
Дубовая зеленая листовертка (Tortrix viridana L.). Бабочка в размахе крыльев 18-23, мм. Передние крылья и грудь ярко-зеленые, задние крылья и брюшко серые (рис. 71).
Лёт в июне, обычно при средней дневной температуре выше 17° С. При снижении до 13° С и вечернего минимума до 4° С бабочки прекращают лёт и прячутся среди листьев. В дневные часы, когда освещенность более 30 000 лк, бабочки летают в пределах всей кроны дерева, а до восхода солнца и вечером сидят неподвижно в нижней части, на кустарниках и траве. Во время яйцекладки самки держатся верхней части крон и откладывают по одному - три (чаще два) яйца на утолщениях между годичными побегами стержневой ветви на рубчиках и у основания черешков листьев, в развилках веточек и других местах, где имеются неровности коры.
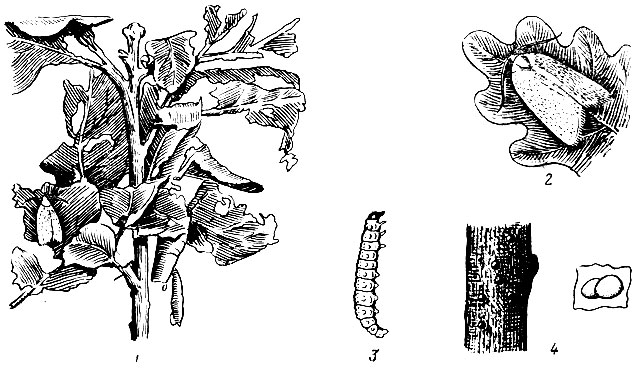
Рис. 71. Дубовая зеленая листовертка: 1 - ветвь дуба с поврежденными листьями (видны сидящая бабочка и куколка, из которой уже вышла бабочка), 2 - бабочка, 3 - гусеница, 4 - яйца на ветви, рядом увеличенные яйца
Кладки яиц величиной около 1 мм покрыты сверху щитками под цвет дубовой коры, образовавшимися из специальных выделений половых желез самки. Яйца под щитком оранжевые округлой формы, диаметром около 0,7 мм, сверху приплюснутые. Они остаются зимовать до весны следующего года.
Гусеницы появляются в конце апреля - начале мая. Обычно их появление совпадает с распусканием почек ранней формы летнего дуба. Гусеницы проникают в почки и выедают их, затем скелетируют листья, а в старших возрастах съедают целиком.
Большинство исследователей считают, что гусеницы младших возрастов могут питаться только дубом, а в последнем возрасте способны переходить на питание листьями липы, клена, ольхи и других лиственных пород. В лабораторных условиях удавалось воспитывать гусениц от момента их отрождения до окукливания на липе и клене. При этом наблюдалась большая смертность гусениц и низкая последовательность бабочек.
Гусеницы линяют четыре раза и имеют пять возрастов. Развитие гусеницы в среднем длится 20 - 25 дней. Популяция гусениц обычно разновозрастная, и они встречаются в насаждениях 30 - 45 дней, после чего происходит массовое окукливание в листьях. Гусеница подгибает край листовой пластинки на нижнюю сторону и пришивает паутиной, образуя "пирожок". Он изнутри выстлан тонкой паутиной в виде мешка, открытого с одной стороны; через этот конец мешка куколка перед вылетом бабочки выдвигается наружу; после вылета бабочки видна шкурка куколки.
Куколка темно-бурая, иногда почти черная. Вершина брюшка притуплена, а на заднем крае сидят восемь коротких слабо согнутых на брюшную сторону щетинок. Длина 9 - 10 мм.
Дубовая зеленая листовертка свето- и теплолюбива. Она заселяет в первую очередь освещенные опушечные или отдельно стоящие деревья, изреженные насаждения, парки, лесополосы ажурной конструкции.
Очаги образуются в изреженных чистых дубовых насаждениях, занимающих наиболее сухие и прогреваемые места, а также в приовражных байрачных дубравах, в парковых рощах и других насаждениях вокруг больших городов, во всей европейской части СССР, на Кавказе и в Крыму, исключая север и Урал. Очаги, как правило, приурочены к насаждениям из ранней формы летнего дуба.
С дубовой листоверткой связано много энтомофагов. В ряде случаев они оказывают существенное влияние на изменение численности популяции, уничтожая в конце вспышки до 68% всех особей. (Смирнов, 1960). Такую же роль играют птицы, особенно скворцы. Однако те и другие недостаточно регулируют численность листовертки. Быстрое нарастание ее численности возможно только при условии оптимального совпадения сроков выхода гусениц из яиц и появления фазы открытой почки у дуба. Если последующий год складывается тоже более или менее благоприятно для питания гусениц, то вспышка массового размножения дубовой листовертки в последующие три-четыре года достигает максимума и продолжается длительный период (до 15 лет).
Затухание вспышек массового размножения листовертки происходит сразу. Оно обычно обусловлено очень низкими температурами зимой. Под Москвой вспышки массового размножения листовертки, как правило, резко обрываются низкими температурами в январе - феврале. То же наблюдается и в других местах.
Гибель подавляющего числа гусениц в яйцах во время диапаузы происходит при температуре минус 30 - 35° С. Низкие температуры в декабре менее губительны для яиц зеленой листовертки. Весной гусеницы гибнут при температурах минус 6 - 8° С.
Надзор в очагах листовертки ведется по кладкам яиц, которые учитываются на стержневых побегах длиной 1 м или по трем ветвям. Кроме того, надзор возможен по летающим бабочкам.
Боярышниковая листовертка (Cacoecia crataegana Hb.). Бабочка в размахе крыльев 19-26 мм. Передние крылья серо-бурые или фиолетово-серые с темным рисунком в виде поперечных полос и пятен на вершинном крае.
Лёт во второй половине июня. Самка откладывает яйца кучками на стволы деревьев. Кладка яиц представляет собой слегка приподнятый плоский щиток овальной формы, в нем яйца плотно прилегают друг к другу и расположены в один ряд. Яйца цементируются белым, воскоподобным веществом. В одной кладке 10 - 90 яиц. Одна самка может отложить одну - три кладки. Яйца зимуют. Гусеницы появляются рано весной, обычно на декаду позднее, чем у зеленой дубовой листовертки. Развитие гусениц продолжается около месяца, они линяют четыре раза и проходят пять возрастов. Гусеница зеленовато-черная с черными бородавками и головой. Фаза куколки длится 10 - 12 дней. Куколки помещаются в листья, которые закручены в трубку с вершины или бокового края. Генерация одногодовая.
Очаги боярышниковой листовертки возникали в дубовых насаждениях степной и лесостепной зон. Гусеницы повреждают дуб, вяз, осину, липу, клен остролистный. Излюбленные породы - дуб и липа, а в садах - плодовые, где она наносит большой вред.
Изменение численности боярышниковой листовертки изучено мало. Однако она более устойчива к зимним морозам, а весной не связана синхронно с распусканием почек дуба, так как питается и на других породах.
Надзор проводится в мае по повреждениям и осенью по кладкам яиц на стволах деревьев.
Хохлатки (Notodontidae)
Ночные бабочки средней и крупной величины с массивным телом и относительно узкими крыльями, которые в спокойном состоянии складываются кровлеобразно. Гусеницы голые или слабоволосистые с пятью парами брюшных ног. Куколки черные, обычно без кокона, в земле. Многие виды приносят в фазе гусеницы большой вред лиственным насаждениям. Наибольшую известность имеют: лунка серебристая, дубовая хохлатка, ильмовый ногохвост. Часта встречается также хохлатка зигзаг и березовая зубчатая хохлатка.
Дубовая хохлатка (Notodonta anceps Goeze). Бабочка в размахе крыльев 4,5 - 6,0 см. Передние крылья серого цвета с тремя неясными зубчатыми линиями и рядом ржаво-бурых пятен перед: внешним краем. Задние крылья желтовато-белые (рис. 72).
Лёт в мае, когда среднесуточная температура воздуха достигает 17° С, а сумма эффективных температур 360 - 370° С. Самки откладывают яйца на тонких (до 1 см) веточках в кронах и на стволах деревьев в один слой. Плодовитость одной самки до 300 яиц, максимальная - 496 (Егоров и Соложеникина, 1960).
Яйцо молочно-белого цвета полушаровидной формы с плоским основанием, в диаметре 1,4 мм. Фаза яйца длится около 10 дней по одним наблюдениям (Егоров и Соложеникина, 1960) и до трех недель по другим (Ильинский, 1965). Вышедшие из яиц гусеницы скелетируют листья дуба, во втором возрасте начинают объедать их с краев, а затем съедают целиком, оставляя толстые жилки. Гусеница до 6 см длиной, голая, цилиндрическая, зеленого цвета, с двумя узкими светлыми полосами на спине и темной полосой между ними. По бокам тела косые красные и желтые полосы. Голова зеленоватая с четырьмя желтыми черточками. Питаются гусеницы только листьями дуба. Они линяют четыре раза и имеют пять возрастов. Общая продолжительность развития 36 - 43 дня.
Окукливание происходит в начале июля под кронами в лесной подстилке на глубине до 5 см. Куколка черная, блестящая, в мягком продолговатом коконе из остатков листьев с прилипшими частицами почвы. Длина куколки 3 - 4 см, средняя масса 0,2 г (самки), 0,9 г (самца). Куколка зимует. Генерация одногодовая, однако в фазе куколки часто наблюдается частичная диапауза, которая может иногда длиться несколько лет.
Дубовая хохлатка - вид тенелюбивый. Ее очаги возникают обычно в молодых и средневозрастных дубовых насаждениях преимущественно порослевого происхождения, произрастающих на супесчаных и щебенчатых бедных почвах в зоне лесостепи и степи. Очаги носят локальный характер, обычно возникают в одних и тех же местах и вследствие диапаузы у куколок длятся в течение ряда лет.
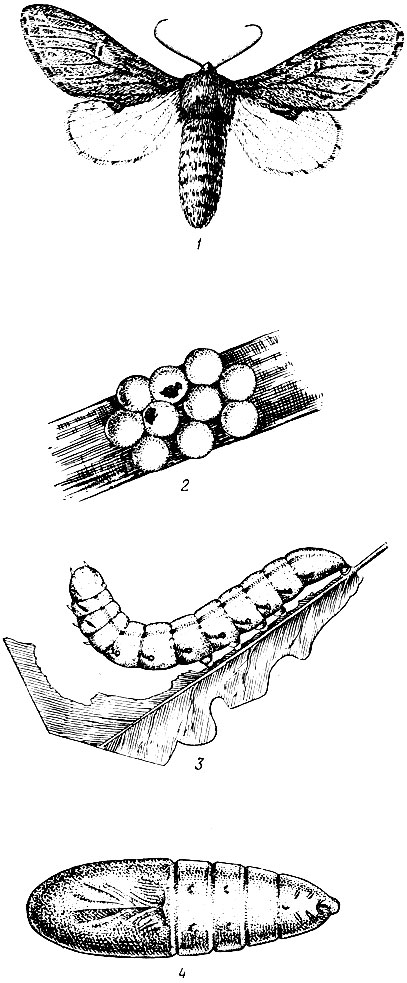
Рис. 72. Дубовая хохлатка:1 - бабочка, 2 - кладка яиц, 3 - гусеница, 4 - куколка
На колебание численности большое влияние оказывают эпизоотии у гусениц и птицы, активно истребляющие гусениц и куколок. Из энтомофагов энергично истребляют гусениц большой зеленый красотел. Надзор нужно проводить осенью, учитывая число куколок в почве.
Лунка серебристая (Phalera bucephala L.). Бабочки в размахе крыльев 5 - 6 см. Передние крылья серебристо-серые. В наружном углу каждого крыла расположено лунообразное золотисто-желтое пятно, с внутренней стороны окаймленное дугообразной двойной темной линией. Задние крылья белые с легким сероватым оттенком (рис. 73).
Массовый лёт бабочек в июне. Отдельные экземпляры встречаются с конца мая до августа. Самка откладывает яйца на нижнюю сторону листьев в один слой, размещая их ближе к краю верхушечной части листа. Кладка состоит обычно из 40 - 60 яиц (с колебаниями от 10 - 12 шт.). Яйца полукруглые с плоским основанием. Нижняя половина яйца темно-зеленая, верхняя светлая с темной точкой в середине. Через 12- 15 дней из яиц выходят светло-желтые с черными точками гусеницы, усаженные волосками. У них черная блестящая большая голова и семь пар ног, а вместо восьмой (последней) пары находятся расположенные горизонтально две черные твердые хитиновые трубки. Гусеницы в первых двух возрастах живут совместно и только скелетируют листья, а начиная с третьего возраста съедают их целиком. К этому времени у них появляется вместо хитиновых трубок восьмая пара ног, а на головной капсуле характерный знак в виде желтоватой вилки. Взрослая гусеница темно-бурая с 10 прерывистыми желтыми продольными полосами и желтыми поперечными перевязями на каждом сегменте. Она покрыта тонкими желтовато-серыми полосками, длина 3 - 3,5 см.
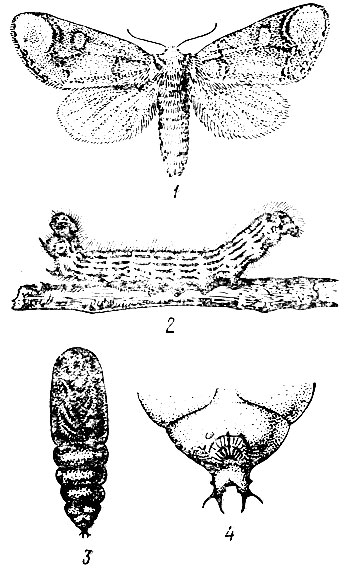
Рис. 73. Лунка серебристая: 1 - бабочка, 2 - гусеница, 3 - куколка (внешний вид), 4 - кремастер
Гусеницы имеют пять возрастов и живут 40 - 50 дней. За это время одна гусеница съедает около 10 - 11 г листвы (20 дубовых листьев средней величины). Гусеницы лунки многоядны, однако предпочитают дуб, затем липу, березу, тополь. При недостатке пищи переходят на деревья других пород. В сентябре гусеницы окукливаются в почве на глубине до 5 см, часть из них углубляется на 10 - 15 см.
Зимует куколка. Она темно-бурая, блестящая, длиной 3 - 5 см. Кремастер оканчивается четырьмя веерообразно расходящимися отростками. Генерация одногодовая. Часть куколок (10 - 15%) обычно перележивает в почве еще год и если они не погибают, то бабочки вылетают только на третий год.
Лунка обитает в байрачных лесах юго-востока, встречается в молодых культурах и лесополосах степной зоны, широко распространена в городских насаждениях различных ландшафтно-географических зон, а также в горных дубовых лесах Кавказа. Она образует преимущественно локальные очаги.
В колебаниях численности лунки большую роль играют энтомофаги. Птицы гусениц не трогают. Затухание вспышки чаще всего является следствием диапаузы, грибных и бактериальных болезней, полиэдроза (Марушина, 1974).
Надзор за размножением лунки проводится по куколкам и несколько осложнен наличием у них диапаузы. Поэтому дополнительно нужно проводить контрольные вылавливания бабочек светоловушками.
Пяденицы (Geometridae)
Бабочки разной величины с широкими крыльями и тонким стройным телом. Гусеницы голые, с двумя парами грудных ног, при передвижении изгибают тело, подтягивая заднюю часть к передней. Куколки без кокона.
Это семейство объединяет очень много видов бабочек, гусеницы которых питаются листвой и хвоей древесных пород. Хвойным породам особенно большой вред наносят сосновая и пихтовая пяденицы, а лиственным - зимняя пяденица, пяденицы-шелкопряды и многие другие.
Сосновая пяденица (Bupalus piniarius L.). Бабочка в размахе крыльев 30 - 40 мм. У самки крылья рыжие, более темной окраски на вершине и по внешнему краю, у самца они черно-бурые, с белыми или желтовато-белыми изменчивой формы пятнами (рис. 74).
Лёт в июне. Самка откладывает яйца рядами до 1 - 32 шт. в каждом на старую хвою, а при массовом размножении и на хвою текущего года. Плодовитость одной самки 80 - 230 яиц. Яйца овальной формы, сдавленные наверху, зеленоватые. Фаза яйца при обычных условиях длится около трех недель, а при температуре 25,5° С - восемь дней.
Вылупившиеся гусеницы зеленого цвета с большой головой. Они выедают в хвоинках продольные желобки, чаще с плоской стороны их. Подросшие гусеницы зазубривают хвоинки по бокам, оставляя нетронутыми оси и основания. Хвоинки покрываются смоляными натеками и засыхают. В старшем возрасте гусеницы съедают хвоинки, оставляя лишь небольшие пеньки.
Гусеницы сначала объедают прошлогоднюю хвою, а в конце августа переходят на питание хвоей текущего года. Питаются они ночью.
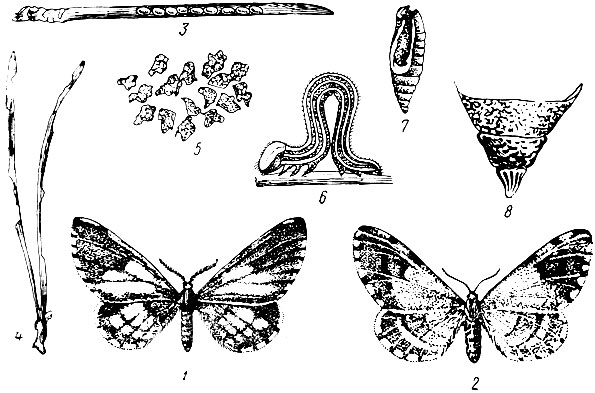
Рис. 74. Сосновая пяденица: 1 - самец, 2 - самка, 3 - яйца на сосновой хвое, 4 - поврежденные хвоинки, 5 - экскременты, 6 - гусеница, 7 - куколка, 8 - кремастер куколки
Взрослые гусеницы зеленые с пятью продольными желтовато-белыми полосками (длина 30 мм), кормятся в кронах до глубокой осени и в октябре спускаются в подстилку, где сразу же окукливаются. Куколка сначала зеленая, а затем становится желтовато-бурой с заостренным грубо морщинистым кремастером, длина 7 - 15 мм. Она зимует. Генерация одногодовая.
Вид мезофильный. Очаги образуются в жердняках и средневозрастных насаждениях различной полноты, в борах-зеленомошниках по ровным и пониженным элементам рельефа, часто даже во влажных условиях травяно-осоковых сосняков.
Комплекс паразитов в отдельных биотопах резко снижает общую численность популяций. Из хищников особенно активны пауки и рыжий лесной муравей, многие птицы. Состав энтомофагов меняется по годам и областям. На яйцах очень часто паразитирует яйцеед трихограмма.
Большая часть наездников и тахины заражают гусениц четвертого-пятого возрастов. Из них только некоторые наездники вызывают гибель гусениц перед окукливанием. Личинки остальных энтомофагов завершают развитие в куколках пяденицы. Наиболее распространены ихневмониды Cratichneumon nigritarius Grav.j Bar ichneumon bilunulattis Grav. и ряд тахин. Куколки поражаются грибными болезнями. Однако в целом энтомофаги и болезни не всегда могут быть причиной затухания вспышек. Последние часто обрываются сильными ранними заморозками, при которых гибнет основная часть популяции гусениц. Вспышка длится около шести лет. Чаще вспышки бывают в Поволжье и прилегающих областях лесостепной зоны, в Западной Сибири и на Южном Урале. Самые северные очаги возникали в Кировской, а самые южные - в Волгоградской областях.
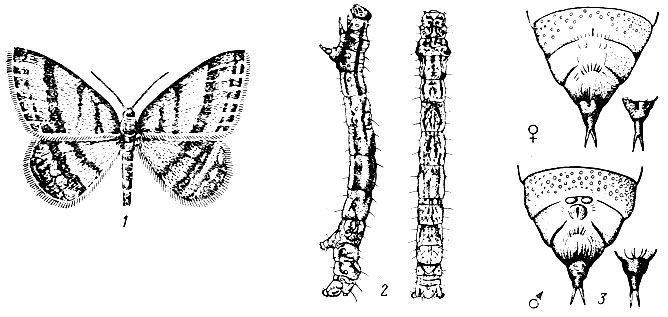
Рис. 75. Пихтовая пяденица: 1 - бабочка, 2 - гусеница, 3 - кремастер куколки
Надзор ведется по калу в сентябре и по куколкам после таяния снега.
Пихтовая пяденица (Boarmia bistortaia Goeze). Бабочка в размахе крыльев 32 - 40 мм. Основной цвет желтовато-серый с темно-бурым рисунком. Передние крылья с поперечной темной полосой, задние с двумя полосами (рис. 75).
Лёт в июне. Самка откладывает яйца кучками по 4 - 10 шт. под отставшую кору сухостойных деревьев, в трещины древесины, реже на растущие деревья пихты, кедра и ели, под отставшую кору сухих сучьев. Плодовитость одной самки около 200 яиц (81 - 331).
Яйца мелкие, гладкие, бледно-зеленого цвета. Через несколько дней приобретают желтоватый оттенок, а перед выходом гусениц - серый. Фаза яйца длится в среднем 10 дней. Вылупившаяся гусеница имеет черный цвет с едва заметными желтоватыми волосками и белыми полосами на первом грудном и брюшном сегментах. По мере роста гусеница принимает окраску от оливкового до грязно-янтарного. Головной щит черный; длина тела 1,7 - 4,2 мм.
Гусеница развивается при температуре 22° С за 30 дней, а при температуре 17,6° С - за 40 дней, пять раз линяет, имеет шесть возрастов. Вылупившиеся гусеницы не откусывают хвоинок, а только пережевывают их, питаясь соком. Гусеницы повреждают только молодую хвою пихты и редко других хвойных пород. Взрослые гусеницы съедают хвою целиком. Всего одна гусеница уничтожает около 1800 мг зеленого корма (370 хвоинок пихты).
Окукливание в августе в лесной подстилке и верхнем слое почвы. Куколка красновато-коричневая, кремастер с длинной развилкой; длина 15 мм. Куколка зимует, генерация одногодовая.
Пихтовая пяденица - полифаг. Гусеницы повреждают в условиях Сибири более ста видов растений. Кроме пихты особенно сильно повреждаются смородина, черемуха, черника, седмичник (Прозоров, 1955). В Европе вредит лиственнице. Вспышки повторяются редко.
Роль энтомофагов и других биологических факторов в динамике численности выяснена недостаточно.
Надзор нужно вести по валу и гусеницам, контрольные учеты - по зимующим куколкам.
Зимняя пяденица (Operophtera brumatq L.). Самец в размахе крыльев 20 - 25 мм, самка с зачаточными крыльями, не летает. Передние крылья у самца желто-серые с тусклыми волнистыми и поперечными полосами, задние крылья беловато-серые. Самка буросерая с длинными ногами (рис. 76).
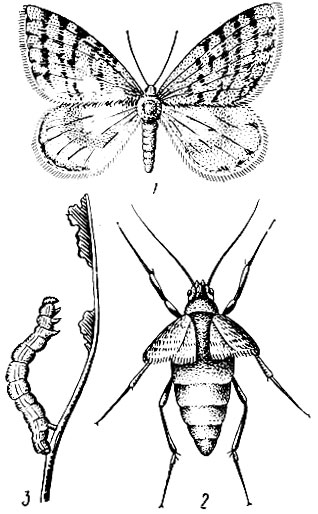
Рис. 76. Зимняя пяденица: 1 - самец, 2 - самка, 3 - гусеница
Лёт начинается в октябре и длится 40 - 50 дней. Наиболее интенсивный лёт наступает через 6 - 10 дней после его начала и продолжается 17 - 19 дней. В Крыму и на Кавказе лёт бывает до января. Наблюдаются случаи, когда единичные куколки перезимовывали и бабочки вылетали только весной.
Максимальная активность бабочек наступает в сумерки, самцы летят на свет. Нормальная жизнедеятельность бабочек возможна лишь при положительной температуре и небольших кратковременных падениях ее ниже нуля. Оптимум в пределах 5 - 12°С. Лёт и спаривание бабочек происходит даже во время дождя. Бабочки гибнут при температуре - 20° С. Самки выдерживают температуру - 9° С в течение недели, но при этом яиц откладывают вдвое меньше и среди них увеличивается число неоплодотворенных. Самки дополнительно не питаются, ротовой аппарат их редуцирован.
После выхода из куколок самки взбираются по деревьям и откладывают яйца около почек и междоузлий, в трещинах и неровностях коры тонких веточек по всему протяжению кроны, а также на коре ствола. Самка откладывает по одному-два яйца или кучками по 10 - 20 шт. Плодовитость самок очень изменчива. В среднем одна самка откладывает 200-300 яиц (колебания - 0 - 550)% часть из которых бывает недоразвита.
Яйца светло-зеленого, фисташкового цвета; через 6 - 10 дней после откладки они становятся оранжевыми. На стадии развития зародышевой полосы у яиц наступает диапауза, длящаяся до весны. Перед вылуплением гусениц яйца становятся серо-фиолетовыми. Они характеризуются большой холодностойкостью и свободно переносят температуру -10° С. Нижний термический порог равен -26° С.
Гусеницы отраждаются в конце апреля - начале мая, одновременно с началом распускания листвы кормовых пород. Первые дни после вылупления они живут под прикрытием почечных чешуек и питаются еще не развернувшимися листочками, затем выгрызают большие отверстия в листьях. При массовом размножении гусеницы оставляют одни только жилки или даже только черешки. Оголенные деревья стоят опутанные паутиной, а гусеницы на паутинах свисают гроздьями.
Гусеницы светло-зеленого цвета, с зеленой головой, темной спинной полосой и тремя белыми линиями по бокам тела; длина да 20 мм. Они многоядны, повреждают около 100 видов растений. Однако не все породы повреждаются в одинаковой степени. В разных географических районах повреждаются разные породы. В лесостепной и степной зонах предпочитают рано распускающиеся формы дуба и ильмовых, в лесной зоне - ильмовые, клен остролистый, березу, ивы, черемуху, ясень; в Приморском крае - граб, клен, дуб и т. д. Повсеместно вредит плодовым деревьям.
Питание гусениц продолжается около месяца, а при неблагоприятных условиях затягивается до 50 дней, за это время они четыре раза линяют и проходят пять возрастов. В конце мая - июне гусеницы окукливаются в почве на глубине до 10 см и частично в лесной подстилке около стволов деревьев, в коконе из части почвы. Фаза куколки длится около четырех месяцев.
Куколка светло-бурая, с Т-образным шипом на заднем конце, длина 5 - 8 мм. Генерация одногодовая.
Зимняя пяденица - вид гигрофильный, ее очаги приурочены к злаковым и грабово-осоковым дубравам на солонцах, пойменным насаждениям с участием дуба и ильмовых, встречаются в парках, запущенных плодовых садах и зарослях с участием дикоплодовых. Она часто встречается вместе с другими видами листогрызущих насекомых, образуя комплексные очаги. Они приурочиваются чаще всего к средневозрастным насаждениям, реже спелым, различно сомкнутым. Несмотря на гигрофильность вида, гусеницы предпочитают опушечные деревья и более освещенные части кроны.
Биотические факторы недостаточно регулируют численность зимней пяденицы. Начало и конец вспышки массового размножения обычно определяются погодными условиями, которые обусловливают синхронность в появлении гусениц и раскрытии почек у ранних форм дуба и вяза, а также влияют непосредственно. Размножению способствуют годы с умеренно влажной и продолжительной осенью, без длительных морозных периодов и нежарким и влажным летом. Вспышка обрывается под влиянием сильных морозов (до -40° С) во второй половине зимы или эпизоотии. В небольших очагах затухание обычно происходит медленно, при участии энтомофагов грибных болезней на фазе куколки, птиц.
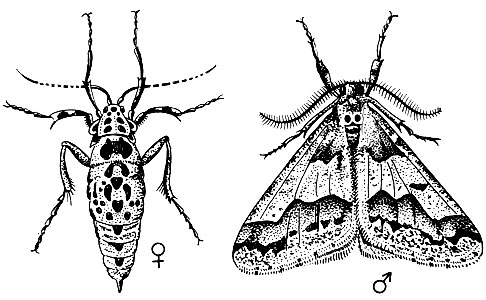
Рис. 77. Пяденица-обдирало (самка и самец)
Рекогносцировочный надзор проводят по бабочкам, используя для этого клеевые кольца и токсические пояса, а для вылавливания самцов - световые ловушки. Детальный надзор - по куколкам в августе.
Пяденица-обдирало (Erannis defoliaria CI.). Самец в размахе крыльев 30-40 мм, самка бескрылая. Передние крылья у самца буро-желтые. Самка темно-желтая (рис. 77). Биологически близка к зимней пяденице. Лёт в сентябре - октябре, зимует в фазе яйца, гусеницы появляются в апреле - мае. Генерация одногодовая. Полифаг. Предпочитает дуб и плодовые. Часто встречается совместно с предыдущим видом.
Надзор нужно вести по бабочкам, используя клеевые кольца и светоловушки и по куколкам в августе.
Пяденицы-шелкопряды объединяют группу видов из семейства пядениц, бабочки которых по внешнему виду напоминают коконопрядов. Общий тон крыльев - серый, брюшко толстое, густо волосистое, усики самцов перистые. Гусеницы крупные от серого до желто-бурого цвета, обладают всеми типичными признаками семейства. Куколки красно-бурые, блестящие, различаются по кремастеру.
В эту группу входят бурополосая (Lycia hirtaria CI.), тополевая (Bistort stratarius Hufn.), фруктовая (Poecilopsis pomonaria Hb.), желтоусая (Apocheima hispidaria Schiff.) и серая волосистая (Phigalia pedaria F.) пяденицы (рис. 78,79).
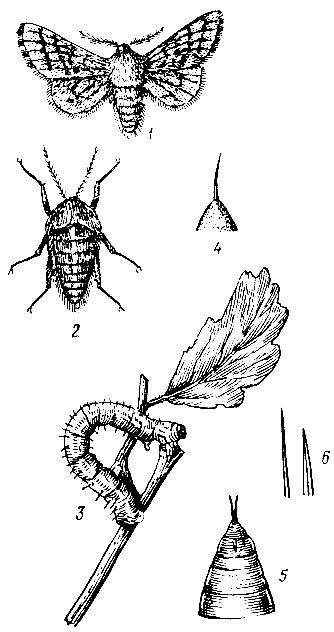
Рис. 78. Желтоусая пяденица-шелкопряд: 1 - самец, 2 - самка, 3 - гусеница, 4 - щетинки на бородавке гусеницы, 5 - кремастер куколки, 6 - щетинки на брюшке

Рис. 79. Бурополосая пяденица: 1 - самец, 2-самка, 3-гусеница, 4 - кремастер куколки
Самые крупные первые два вида. Их гусеницы имеют шесть возрастов и развиваются в течение 50-60 дней. Гусеницы остальных трех видов развиваются 30-33 дня и имеют пять возрастов. У всех видов, кроме бурополосой пяденицы, самки с редуцированными крыльями. Они очень плодовиты, особенно тополевая (до 2000 яиц).
Лёт бабочек происходит ранней весной, часто до вскрытия рек (у желтоусой и серой волосистой пядениц в конце марта, у остальных в апреле). Яйца размещаются самками на скелетных ветвях и кронах деревьев. Фаза яйца длится около 25 дней. Гусеницы повреждают раскрывающиеся почки и молодую листву дуба, вяза, плодовых и других древесных пород. Окукливание происходит в июне - июле в лесной подстилке и почве. Куколки зимуют, генерация у всех видов одногодовая.
Сильные вспышки массового размножения пядениц-шелкопрядов неоднократно наблюдались в Нижнем Поволжье и других местах степной зоны, в Казахстане и Западной Сибири (бурополосая пяденица). Очаги образуются в степных дубравах, пойменных насаждениях с господством вяза и дуба, в заброшенных плодовых садах и березово-осиновых колках.
Затухание очагов во время крупной вспышки на юго-востоке европейской части СССР с господством желтоусой пяденицы (1958 - 1961) происходило под влиянием быстрого развития полиэдроза, вызванного неблагоприятными условиями жизни популяции в последние два года (Воронцов, Голосова, 1968).
Надзор следует вести по куколкам осенью и по калу гусениц весной.
Коконопряды (Lasiocampidae)
Крупные или средней величины бабочки с толстым волосистым телом. Передние крылья больше задних, хоботок редуцирован, бабочки не питаются. Усики гребенчатые, у самцов перистые. Гусеницы с пятью парами брюшных ног, волосистые, куколки в паутинном коконе.
К этому семейству принадлежит ряд видов, гусеницы которых повреждают хвою и листву древесных пород. Не все из них дают большие вспышки массового размножения, часто ограничиваясь небольшим подъемом численности.
Наибольшее значение в лесном хозяйстве имеют сибирский, сосновый и кольчатый коконопряды.
Сосновый коконопряд (Dendrolimus pini L.). Бабочка в размахе крыльев 60 - 80 мм. Окраска крайне изменчива, чаще серовато-коричневая, под цвет сосновой коры. На передних крыльях широкая, неправильной формы, красно-бурая полоса и маленькое белое полулунное пятно. Задние крылья одноцветные (рис. 80).
Лёт с половины июня до конца июля. Самки откладывают яйца кучками на хвою, а при массовом размножении - на ветви и стволы деревьев. В каждой кучке по 20 - 150 шт. Плодовитость одной самки 280-330 яиц. Фаза яйца при температуре 16 - 18° С длится 16 - 20 дней.
Отродившиеся гусеницы серые, с темными бородавками, покрытыми светлыми волосками, но уже после второй линьки приобретают основную окраску. Длина взрослой гусеницы до 75 мм, на втором-третьем сегменте имеются две темно-синие бархатные полосы (их отличительный признак). Зимует в почве на глубине до 10 см.
Весенний выход гусениц начинается после того, как температура почвы на глубине 2 см поднимается выше 10° С и продолжается около двух недель. В этот период гусеницы отличаются чрезвычайной прожорливостью и объедают всю старую хвою. При недостатке старой хвои они объедают и молодую, обгладывают майские побеги и почки. Гусеницы кормятся до середины июня, линяют за это время еще два - четыре раза, а затем окукливаются на ветвях и стволах. За весь период развития гусеницы линяют шесть раз и проходят семь возрастов. Одна гусеница за свою жизнь съедает 20 - 35 г хвои, причем 2 - 3 г в осенний и 18 - 32 г в весенний периоды питания.
Куколка помещается в коконе длиной 30 - 50 мм. Коконы мягкие, удлиненные, буровато-серые. Куколка светло- или темно-бурая, длиной 18 - 40 мм.
Фаза куколки при температуре 16 - 18° С длится четыре-пять недель. В период вспышки массового размножения генерация одногодовая, во время депрессии численности у части популяции затягивается до двух лет за счет вторичной зимовки гусениц.
Гусеницы предпочитают сосну обыкновенную и на других хвойных встречаются очень редко, когда недостает корма. Вид светло- и теплолюбивый, ксерофильный.
Очаги соснового коконопряда возникают в чистых сухих сосняках средней полноты, расположенных на возвышенных местах, в борах-зеленомошниках и в сосновых культурах 12 - 40-летнего возраста разной полноты, на бедных сухих почвах, старопашнях и: песках, изолированных от естественных сосновых лесов.
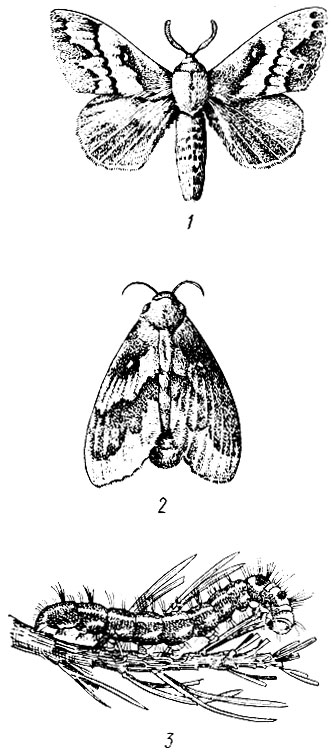
Рис. 80. Сосновый коконопряд: 1 - самец, 2 - самка, 3 - гусеница
В естественных сосновых лестах на всех фазах развития соснового коконопряда паразитирует большой, но медленно действующий комплекс энтомофагов. Главнейшие из них - трихограмма (Trichogramma embryophagum) и теленомус (Telenomus verticillatus) -паразитирует на фазе яйца. В конце вспышки за счет гусениц и куколок развиваются тахины и саркофагины. В изолированных сосновых посадках энтомофаги почти отсутствуют, что также способствует затяжному характеру вспышек в этих условиях.
Вспышки массового размножения длятся шесть - восемь лет. Чаще всего они повторяются в Восточной Украине и ленточных борах Алтая и Казахстана, а также в сосновых лесах лесостепи РСФСР.
Надзор проводится в июне по калу гусениц, осенью по гусеницам в подстилке, весной кольцеванием деревьев в очагах.
Сибирский коконопряд (Dendrolimus slbiricum Tschetw.). Бабочка в размахе крыльев до 80 мм (самка 60-80 мм; самец 40 - 60 мм). Окраска сильно варьирует от светло-желтовато-коричневого или светло-серого до почти черного цвета. Передние крылья пересекаются тремя более темными полосами. В середине каждого крыла находится большое белое пятно. Задние крылья одноцветные (рис. 81).
Ревизия рода Dendrolimus показала, что сибирский шелкопряд является подвидом большого хвойного шелкопряда Dendrolimus superans Butb. Поскольку сибирский шелкопряд может быть признан только подвидом, его эколого-морфологические формы следует считать племенами. Таких племен на территории СССР три: лиственничное, кедровое и уссурийское. Первое - занимает почти весь ареал подвида, два остальных - имеют ограниченное распространение. У лиственничного племени описано пять аберраций.
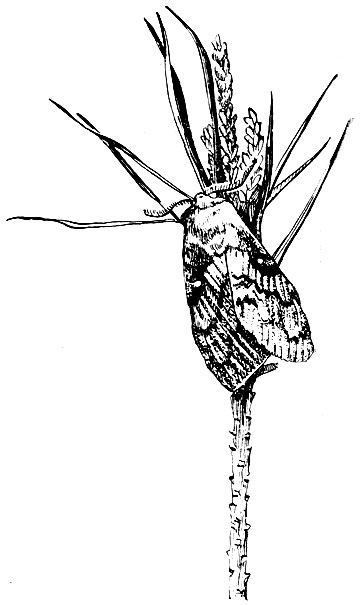
Рис. 81. Сибирский шелкопряд (бабочка)
Массовый лёт во второй декаде июля. На южных склонах гор он происходит раньше, на северных - позднее.
Сразу же после спаривания самки откладывают яйца на хвою, преимущественно в нижней части кроны, а в периоды очень большой численности - всюду: на сухие ветви, лишайники на стволах, подлесок, травяной покров, лесную подстилку. В одной кладке обычно бывает несколько десятков яиц (колебания 1 - 4 до 200 шт.), а всего самка может отложить до 800 яиц, однако чаще всего плодовитость не превышает 200 - 300 яиц.
Яйца почти шаровидной формы в диаметре до 2 мм, сначала голубовато-зеленого цвета с темно-коричневой точкой на одном конце, затем сероватые (рис. 82). Развитие яиц в лесу длится 13 - 15 дней, иногда 20 - 22 дня.
Гусеницы по окраске варьируют от серо-бурого до темно-коричневого цвета; на втором и третьем сегментах тела имеются черные с синеватым отливом поперечные полосы, а на 4 - 12-м сегментах - черные подковообразные пятна; длина тела 55-70 мм. В первом возрасте они объедают только края хвоинок и через 9 - 12 дней линяют. Во втором возрасте они становятся более прожорливыми, через три-четыре недели вновь линяют и в конце сентября опускаются в почву, где, свернувшись кольцом, зимуют под мохом.

Рис. 82. Сибирский шелкопряд: 1 - кладка яиц, 2 - коконы на ветви
В конце апреля гусеницы поднимаются в кроны и начинают питаться, целиком поедая хвоинки, а при недостатке пищи - кору тонких побегов и молодые шишки. Примерно через месяц гусеницы линяют. В второй половине июле - августа они линяют еще раз и осенью уходят на второю зимовку. В мае - июне следующего года взрослые гусеницы интенсивно питаются, причиняя наибольший вред. В этот период они съедают 95% пищи, необходимой для полного развития. Они линяют пять - семь раз и соответственно проходят шесть - восемь возрастов.
Гусеницы питаются хвоей почти всех хвойных пород, произрастающих за Уралом. В июне они окукливаются в буро-сером продолговатом коконе. Куколка вначале светлая, коричнево-красная, затем темно-коричневая, почти черная; длина 25 - 45 мм. Развитие куколки зависит от температуры и длится около месяца.
Цикл развития сибирского коконопряда обычно длится два года, однако на юге ареала развитие заканчивается почти всегда за один год, а на севере и в высокогорных лесах иногда бывает трехгодовая генерация. В образовании одногодовой или двухгодовой генерации решающую роль играет тепло, обусловленное прохождение гусеницами диапаузы. Характерно, что переход на однолетний цикл развития в местах двухгодовой генерацией наблюдается у коконопряда чаще всего в период вспышки массового размножения. Есть указания также, что одногодовой цикл развития наступает в том случае, если сумма температур поднимается до 2100 - 2200° С. При уменьшении суммы тепла до 1800 - 1900° С генерация становится двухгодовой, а при 2000° С - смешанной.
Основные периоды жизни сибирского шелкопряда (лёт, развитие гусениц и т. д.) очень растянуты, а лёт его наблюдается ежегодно, что происходит вследствие существования смешанной генерации. Однако при ярко выраженном двухлетнем цикле развития лётные годы бывают через год.
Шелкопряд повреждает 20 видов древесных пород из рода Pinus, Abies, Picea, Larix и Tsuga, появляется в массе в разные годы и характеризуется изменчивыми формами градационной кривой.
Вспышки массового размножения коконопряда чаще возникают после двух-трех засушливых вегетационных периодов и сопровождающих их сильных весенних и осенних лесных пожаров. В такие годы под влиянием определенным образом складывающегося обмена веществ отрождаются наиболее жизнеспособные и плодовитые особи, благополучно переносящие воздействие внешних условий в критические периоды развития (младшие возрасты гусениц). Лесные пожары способствуют размножению вредителя, сжигая лесную подстилку, в которой гибнут энтомофаги (теленонус). В равнинных лесах вспышкам обычно предшествуют малоснежные суровые зимы, в которые происходит вымерзание энтомофагов, менее холодоустойчивых, чем гусеницы шелкопряда.
Вспышки возникают в первую очередь в изреженных рубками и пожарами лесах, вблизи сырьевых баз, при низкой полноте насаждений разного возраста и состава. Чаще это бывают перестойные и спелые, реже средневозрастные чистые насаждения с редким подлеском и незначительной примесью лиственных пород. В начале вспышки и в периоды депрессии у коконопряда ясно выражена приуроченность к определенным типам леса, формам рельефа, фитоклимату и другим экологическим особенностям насаждений. Так, в равнинной части Западной Сибири очаги образуются обычно в пихтачах-кисличниках и зеленомошниках, а в горной - листвягах-зеленомошниках. В зоне хвойно-широколиствеиных лесов Дальнего Востока очаги связаны со смешанными кедровыми и кедрово-пихтовыми насаждениями, а в Восточной Сибири их размещение тесно связано с особенностями рельефа горных лесов и господством лиственницы и кедра.
По питательности для гусениц на первом месте стоит хвоя лиственницы, затем пихты; хвоя кедра занимает лишь третье место. Поэтому в лиственничных насаждениях наблюдается наибольшая плодовитость бабочек и энергия размножения, а в кедровых - средняя. В пихтачах идет быстрое развитие гусениц по одногодовому циклу, но в ущерб плодовитости, падающей до средних размеров. При питании хвоей ели и сосны происходит быстрое измельчение особей, падение плодовитости и выживаемости.
В колебаниях численности сибирского шелкопряда большую роль играют энтомофаги. Так, в лесах Западной Сибири выявлено 36 видов первичных паразитов, из которых пять видов паразитирует на яйцах, девять - на гусеницах и 22 вида - на куколках шелкопряда.
Главнейший паразит сибирского шелкопряда - Telenomus gracilis, рекомендуемый для борьбы с ним. Он паразитирует на яйцах шелкопряда, переносится бабочками во время их разлета, зимует в лесной подстилке.
Среди возбудителей эпизоотий у сибирского коконопряда ведущее место занимает бактерия Bacillus dendrolimi Tal., описанная Е. В. Талалаевым. Она также используется для борьбы с шелкопрядом.
Вспышки массового размножения длятся 7 - 10 лет, из которых четыре-пять лет насаждениям наносится ощутимый вред. Оголенные гусеницами древостой усыхают и заселяются стволовыми вредителями. Самая неустойчивая порода в тайге - пихта (сибирская, белокорая), наиболее устойчива лиственница (сибирская, даурская, Сукачева).
В первый год сильного повреждения гусеницами деревьев хвойных пород последние заселяются стволовыми вредителями только при полном обесхвоивании. В последующие годы их активная роль и численность быстро нарастают, а еще через два - четыре года вновь начинается резкое падение.
Сибирский коконопряд - страшный враг таежных лесов, и наносимые им убытки столь же велики, как и от лесных пожаров, а сфера вредной деятельности распространяется от Урала до Приморья, захватывает также Монголию, Сахалин, Курильские о-ва, часть Китая, Японии и Северной Кореи.
Надзор за сибирским шелкопрядом нужно сосредоточить в местах его наиболее частых размножений и особоенно тщательно проводить после засушливых периодов, которые благоприятствуют нарастанию численности. Надзор ведется по гусеницам, калу и летающим бабочкам, которые вылавливаются светоловушками. Надзор должен обязательно сопровождаться воздушной разведкой и наземными лесопатологическими обследованиями районов с нарастающей численностью вредителя.
Кольчатый коконопряд (Malacosoma neustria L.), Бабочка в размахе крыльев 32 - 40 мм. Передние крылья охряно-желтые или кирпично-бурые, с двумя поперечными полосами (рис. 83). Лёт в июле. Самка откладывает яйца на побегах, тонких веточках и даже черешках листьев различных деревьев спирально в несколько оборотов. Кладка содержит от 100 до 400 яиц и имеет вид широкого темного кольца, плотно охватывающего побеги, и долго остается на дереве после выхода гусениц.
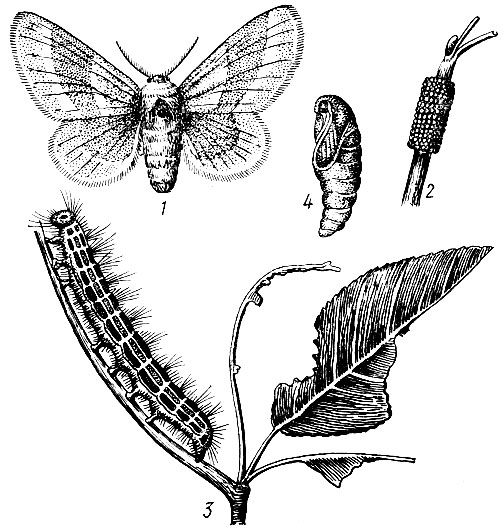
Рис. 83. Кольчатый коконопряд: 1 - бабочка, 2 - кладка яиц, 3 - гусеница и поврежденные листья, 4 - куколка
Яйца зимуют с сформировавшимися в них гусеницами. Обычно зима не оказывает отрицательного влияния на кольчатого коконопряда. Однако гусеницы в яйцах полностью погибают при недельной температуре - 35° С, сильно увеличивается их смертность при - 18° С в течение четырех месяцев.
Гусеницы выходят рано весной (вторая половина апреля, начало мая), за 5 - 17 дней до начала цветения яблони. Гусеницы живут колониями до четвертого возраста, выедают еще нераспустившиеся почки и затем объедают листья с краев, оставляя центральную жилку. Питание гусениц происходит в ночное время; днем они концентрируются в развилках толстых сучьев, где устраивают паутинные гнезда, в которых прячутся в плохую погоду.
Гусеницы голубовато-серого цвета, покрыты довольно густыми тонкими мягкими волосками. Вдоль спины идет белая полоса, на боках несколько желтых полос. Голова серовато-голубая. Молодые гусеницы очень устойчивы к колебаниям температуры и влажности, светолюбивы. Развитие гусениц в природе длится 40 - 45 дней. За это время гусеницы линяют четыре-пять раз и проходят пять-шесть возрастов.
Перед окукливанием гусеницы расползаются, и поэтому коконы никогда не попадаются так скученно, как гусеницы. Окукливание - в июне среди листьев, в трещинах и в листьях, при массовом размножении на травяном покрове. Буро-коричневая куколка находится в двухслойном лимонно-белом коконе средней плотности. Фаза куколки 15 дней. Генерация одногодовая.
Кольчатый коконопряд известен как вредитель плодовых садов, особенно яблони. Однако часто вспышки его массового размножения возникают в парках и дубравах. Гусеницы явно предпочитают дуб, развиваясь на котором дают плодовитое жизнеспособное потомство. Они питаются также тополями, ильмовыми, липой, рябиной, боярышником, терном, шиповником, малиной и т. д.
В комплекс энтомофагов входят главным образом многоядные виды, обычно размножающиеся в нескольких поколениях в год и несинхронные в развитии с кольчатым коконопрядом. Для большинства энтомофагов он является дополнительным хозяином, часто случайным. Ислючение составляет только яйцеед Telenomus laeviusculus Ratz, являющийся специализированным паразитом (Степанова, 1977). Всего за счет кольчатого коконопряда развивается 18 видов яйцеедов (Романова, Лозинский, 1968).
Причиной затухания очагов кольчатого коконопряда чаще всего являются бактериазы в сочетании с неблагопрятной погодой в конце питания гусениц.
Надзор ведется по кладкам яиц в сочетании с наблюдениями за повреждениями насаждений гусеницами и вылавливанием бабочек на светоловушки.
Волнянки (Orgyidae)
Крупные и средней величины волосистые бабочки, похожие на коконопрядов, но глаза голые. Ротовой аппарат редуцирован. Гусеницы с пятью парами брюшных ног, волосистые, покрыты бородавками. Куколки оплетаются паутиной, волосистые.
К этому семейству относится много вредителей древесных пород, дающих вспышки массового размножения на огромных площадях. Из них наиболее известны монашенка, непарный шелкопряд, златогузка, краснохвост, ивовая и античная волнянка, ряд стрельчаток.
Монашенка (Ocneria monacha L.). Бабочка в размахе крыльев 35 - 60 мм. Передние крылья сероватые с четырьмя зигзагообразными черными поперечными линиями: задние крылья беловато-серые. Брюшко розовое, с черными полосками (рис. 84).
Лёт во второй половине июля - августе. Бабочки откладывают обычно 100 - 300 яиц в несколько приемов, максимум по 15-140 яиц каждый раз. Плодовитость и соотношение полов очень изменчивы. Откладка яиц на деревья ели и пихты производится преимущественно в нижней части ствола, а при массовом размножении по всему стволу. В сосновых лесах самки откладывают яйца в самой нижней части стволов (до 0,5 м) в трещины и неровности коры. В чистых сосняках они встречаются чаще на более крупных деревьях, в пониженных местах и на южных склонах. В подлеске и на подросте их можно обнаружить на высоте до 1 м.
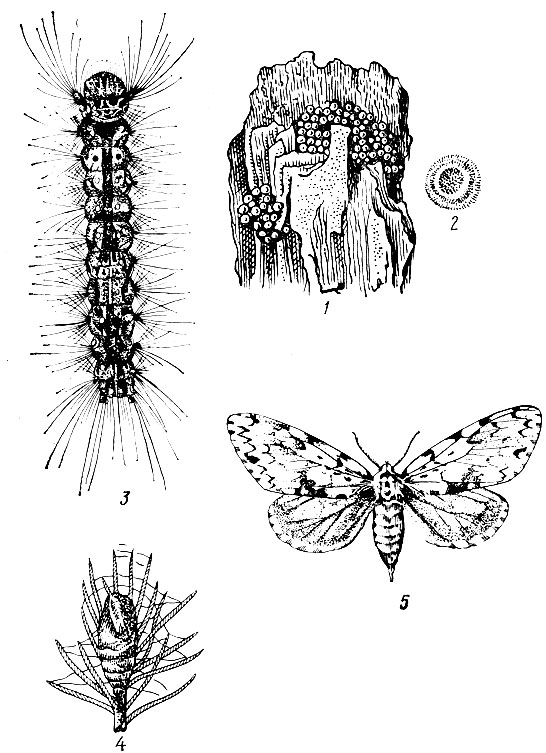
Рис. 84. Монашенка: 1 - кладка яиц на коре, 2 - яйцо (сильно увеличено), 3 - гусеница, 4 - куколка, 5 - бабочка
Яйца имеют шаровидную форму, несколько сдавлены сверху и снизу, величиной несколько больше макового зерна, вначале розовые, затем темно-бурые или серые.
Через четыре недели в яйце развивается гусеница, просвечивающая сквозь оболочку яйца. Она перезимовывает и выходит только следующей весной. Во время нахождения в яйцах зимой гусеницы очень холодостойки и выдерживают низкие температуры - до -30° С. Только что вылупившиеся гусеницы еще несколько дней остаются все вместе, а затем расползаются, опутывая крону нитями. В это время они почти черные, покрыты длинными тонкими волосками и легко переносятся ветром на большие расстояния. Фаза гусеницы длится в зависимости от погодных условий и широты места 45 - 80 дней. Для полного развития гусениц требуется сумма эффективных температур 800° С. Обычно у самок бывает шесть, а у самцов пять возрастов.
Гусеницы монашенки многоядны и питаются хвоей ели, пихты, сосны, лиственницы, листьями дуба, бука, граба, охотно поедают чернику, бруснику и малину. Предпочитаемые породы - сосна и ель. Молодые гусеницы на хвойных деревьях объедают сначала только майские побеги и тронувшиеся в рост почки, затем пыльцу мужских соцветий. Начиная с третьего возраста, гусеницы могут питаться старой хвоей. На лиственных породах гусеницы первого возраста выгрызают набухшие почки, а затем выгрызают дыры в листьях. Гусеницы очень прожорливы. Они откусывают верхнюю половину хвоинки, которая падает на землю, и съедают до конца остальную ее часть.
Окукливание происходит в рыхлой паутине в местах питания гусениц или в щелях коры и на стволах деревьев.
Куколка бронзово-бурая, длиной 15 - 25 мм, с пучком крюкообразных щетинок на заднем конце. Фаза куколки длится 11 - 19 дней. Генерация одногодовая. В западных районах Сибири очаги монашенки возникают в основном в густых (полнота 0,8 - 1) ельниках-зеленомошниках I и II бонитетов, реже в ельниках-долгомошниках или сосняках с хорошо выраженным вторым ярусом из ели. На остальной территории Советского Союза очаги появляются в основных лесах. Это объясняется формовым разнообразием ели: в западной части преобладает рано распускающаяся красношишечная форма ели, а в центральных областях (до Урала) - поздно распускающаяся зеленошишечная форма, которая не может своевременно обеспечить питание гусениц молодой хвоей.
Наиболее частые и крупные вспышки монашенки наблюдаются в лесостепи и прилегающих районах лесной зоны, особенно в сосновых лесах Поволжья, на среднем Урале, ленточных борах Западной Сибири, в БССР.
В лесах этих областей очаги возникают главным образом в глубине массивов сухих и свежих боров, в насаждениях средней густоты II - V классов возраста, очень часто в чистых сосновых культурах II - III классов возраста, произрастающих в относительно пониженных местах рельефа на песчаных и супесчаных почвах.
В колебаниях численности монашенки большую роль играют болезни. Затухание вспышек обычно обусловлено болезнями фляшерией и полиэдразом, развитию которых способствует влажная погода, чаще всего сопровождающая конец вспышки. Влажная погода с обильными осадками в период лёта бабочек делает их малоактивными и препятствует нормальному спариванию и откладке яиц.
Яйца монашенки истребляются поползнями, пищухами и синицами, а гусеницы - кукушкой и иволгой. Комплекс энтомофагов малоэффективен.
Вспышка массового размножения монашенки длится около шести лет. Заметный вред гусеницы приносят два-четыре года. Особенно опасны очаги монашенки в еловых насаждениях. Монашенка - злейший враг ели. После потери хвои ель обычно гибнет, так как отсутствие затенения кронами приводит к быстрому перегреву камбия солнечными лучами и заселению ели короедом-типографом и его спутниками.
Надзор ведется по следующим признакам: в июне-июле - по калу гусениц и огрызкам хвоинок, а со второй половины июля и в августе - по лёту бабочек. В целях надзора используются также ловушки и контрольное кольцевание в сосняках. Учитывается среднее количество яиц на одно дерево. Это служит основным критерием для краткосрочного прогноза.
Непарный шелкопряд (Ocneria dispar L.). Это один из самых распространенных вредителей лесов, садов и зеленых насаждений (рис. 85). Свое название непарный шелкопряд получил за резкое различие между самцом и самкой. Самка в размахе крыльев до. 75 мм, с толстым массивным брюшком, на конце покрытым густыми бурыми волосками. Самец меньше самки (в размахе крыльев до 45 мм), с тонким брюшком и пористыми усиками. Крылья у самки грязновато-белые, с несколькими черными зигзагообразными линиями, а у самца - буровато-серые с широкими прерывистыми темными поперечными полосками и бахромой в темных пятнах по краю. Задние крылья у самца бурые с темным краем и светлой бахромой.
Лёт бабочек непарного шелкопряда происходит в июле. При благоприятных погодных условиях в южных районах европейской части РСФСР он начинается в конце июня - начале июля. В средней полосе массовый лёт бывает во второй половине июля и продолжается до середины августа.
Самки откладывают обычно все яйца сразу, переслаивая их волосками своего брюшка, вследствие чего кладка яиц напоминает кусочек войлока. Кладки бывают разной формы и занимают поверхность в несколько квадратных сантиметров. Свежеотложенные кладки яиц выпуклые и на ощупь упругие; сохранившиеся с прошлого года - серые, пустые, на поверхности их видны маленькие отверстия, оставшиеся после выхода гусениц.
В центральных и северных районах самки откладывают яйца, как правило, в самой нижней части стволов деревьев, не выше 20 - 50 см от поверхности земли. В южных районах кладки можно встретить на всем дереве. В периоды массового размножения самки откладывают яйца всюду: на пнях, валежнике, камнях, постройках, столбах и т. д. Иногда кладки яиц бывают в таком огромном количестве, что, сливаясь, образуют сплошную шерстистую поверхность.
Число яиц в кладке разное. Плодовитость непарного шелкопряда сильно колеблется по годам и географическим районам. В среднем одна самка откладывает 300 - 450, а в отдельных случаях до 1200 яиц.
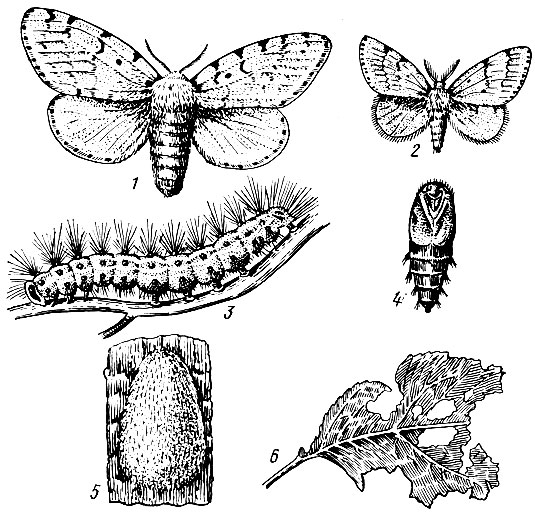
Рис. 85. Непарный шелкопряд: 1 - самка, 2 - самец, 3 - гусеница, 4 - куколка, 5 - кладка яиц, 6 - повреждения
Яйца, будучи покрыты волосками с брюшка самки, устойчивы против воды и морозов (выживают при t до -25 - 30° С).
Формирование гусениц в яйце происходит осенью, после чего они уходят в диапаузу. Для нормального формирования диапаузы необходимо достаточное количество тепла осенью и умеренно-сильные морозы в первую половину зимы. Если гусеницы в яйце уйдут в диапаузу недостаточно окрепшими, последняя бывает неустойчива, и в течение зимы гусеницы в яйцах вымирают. Затяжная осень и теплая зима, прерывающаяся сильными оттепелями, также отрицательно влияют на выживаемость гусениц, усиливают обмен веществ и расход жировых резервов, вызывают их повышенную смертность в яйце, затем, в первом возрасте, усиливают развитие передающейся в поколениях латентной вирусной инфекции, снижают устойчивость к весенним заморозкам.
Выход гусениц из яйца начинается в средней полосе обычно в конце первой декады мая, а на юге несколько раньше: он почти всегда совпадает с наступлением устойчивой теплой погоды (при полуденных температурах выше +10 - 25° С, а среднесуточных - ниже +6° С). Вылупившиеся гусеницы с матово-черной головой и шестью продольными рядами темных бородавок, покрытых длинными тонкими короткими щетинкообразными волосками. Они съедают оболочку яиц и несколько дней сидят кучками, а затем поднимаются по стволам, расползаются по кронам деревьев и начинают поедать листья.
По мере роста гусениц волосистость их становится меньше и окраска бывает серая или серовато-бурая. Характерным и постоянным признаком взрослой гусеницы является на ее теле наличие цветных волосистых бородавок: на первых пяти спинных сегментах - по две синих, на следующих шести - по две красных; по бокам более мелкие, также волосистые красноватые бородавки. Длина взрослой гусеницы колеблется от 45 до 75 мм.
Гусеницы непарного шелкопряда теплолюбивы, предпочитают доступные лучам солнца кроны деревьев, где температура окружающего воздуха достигает 20 - 25° С. При более низких температурах гусеницы плохо питаются, становятся вялыми, а при падении температуры ниже +10° С их развитие полностью прекращается.
Гусеницы, развивающиеся в самцов, линяют четыре раза и проходят пять возрастов; развивающиеся в самок линяют пять раз и имеют шесть возрастов. Для развития гусениц требуется сумма среднесуточных температур около 650 - 700° С. При оптимальных условиях развитие может заканчиваться в 34 - 40 дней, при менее благоприятных - в 50 - 80 дней. Гусеницы становятся взрослыми и готовыми к окукливанию - во второй половине июня.
Непарный шелкопряд - многоядный вредитель. Его гусеницы повреждают более трехсот различных видов растений, в том числе культурные злаки, корнеплоды, клюкву, бруснику, плодовые деревья, большинство кустарников, многие хвойные и почти все лиственные древесные породы. Однако, несмотря на многоядность, гусеницы нормально развиваются и дают плодовитых бабочек только на ограниченном количестве растений. При этом в разных географических зонах набор растений, предпочитаемых непарным шелкопрядом, неодинаков. Наиболее плодовитое и жизнеспособное потомство получается при питании гусениц дубом, тополями, а также плодовыми. Питание кленами вызывает нарушение обмена веществ и резко снижает плодовитость. В лесостепной зоне такое же влияние оказывает питание липой и березой. В северных же районах излюбленными кормовыми породами гусениц являются береза и ива. Гусеницы избегают ясень, белую и желтую акацию, полевой клен, жимолость, бирючину и др.
Гусеницы окукливаются в кронах и на стволах деревьев, в трещинах коры. Они обычно собираются большими группами и оплетают остатки съеденных листьев, образуя своеобразные гнезда.
Куколка матовая, темно-коричневая, почти черного цвета, покрыта ржаво-бурыми волосками. Через две-три недели из куколок выходит новое поколение бабочек; генерация всегда одногодовая.
Непарный шелкопряд - тепло- и светолюбивое насекомое. Он предпочитает хорошо прогреваемые и освещенные чистые насаждения преимущественно порослевого происхождения иди культуры свыше 20-летнего возраста.
В равнинных лесах европейской части СССР первичные очаги возникают в изреженных насаждениях, в рединах и по южным опушкам более густых древостоев, состоящих из ранней формы летнего дуба, березы или граба, а по поймам рек - в насаждениях из вяза и тополя, реже ветлы. Насаждения, в которых возникают первичные очаги, обычно произрастают на бедных почвах (на песках, солонцеватых суглинках, торфяных почвах и т. д.), характеризуются однообразным травяным покровом (осоки, злаки) и плохим ростом. Первичные очаги возникают также в насаждениях, расположенных вблизи населенных пунктов и характеризующихся слабым развитием второго яруса и почвозащитного подлеска, уничтоженных неумеренной пастьбой скота.
В горных лесах Алтая очаги формируются в пихтовых и лиственничных насаждениях. В насаждениях из лиственницы очаги возникают в ряде мест Восточной Сибири и Бурятской АССР. В Средней Азии непарный шелкопряд преобладает в нижней лесной зоне гор и в предгорьях, где образует очаги в плодовых насаждениях.
В Восточном Казахстане очаги непарного шелкопряда образовались в ущельях, хорошо прогреваемых солнцем и защищенных от северных ветров, где гусеницы повреждали пихту, а по поймам ручьев и речек - осину, березу, черемуху, иву.
Частые вспышки массового размножения непарного шелкопряда в значительной степени обусловлены огромным ареалом, большой биологической пластичностью, полифагией и способностью к быстрому расселению гусениц первого возраста.
Рост численности обычно совпадает с господством в течение ряда лет восточной атмосферной циркуляции, которая характеризуется преобладанием антициклональной погоды (жаркое сухое лето, снежная холодная зима без оттепелей).
Вспышки массового размножения наблюдаются ежегодно в тех или иных частях его ареала. Часто они носят местный характер и образуются на небольших площадях, иногда же распространяются на огромных пространствах, захватывая несколько ландшафтно-географических зон. При этом часто формируются в северной части ареала миграционные очаги, возникающие вследствие переноса бабочек циклоном (Воронцов, 1977).
Большую роль в колебаниях численности непарного шелкопряда играют энтомофаги, ускоряющие затухание его очагов. Известно около 150 видов насекомых, паразитирующих на всех фазах развития непарного шелкопряда. Однако размещение их по ареалу вредителя и регулирующее воздействие на популяции неравномерно.
Чаще всего вспышки массового размножения непарного шелкопряда затухают под влиянием полиэдроза. Гусеницы поражаются также бактериальными и протозойными болезнями, часто заражаются нематодами.
В период массового размножения самые различные по биологии лесные птицы переходят на питание доступными и легко отыскиваемыми гусеницами и куколками непарного шелкопряда (например, соловей, синицы, садовая славка, зяблик, мухоловка-белошвейка, кукушка, иволга и др.).
Непарный шелкопряд наносит очень большой вред лесным насаждениям. Общепризнано, что он - одно из важнейших звеньев в общей цепи причин массового усыхания дубрав.
Надзор проводится осенью по отложенным кладкам яиц в насаждениях, наиболее благоприятных для его размножения, а также по бабочкам с помощью аттрактантно-клеевых ловушек с диспарлюром.
Меры борьбы. Даже при незначительной зараженности насаждений (до 0,5 кладки на дерево) в небольших очагах рекомендуется осенняя борьба, заключающаяся в уничтожении кладок яиц.
Кладки яиц уничтожаются путем пропитывания убивающими их веществами или соскабливанием. Для пропитки применяют нефть, жидкий мазут, смесь 2/з керосина и Уз дегтя или мазута, соляровое масло, подсмоленную воду, полученную при сухой перегонке дерева, карболинеум, креозот и дизельное топливо. В керосин для большей эффективности можно добавить технический гексахлоран (2 г на 1 л керосина) или его 12%-ный дуст на 1 л керосина. Можно использовать хлорофос и другие препараты. Хорошее пропитывание кладки яиц достигается двух- или трехкратным нажимом на нее обильно смоченной в керосине кистью.
Соскабливать кладки яиц следует только на молодых стволах деревьев с гладкой корой.
В больших очагах проводятся меры борьбы, указанные для всех хвое- и листогрызущих вредетелей.
Златогузка (Euproctis chrysorrhoea L.). Бабочка в размахе крыльев 30 - 40 мм, снежно-белого цвета с шелковистым блеском. На конце брюшка имеется пучок золотистых (у самки) или бурых (у самца) волосков, откуда и происходит их название (рис. 86).
Лёт в июне - первой половине июля. Самка откладывает яйца на нижнюю сторону листьев кучками продолговатой формы, по 200-500 шт. в каждой, покрывая их золотистым пушком из своего брюшка. Через 15-20 дней (обычно во второй половине июля) из яиц выходят темные волосистые гусеницы, скелетирующие листья дуба и плодовых деревьев, а при массовом размножении - липы, вяза, осины и других пород. Скелетированные листья гусеницы стягивают паутиной, так что получается очень плотное гнездо серого цвета, в котором после второй линьки зимуют в количестве от 200 до 2 000 шт. О количестве содержащихся в гнезде гусениц можно судить по их массе. Средней величины гнездо (около 500 гусениц) весит 4 г. В 1 кг гнезд бывает 180 000 гусениц (Померанцев, 1949).
Весной гусеницы выходят из паутинных гнезд во время раскрывания листовых почек рано распускающейся формы дуба и приступают к питанию почками и молодыми листьями, часто полностью лишая деревья листвы. После выхода из гнезда гусеницы еще линяют два-три раза. Всего у них бывает семь возрастов. Взрослая гусеница серовато-черная, с красными бородавками и белыми пятнами вдоль спины, образующими две красные и две белые продольные полосы; она покрыта желто-бурыми ядовитыми полосками, вызывающими сильное раздражение кожи.
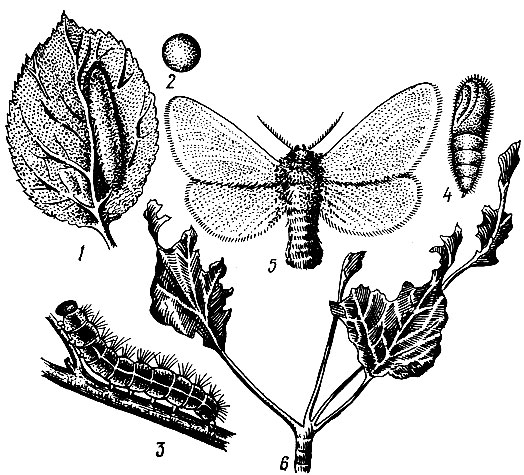
Рис. 86. Златогузка: 1 - кладка яиц, 2 - яйцо, 3 - гусеница, 4 - куколка, 5 - самка, 6 - повреждение
До зимовки питание и развитие гусениц протекает медленно, весной интенсивно и зависит от температуры воздуха. При температуре 20 - 30° С развитие гусениц заканчивается в 30 - 44 дня. Оптимум влажности в пределах 40 - 75%.
Низкие температуры во время зимовки в гнездах гусеницы переносят хорошо, однако холодостойкость их резко меняется в различные периоды жизни. У гусениц в начале зимовки она гораздо выше, чем у гусениц, окончивших зимовку. Холодоустойчивость гусениц зависит также от их питания в летний период. При питании на вторично облиственных деревьях, потерявших весной листву, холодоустойчивость гусениц в пять раз ниже, чем при питании нормальной осенней листвой.
Окукливание гусениц происходит в июне поодиночке или группами в редких паутинных коконах среди недоеденных листьев в кронах и на стволах, а при массовом размножении - и на траве под деревьями. Фаза куколки 15 - 20 дней.
Куколка темно-бурая, с более светлыми волосками, на заднем конце ее имеется вырост с крючками. Генерация одногодовая.
Златогузка отличается исключительно свето- и теплолюбием. Это определяет особенности ее расселения в насаждениях. Гнезда на деревьях распределяются неравномерно. Они приурочены к наиболее освещенным, периферийным частям крон более или менее свободно стоящих деревьев, преобладают на южных опушках, а в сомкнутых насаждениях располагаются на вершинах самых высоких деревьев. На освещенных местах гнезда обычно бывают крупнее, чем в насаждениях.
Первичные очаги возникают в наиболее сухих и прогреваемых солнцем расстроенных дубравах, состоящих из ранней формы летнего дуба, особенно в порослевых несомкнувшихся дубовых молодняках и культурах, в байрачных лесах по балкам и дубовых колках в степях, в запущенных плодовых садах и зарослях дикоплодовых. Очень часто источником распространения злаготузки служат заросли терна и боярышника, представляющие ее резервации в степях.
Несмотря на кажущуюся многоядность, златогузка - типичный вредитель дубрав. Поэтому и вторичные очаги создаются в дубовых культурах и молодняках, но уже сомкнувшихся и более густых, хотя и лишенных почвозащитного подлеска. Они располагаются по юго-восточным склонам балок, иногда в более старых дубравах низкой производительности, ослабленных пастьбой скота, и в поймах степных рек.
Подъем численности златогузки обусловлен определенным устойчивым режимом погоды, наступающим сразу на значительных территориях, и чаще всего совпадает, как и у непарного шелкопряда, с господством восточного типа атмосферной циркуляции. Для развития гусениц златогузки важен не только гигротермический режим весны, но и осенних месяцев.
В колебаниях численности златогузки энтомофаги решающей роли не играют, хотя паразитируют на всех фазах ее развития 87 видов наездников и тахин. Большое значение имеют птицы, особенно большая синица, уничтожающая огромное количество гусениц зимой. Очень часто в очагах бывает расклевано 60 - 70% всех зимних гнезд. Летом гусениц активно уничтожает кукушка.
Затухание вспышки часто происходит под влиянием бактериоза. Его развитие обычно совпадает с дождливой погодой в конце мая - первой половине июня, усиливающей гибель гусениц (Покозий, 1966).
Под влиянием утраты листвы насаждения резко снижают прирост или теряют его совершенно. Одновременно теряется урожай плодов и желудей. При большом скоплении гусениц воздух насыщается их чрезвычайно ядовитыми волосками.
Надзор ведется осенью по гусеницам в гнездах, по количеству и величине гнезд, а также по кладкам яиц и по степени объедания листвы.
Лиственничная волнянка (Dasychira albodentaia Bremer.). Бабочка в размахе крыльев 32 - 50 мм, черно-серая (рис. 87). Лёт в июле. Самка откладывает серого цвета яйца кучками на кору стволов деревьев, реже на сухие веточки и хвою, преимущественно в хорошо прогреваемых местах. Одна самка может отложить 110 - 250 яиц.
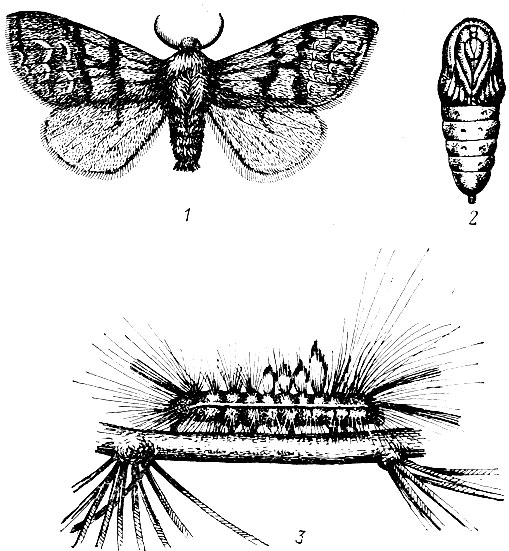
Рис. 87. Лиственничная волнянка: 1 - бабочка, 2 - куколка, 3 - гусеница
Через две недели из яиц выходят гусеницы. Они выгрызают края хвоинок и после третьей (реже второй) линьки уходят в лесную подстилку на зимовку. В начале мая (при температуре в подстилке + 6 - 7° С) гусеницы поднимаются в кроны и продолжают питаться, съедая хвоинки целиком. В этот период они еще три раза линяют и в середине июня окукливаются в кронах деревьев, помещаясь в редком бледно-желтом коконе. Всего гусеницы линяют шесть раз и имеют семь возрастов. Взрослая гусеница почти черная, на четвертом - седьмом сегментах имеются четыре пучка желтовато-белых волосков. На последнем сегменте - желтая волосяная кисточка.
Фаза куколки длится 15 - 18 дней. Куколка коричневая с пучками длинных волосков по бокам и на дорзальной стороне сильно расширена посредине и сужена к концу, длина 23 - 26 мм. Генерация одногодовая.
Волнянка распространена в светлохвойных лесах Восточной Сибири и Дальнего Востока, где образует очаги в сухих, средне- и низкополнотных сосново-лиственничных древостоях.
Кормовые породы - лиственница даурская и сосна обыкновенная, иногда питаются хвоей кедрового стланика и корейского кедра.
Вид теплолюбивый, мезофильный, часто предшествует массовому размножению сибирского шелкопряда или сопутствует ему. Ряд паразитов у обоих видов общий (Telenomus gracilis, Krameria schutzei и др.). Увеличение их численности за счет волнянки в Приамурье задерживает вспышку размножения сибирского шелкопряда.
Античная волнянка (Orgyia antiqua L.). Самец в размахе крыльев 25 - 30 мм, самка с зачаточными крыльями, летать не может. Самец ржаво-бурый, на передних крыльях две явственные поперечные темные полосы, около заднего угла белое пятно. Самка желтовато-серая с хорошо развитыми ногами.
Самки откладывают яйца на поверхности своего кокона в один слой, плотно приклеивая их друг к другу. Яйца шаровидные, приплюснутые, с темной точкой на вдавленной поверхности, белые, затем серовато-желтые. В период зимней диапаузы яйца переносят пониженные температуры до - 40° С. Массовая гибель яиц наблюдалась в Сибири при резких сменах температур в марте - апреле. Для весеннего развития яиц требуется сумма температур 170° С (Прозоров, 1963).
Отрождение гусениц начинается в начале июня. Они имеют пять возрастов. Для их полного развития необходима сумма эффективных температур: для самок 435° С, для самцов 315° С (Прозоров, 1963). В природе длительность развития колеблется в пределах до 45 дней, после чего начинается массовое окукливание. Взрослая гусеница длиной до 40 мм, серая, с тонкими красно-желтыми продольными линиями и красными бородавками. На 11-м сегменте - черная волосяная кисточка.
Окукливаются гусеницы в кроне деревьев и трещинах коры, помещаясь в рыжем коконе. Сумма эффективных температур, необходимая для развития куколок самцов, равна 201С, самок - 132° С. Фаза куколки длится от 7 до 15 дней. Генерация по наблюдениям в Сибири одногодовая (Прозоров, 1963). В лесостепной и степной зонах европейской части СССР генерация двойная. В этом случае лёт бывает в конце июня - июле, гусеницы появляются в июле и окукливаются во второй половине августа. В сентябре летают бабочки второй генерации и откладывают яйца, которые зимуют. Гусеницы появляются с конца апреля и окукливаются в июне.
Античная волнянка - полифаг. В европейской части Союза она предпочитает березу и дуб, но может питаться всеми лиственными породами и многими кустарниками. В Сибири очаги ее массового размножения возникали в лиственничных насаждениях. На других хвойных почти не развивается.
Вспышки массового размножения античной волнянки в лиственничных лесах приурочены к хорошо прогреваемым и освещенным старым древостоям, произрастающим на южных склонах и вершинах гор и сильно изреженным. Длительность вспышки размножения шесть-семь лет. В динамике численности волнянки большую роль играют многочисленные паразиты, из которых особенно активен яйцеед Acholcus dalmanni Rtzb. В Монголии этот паразит был главной причиной затухания очагов античной волнянки и может широко применяться в борьбе с ней.
Надзор весьма труден, так как вся жизнь волнянки проходит в кронах. Лучше всего его проводить на грани двух поколений, наблюдая питание гусениц и их окукливание. В это же время можно обнаружить под деревьями кал. Бабочки-самцы вылавливаются светоловушками.
Краснохвост (Dasychira pudibunda L.). Бабочка в размахе крыльев 35 - 60 мм, светло-серая. На передних крыльях имеются волнистые поперечные полосы темного цвета и бурый налет.
Лёт бабочек во второй половине мая - июне. Самка откладывает серого цвета яйца кучками по 100 - 400 шт. на кору стволов деревьев, преимущественно в нижней части ствола на высоте до 3 м, изредка под кроной. Одна самка может отложить до 1000 яиц.
Через три недели из яиц выходят гусеницы. Они поднимаются в кроны и сначала скелетируют листья, а затем съедают их целиком. Гусеницы имеют шесть-семь возрастов. Причиняют заметный вред с конца июля до половины сентября, а иногда задерживаются на деревьях до октября и питаются уже желтеющими листьями. Они многоядны, но предпочитают дуб, граб, березу, лещины, бук, яблоню. Могут повреждать лиственницу и кустарники.
Взрослая гусеница имеет изменчивую окраску - от лимонно-желтой до серой. Тело густо покрыто волосками; на конце ярко-красная кисточка из волосков, за которую вид получил свое русское название. Длина 4 - 5 мм. Окукливание происходит в лесной подстилке в двухслойном коконе. Куколка зимует. Она темно-бурая с желтоватыми волосками. Булавовидный кремастер с многочисленными крючковатыми щетинками на булаве.
Краснохвост - умеренно гигрофильный вид. Он заселяет разнообразные насаждения. В европейской части СССР первичные очаги возникают главным образом в густых (полнота 0,7 - 1,0) дубовых средневозрастных насаждениях с примесью лещины и березы, расположенных по ровным и пониженным элементам рельефа; преимущественно в типах свежих снытьевых дубрав. Вторичные очаги образуются в более старых и изреженных осоковых дубравах. Вспышки массового размножения носят локальный характер и преимущественно ограничены юго-западной частью Центрального Черноземного района, бывают на Украине. В колебаниях численности большую роль играют многочисленные энтомофаги.
Гусеницы краснохвоста, уничтожая листву во второй половине лета, мало влияют на прирост. Однако при повторных повреждениях и ранних заморозках могут сильно страдать не успевшие подготовиться к зиме побеги.
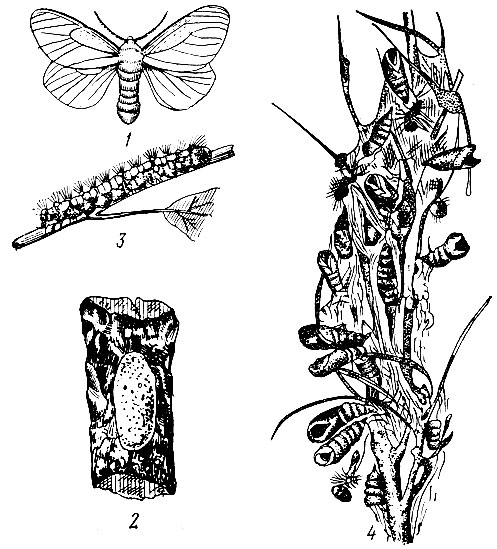
Рис. 88. Ивовая волнянка: 1 - бабочка, 2 - кладка яиц, 3 - гусеница, 4 - скопление куколок в кроне
Надзор ведется по куколкам в сентябре - октябре.
Ивовая волнянка (Leucoma salicis L.). Бабочка в размахе крыльев 40-50 мм. Крылья одноцветные шелковисто-белые, голени, лапки и усики черные (рис. 88).
Лёт в июне - июле. Самка откладывает зеленоватые яйца несколькими кучками на стволы, ветви и листья деревьев, а при массовом размножении в населенных пунктах - на столбы, заборы и стены строений. Кладки яиц, покрытые пенистообразной серебристо-серой пленкой, имеют вид круглых, беловатых, несколько приплюснутых подушечек. Количество яиц в одной кладке колеблется от 5 до 200, плодовитость самки в среднем 500 - 700 яиц, а в благоприятные годы свыше 1000 шт.
Развитие яиц длится при температуре 20 - 25° С 7 - 10 дней. Вылупившиеся из яиц гусеницы скелетируют листья, линяют и во втором возрасте, еще задолго до осеннего понижения температуры, уходят на зимовку в трещины и щели коры, в дупла и под опавшую листву.
Перезимовавшие гусеницы появляются при наступлении устойчивой среднесуточной температуры выше +10° С. Чаще всего это бывает в конце апреля на юге и в первой половине мая в средней полосе. Гусеницы растут быстро и часто линяют. Всего у гусениц шесть возрастов, а у самок бывает семь. Гусеницы питаются только ивоцветными, предпочитая тополя. Среди тополей гусеницы повреждают в первую очередь бальзамический.
В конце июня - июле гусеницы окукливаются на стволах деревьев, среди листьев, на заборах и столбах. Гусеницы собираются перед окукливанием группами по 10 - 25 шт. и оплетают себя паутиной. Куколка блестящая, черная, в белых или желтых пятнышках с пучками седых волосков. В средней полосе ивовая волнянка имеет одну генерацию, на юге - две, а в ряде районов Кавказа - три.
Ивовая волнянка - гигрофильный вид. Она предпочитает густые, хорошо увлажненные насаждения различных возрастов и обильно поливаемые уличные посадки и плантации; дает частые локальные вспышки массового размножения, длящиеся несколько лет. Затухание вспышки происходит сравнительно медленно, обычно под влиянием комплекса энтомофагов и болезней. Последние наиболее сильное влияние оказывают во влажные годы.
Надзор ведется в период питания гусениц и по бабочкам с применением светоловушек и путем обычных наблюдений.
Совки (Noctuidae)
Бабочки различной величины, обычно темной окраски. На внешней стороне передних крыльев характерный рисунок из клиновидных, круглых и почковидных пятен с черточками. Ротовые органы развиты. Усики чаще щетинковидные или нитевидные.
К этому семейству относится много видов, повреждающих листву древесных пород, однако на больших площадях они размножаются редко. В хвойных насаждениях распространена сосновая совка.
Сосновая совка (Panolis flammea Schiff.). Бабочка в размахе крыльев 30 - 35 мм. Передние крылья от серо-бурого до коричнево-красного цвета с поперечными темно-бурыми полосками и белыми крыльями. В середине два пятна. Задние крылья серо-бурые (рис. 89).
Лёт начинается ранней весной с появлением первых проталин и продолжается 10 - 20 дней. Бабочки летают и спариваются по вечерам и ночью в кронах деревьев. Яйца откладывают рядами по 1 - 12 шт. (до 25), в кладке чаще два-три яйца.
Плодовитость одной самки в среднем от 80 до 150 яиц. Яйцо светло-зеленое, позднее темнеет и становится бурым. Фаза яйца 10 - 15 дней, порог развития 62° С, сумма эффективных температур 125° С.
Гусеницы располагаются поодиночке, при случайном контакте выделяют отталкивающий секрет. Только что отродившиеся гусеницы зеленого цвета и похожи на пядениц. В первом - третьем возрастах питаются молодой хвоей, выедают почки, обгладывают побеги, при беспокойстве повисают на паутине. В старших возрастах (IV-V) питаются хвоей прежних лет. В этот период гусеницы становятся светло-зелеными с пятью продольными белыми полосами и оранжевой полосой, проходящей сбоку тела.
Рис. 89. Сосновая совка: 1 - самка, 2 - гусеница, 3 - куколка, 4 - задний конец тела куколки, 5 - экскременты, 6- повреждения майского побега сосны
Развитие гусениц протекает довольно быстро: при температуре 20 - 24° С за 22 - 28 дней. Для полного развития гусениц требуется сумма эффективных температур 460° С. Порог развития гусениц равен 4,7° С. В конце июня - начале июля гусеницы уходят в подстилку и окукливаются через четыре-пять дней. При толщине подстилки до 4 см куколки располагаются на границе между ней и минеральным слоем почвы, при толстом моховом покрове - под слоем мха.
Куколка - красно-коричневая, слабо блестящая. На конце брюшка явственно выступают два шипа, слегка изогнутых вершинами друг к другу, длина 12 - 22 мм. Куколка зимует. Генерация одногодовая.
У сосновой совки много врагов. Большая часть популяции погибает на фазе гусеницы от паразитов и болезней, особенно в последних возрастах от мух-тахин. Гусеницы часто страдают от эпизоотий, которые наряду с паразитами неоднократно были причиной затухания вспышки. Куколки истребляются мышами и землеройками, поражаются микозами.
Размножение совки обычно происходит в крупных лесных массивах. Очаги образуются в нормально сомкнутых чистых сосновых насаждениях жерднякового и среднего возраста, особенно в культурах, расположенных по повышенным элементам рельефа, в борах-беломошниках и зеленомошниках. Повреждения сосновой совки сильно сказываются на приросте и часто ведут к размножению стволовых вредителей и гибели насаждений. Массовые размножения совки известны в разных областях, особенно на Южном Урале и в Тюменской области, в лесах Западной Сибири, Алтайского края, в лесостепной зоне европейской части СССР.
Надзор за размножением сосновой совки лучше всего проводить по куколкам в подстилке, на пробных площадях осенью.
Перепончатокрылые (Hymenoptera)
Тентредообразные пилильщики (Tenthredinidae)
Тентредообразные пилильщики имеют две пары перепончатых крыльев, из которых задние несколько меньше передних. Ротовые органы грызущие; все отделы груди плотно слиты между собой. Брюшко сидячее и его последний сегмент у самки снабжен пиловидным яйцекладом, за что и получили название пилильщиков. Окраска тела различная, темная или яркая.
Взрослые насекомые активны перимущественно в солнечную, лишенную ветра погоду; больших перелетов не делают. Большинство пилильщиков откладывает яйца в листья (хвою), глубоко погружая их в паренхиму, и лишь немногие виды - в зеленые побеги, черешки листьев, бутоны и прямо на листья. Самка пропиливает яйцекладом ткань растения и в образовавшийся кармашек помещает яйцо.
Личинки (ложногусеницы) - 20 - 22-ногие. Тело голое или с редкими короткими волосками; ярко окрашены или имеют покровительственную окраску (под цвет листвы) и тогда мало заметны. Многие виды живут гнездами и при тревоге одновременно быстро поднимают вверх переднюю часть тела, изгибаясь в виде буквы S. Активная жизнь личинок продолжается всего около месяца (4 - 6 недель), но закоконировавшаяся личинка может впадать в диапаузу, которая иногда длится несколько лет.
При нормальном развитии у большинства пилильщиков бывает две генерации в год, встречаются виды, имеющие три поколения в год и строго одногодичный цикл.
Большинство пилильщиков в фазе личинки развивается за счет одной или нескольких кормовых пород (олигофаги), очень немногие виды многоядны.
Личинки окукливаются в кожистых плотных, водонепроницаемых и темноокрашенных коконах. Только у небольшого числа видов они рыхлые, сетчатые или отсутствуют вовсе. Коконы помещаются обычно в земле (на глубине до 20 см) или в кронах деревьев. Находясь в коконах, личинки весьма требовательны к относительной влажности. Кроме листвы личинки ряда видов повреждают плоды и побеги, выедая их сердцевину.
Очаги чаще всего создаются на сравнительно ограниченных площадях и носят пятнистый характер, что связано со слабой лётной способностью самок и гнездовым образом жизни личинок. Они образуются в молодых еще не сомкнувшихся кронами насаждениях и жердняках, реже во взрослых древостоях. Пилильщики очень распространены в полезащитных полосах, лесопарках, в питомниках и в плодово-ягодных насаждениях.
Тентредообразные пилильщики делятся на пять семейств, включающих большое число видов. Наиболее опасные вредители хвойных пород относятся к семействам Diprionidae (Diprion pint, Neodiprion sertifer, Gilpinia pallida и др.) и Tenthredinidae (Pristiphora obietina, P. erichsoni и др.). Лиственным древесным породам вредит много видов, наиболее распространенные из них относятся к семействам Cimbb cidae (Cimbex femoratus и др.), и Tenthredinidae (Cre sus septentrionalis, Macrophya punctum-album, Caliroa cinxia и др.), Argidae (Arge rosae и др.).
Рис. 90. Обыкновенный сосновый пилильщик: 1 - самец, 2 - самка
Обыкновенный сосновый пилильщик (Diprion pini L., рис. 90). Тело овальное, 7-10 мм длиной. Самка всегда больше самца. У самки брюшко желтое, с черными кольцами по середине, голова черная, усики желтые пиловидные; у самца тело почти черное, лишь конец брюшка желтоватый, усики черные перистые.
Развивается обычно в двух поколениях. Лёт первого поколения в начале мая. Самки откладывают яйца в старой хвое по 10 - 15 шт. в каждую хвоинку. Плодовитость одной самки 80 - 150 яиц. Развитие яиц в природе длится около двух недель.
Личинки зеленые с желтоватым оттенком и бурой головой, длина 22 - 26 мм. Над каждой из брюшных ног, за исключением последней пары, имеются пятна в виде черной точки с запятой. По выходе из яиц личинки начинают объедать хвою с боков, оставляя нетронутыми вершинки и срединную жилку. Остатки хвоинок желтеют, при усыхании скручиваются, придавая поврежденным ветвям заметную издали курчавость. Подросшие личинки объедают хвоинки целиком, оставляя только пенечки, иногда обгладывая и самую кору майских побегов. Они держатся гнездами и, уничтожив сообща хвою на одной ветви, переползают на другие.
Развитие личинок длится 4 - 6 недель и зависит от температуры. При 26 - 30° С оно проходит всего 25 дней, а при 15 - 16° С длится до 36 дней. Дающие самок личинки имеют шесть возрастов, самцов - пять.
Личинки первого поколения окукливаются в конце июня - начале июля на ветвях в твердом коконе бочкообразной формы серого цвета. Фаза куколки длится 12 - 15 дней. Появившиеся из куколок взрослые пилильщики второй генерации откладывают яйца в молодую хвою и во второй половине июля - начале августа появляются личинки второго поколения. Они питаются хвоей текущего года до сентября - октября и затем спускаются в лесную подстилку, где окукливаются в бурых коконах и зимуют. В период вспышки массового размножения пилильщика очень часто наблюдается частичная диапауза закоконировавшихся личинок второго поколения.
В лесной зоне у пилильщика бывает одна генерация в год, и развитие всех фаз замедлено. В этом случае в коконах зимуют личинки, и только весной они превращаются в куколок.
Очаги массового размножения пилильщика возникают чаще в насаждениях жерднякового возраста, реже - в еще несомкнувшихся культурах и спелых древостоях (40 - 100 лет) на повышенных частях рельефа с более сухими песчаными или супесчанными почвами по опушкам леса. Вспышка при двойной рации длится всего" три-четыре года и подавляется комплексом энтомофагов, бактериозами и полиэдренным вирусом.
Основным паразитом яиц является яйцеед - Chrysonotomyia ruforum Krausse. Его лёт совпадает с яйцекладкой у пилильщика. Главные паразиты личинок - тахины - Drino inconspicua Meig." Diplostichus janithrix Htg. Они дают два поколения и в своем развитии синхронны с пилильщиком. Паразиты закоконировавшихся личинок - эуфелид - Dahlbomlnus fuscipennis 3ett., ихневмонид - Pleolophus basizonius, Grav.
Надзор ведется по наличию кала во время питания гусениц и по коконам. Раньше, чем организовать надзор, нужно точно знать, сколько генераций имеет пилильщик в данной местности.
Рыжий сосновый пилильщик (Neodiprion sertifer Geoffr). Во взрослой фазе отличается от обыкновенного соснового пилильщика тем, что самка вся красновато-рыжая, а самец черный. У самки тело более узкое, стройное, длиной 7 - 9 мм, среднегрудь и щиток гладкие, блестящие.
Лёт в конце августа - сентябре. Самки откладывают яйца внутрь хвоинок текущего года. Яйца зимуют, и в первой половине мая из них появляются личинки. Личинки грязно-зеленого цвета с узкой светлой полоской вдоль спины и с черной блестящей головой; над брюшными ногами нет пятен в виде точки с запятой.
Личинки предпочитают сосну обыкновенную и реже повреждают другие виды сосен. Живут гнездами и, уничтожив сообща хвою на одной ветви, переползают всей группой на другую. Характер повреждения хвои такой же, как и у обыкновенного пилильщика. При этом наиболее поврежденной бывает опушка насаждения, редины и подрост в "окнах" древостоя. Питание личинок происходит до середины июня. Они имеют пять возрастов. G середины июня личинки начинают покидать деревья и опускаются в лесную подстилку, где коконируются в плотных бочкообразных золотисто-желтых коконах, а незадолго до вылета превращаются в куколку. Генерация одногодовая.
У личинок (эонимф) часто наблюдается частичная диапауза, когда часть популяции превращается во взрослых насекомых и вылетает осенью, а часть популяции может перележать в коконе один-два года, причем небольшой процент пилильщиков, особенно в высокогорных и более северных условиях, может вылетать в мае, а не осенью. Диапауза нарушает закономерное развитие вспышки массового размножения, и кривая численности через один-два года после падения вновь может подняться. Практически эта бывает редко, так как диапаузирующие коконы в огромном количестве истребляются позвоночными животными, энтомофагами и поражаются мюскардинозом. Кроме того, диапауза чаще наблюдается в конце вспышки, в начале же ее диапаузирующих коконов бывает мало.
Рыжий пилильщик очень пластичен и широко распространен на территории СССР. Очаги его массового размножения возникали в сосняках от Западного Казахстана до Кировской и Ленинградской областей.
Очаги рыжего пилильщика возникают в разнообразных насаждениях естественного происхождения и культурах разных возрастов, полнот и типов, начиная с сухих сосняков и кончая сфагновыми болотами. Примесь ели, дуба и других лиственных пород резко снижает численность пилильщика в сосняках. Наибольший вред он приносит молодым культурам до 20-летнего возраста, где обычно выше всего его численность.
Видовой состав энтомофагов такой же, как у обыкновенного соснового пилильщика, но в регуляции численности энтомофаги играют меньшую роль. Это объясняется тем, что большинство активных энтомофагов, особенно паразитирующих на яйцах и молодых личинках, синхронно в своем развитии с обыкновенным, но не синхронно с рыжим пилильщиком. Паразитокомплекс этого вида детально описан в книге Н. Г. Коломийца, А. И. Воронцова, Г. В. Стадницкого (1972).
Рыжий пилильщик дает частые, но неустойчивые и непродолжительные вспышки массового размножения. Вся вспышка протекает за три-четыре года, что достигается за счет сокращения длительности 3 - 4-й фаз массового размножения. Затухание вспышки обычно обусловлено эпизоотией полиэдренной болезни и диапаузой. Вспышки массового размножения у рыжего пилильщика повторяются гораздо чаще, чем у обыкновенного соснового пилильщика.
Ткачи (Pamphilildae)
По внешнему виду ткачи напоминают настоящих пилильщиков. У взрослых насекомых тело очень широкое, брюшко большей частью сплющенное сверху, голова большая, усики многочлениковые. Крылья очень широкие с характерным жилкованием, отличающимся на первый взгляд по очень извилистому, запутанному ходу жилок.
Рис. 91. Звездчатый ткач: 1 - самка, 2 - яйцо на хвое сосны
Личинки без ложноножек на брюшке, но с хорошо развитыми брюшными ногами: 7 - 8-члениковыми усиками и церками на последнем сегменте. Они живут обществами в паутинных гнездах. Ряд видов этого семейства наносит большой вред молодым посадкам хвойных пород, особенно сосне. Наибольшее распространение имеют звездчатый и красноголовый ткачи. За последние годы вспышки массового размножения их участились в ряде районов лесостепной зоны, в ленточных борах Алтая и Сибири.
Звездчатый пилильщик-ткач (Acantholyda stellata Chr.). Голова и грудь черные, с желтовато-белым рисунком, брюшко рыжее, снизу светлее, сверху вдоль середины обычно широко зачернено, ноги рыжие, крылья прозрачные, к вершине слегка сероватые с бурыми жилками. Усики рыжие, с более темной вершиной. Длина тела 11 - 15 мм.
Лёт ткача начинается во второй декаде мая и достигает максимума к концу третьей декады. Отдельные особи встречаются до середины июня. Массовый лёт обычно совпадает с пылением мужских "цветков" сосны и цветением сирени. После того как самки отложат примерно половину всего запаса яиц, они становятся способными к перелетам и образуют новые очаги (Малый, 1979).
Самки откладывают яйца на хвоинки сосен. На одной хвоинке обычно бывает одно - пять яиц. Яйцо имеет форму лодочки с заостренными приподнятыми краями (рис. 91). Оно желтовато-белого цвета, гладкое, блестящее, позднее становится почти коричневым, длина до 3 мм. Плодовитость одной самки изменчива и по данным разных авторов колеблется от 20 до 120 яиц. Средняя плодовитость составляет около 30 яиц. Фаза яйца длится 12 - 13 дней. Из яйца выходит личинка желтого цвета. С самого начала жизни личинка начинает готовить для себя паутинное гнездо. Сначала она обматывает хвоинку, где помещалось яйцо, из которого она вышла, а затем - соседние хвоинки. Хвоя в это время поедается очень экономно. Личинки старших возрастов покидают гнезда и питаются открыто, поедая старую хвою. Личинка живет около месяца и за это время проходит шесть возрастов. В жаркую сухую погоду развитие личинки заканчивается в 16 - 18 дней. В первой половине июля личинки спускаются на почву и уходят на зимовку. Личинки устраивают в почве специальную колыбельку с плотными стенками и там, согнувшись серповидно, покоятся всю зиму в фазе пронимфы. Окукливание происходит весной, в мае. Фаза куколки длится 10 - 14 дней.
Нормальная одногодовая генерация ткача прерывается диапаузой. Ежегодно диапаузирует около одной трети всех личинок, почему создается видимость трехгодовой генерации. Личинки в почве меняют окраску. Образуются две ярко выраженные цветные аберрации: желтая и зеленая. Для правильного прогноза важно с осени определить количество диапаузирующих личинок. У личинок, не впавших в диапаузу, в конце сентября - октябре на голове появляются темные пятна (места будущих глаз имаго), после чего тело личинки постепенно выпрямляется. У личинок диапаузирующих пятна на голове отсутствуют.

Рис. 92. Красноголовый ткач: 1 - кладка яиц, 2 - личинка, 3 - имаго, 4 - гнездо личинки на поврежденном побеге сосны
Звездчатый ткач образует очаги в чистых сосновых культурах на песках в возрасте 7- 30 лет. Изредка встречается в более старых насаждениях. Заселяет культуры разной сомкнутости, преимущественно густые. Период массового размножения длится девять лет, а иногда и дольше. В последнем случае очаги перемещаются по территории лесного массива и возникают не везде одновременно.
Надзор ведется по принимфам в сентябре - октябре или весной, до окукливания.
Красноголовый пилильщик-ткач (Acantholyda erythrocephala L.). Тело сине-стального цвета; длина 10 - 14 мм (рис. 92). Голова /у самок красная, у самцов с желтым передним краем. Лёт проходит дружно и обычно совпадает с цветением дуба черешчатого. В лесостепной зоне он начинается в третьей декаде апреля и заканчивается в начале мая. В Западной Сибири лёт происходит с конца мая. Взрослые насекомые живут 11 - 18 дней. Самки откладывают продолговатые яйца на прошлогоднюю хвою. Средняя плодовитость 40 яиц (Ляшенко, 1971). Фаза яйца длится 16 - 22 дня.
Личинки живут в паутинных гнездах большими группами или поодиночке. Размеры гнезда по мере роса личинок увеличиваются. Они задерживают экскременты и огрызки хвои, которые подсыхают и приобретают ржаво-бурый цвет. Личинки последних возрастов (V - VI) устраивают индивидуальные гнезда в виде паутинных трубочек, по длине равных размерам личинки. Одна личинка съедает от 42 - 123 шт. хвоинок. Кормовая норма по сухой массе в среднем составляет 390 мг (колебания 308 - 581 мг). Усвоение корма достигает всего 36,5% (Ляшенко, 1971). Угроза сплошного объедания в 20-летних сосновых культурах возникает при 120 зимующих пронимфах самок на 1 м2 (Ляшенко, 1974). Личинки развиваются в течение 20 - 25 дней и в конце июля уходят в почву. В августе появляются пронимфы, весной они превращаются в куколок.
Генерация одногодовая. Личинкам присуща диапауза, за счет которой часто удлиняется генерация до двух лет.
Красноголовый ткач заселяет сосновые культуры на песках в возрасте 10 - 30 лет. Изредка встречается в более старых насаждениях, преимущественно ослабленных антропогенным воздействием. Период массового размножения - семь лет. Надзор - по пронимфам.
Литература
Белов А. Н. Размер выборки при учете кладок яиц непарного шелкопряда в дубовых древостоях.- Лесоведение, 1978, № 6.
Бенкевич В. И. К вопросу о корреляционных связях между солнечной активностью, циркуляционными преобразованиями атмосферы и числом областей в европейской части СССР, охваченных массовым появлением непарного шелкопряда.- Уч. зап. Моск. обл. пед. ин-та, т. 273, Зоология, вып. 8. М., 1970.
Болдаруев В. О. Динамика численности сибирского шелкопряда и его паразитов. Улан-Удэ, 1969.
Васильева Т. Г. Зависимость плодовитости хвоегрызуших чешуекрылых oт биохимического состава хвои поврежденных деревьев.- В кн.: Анатомические, гистохимические и биохимические преобразования у лиственницы при повреждении насекомыми. Иркутск, 1972.
Васильева Т. Г. Особенности развития и размножения античной волнянки (Orgyia antiqua) на хвойных породах Восточной Сибири.- В кн.: Хвойные деревья и насекомые-дендрофаги. Иркутск, 1978.
Вержуцкий Б. Н. Пилильщики Прибайкалья. М., 1966.
Вержуцкий Б. Н. Динамика численности и фенология пилильщиков (Hymenoptera, Symphyta) лиственничной консорции.- В кн.: Ритмы природы Сибири и Дальнего Востока. Иркутск, 1975.
Воронцов А. И. Критерии для назначения химической борьбы в лиственных насаждениях.- Сб. работ Моск. лесотехн. ин-та, вып. 15. Вопросы защиты леса. М., 1967.
Воронцов А. И. Материалы по биологии и экологии зеленой дубовой листовертки.- Науч. тр. Моск. лесотехн. ин-та, вып. 65. М., 1974.
Воронцов А. И. Некоторые итоги изучения непарного шелкопряда. - В кн.: Насекомые - вредители лесов Башкирии. Уфа, 1977.
Воронцов А. И., Иерусалимов Е. В., Мозолевская Е. Г. Роль листогрызущих насекомых в лесном биогеоценозе.- Журн. общ. биолог., т. XXVIII, вып. 2. М.,. 1967.
Воронцов А. И., Голосова М. А. Биология желтоусой пяденицы Biston hispidaria Schiff. в Хоперском Госзаповеднике.- Науч. тр. Ленингр. лесотехн. акад.,. № 115. Защита леса, вып. 1. Л., 1968.
Голубев А. В., Семевский Ф. Н. Распределение депрессирующей популяции непарного шелкопряда.- Зоолог, журн., т. 48, вып. 6. М., 1969.
Голубев А. В. Оптимальная система учета яйцекладок рыжего соснового пилильщика.- Науч. тр. Моск. лесотехн. ин-та, вып. 41. М., 1973.
Голубев А. В. Метод установления оптимального размера пробной площади при учете хвое- и листогрызущих вредителей, зимующих или окукливающихся в почве.- Науч. тр. Моск. лесотехн. ин-та, вып. 65. М., 1974.
Гримальский В. И. Устойчивость сосновых насаждений против хвоегрызуших вредителей. М., 1971.
Данилевский А. С, Кузнецов В. Н. Листовертки (Tortricidae).- Фауна СССР. Насекомые чешуекрылые, т. V, вып. 1. Л., 1968.
Егоров Н. Н., Соложеникина Т. Н. Дубовая хохлатка и борьба с ней в Воронежской обл.- Науч. зап. Воронеж. Лесотехн. ин-та, т. XXI. Воронеж, 1960.
Егоров Н. И., Рубцов Н. И., Соложеникина Т. Н. Дубовая листовертка в Воронежской обл.- Зоолог, журн., т. 40, вып. 8. М., 1961.
Ермоленко В. М.- Фауна Украины, т. 10, вып. 2. Киев, 1972; вып. 3, 1975 (на укр. яз.).
Ефремова В. А., Инсаров Г. Э., Семевский Ф. Н. Оптимальная система слежения за плотностью популяции лесных насекомых.- Журн. общ. биолог., т. 38. М., 1977.
Злотин Р. И., Худашова К. С. Роль животных в биологическом круговороте лесостепных экосистем. М., 1974.
Знаменский В. С. Влияние инсектицидов на фауну насекомых дубрав.- Сб. науч. тр. ВНИИЛМ. Защита леса от вредителей и болезней. М., 1973.
Знаменский В. С. Изменение параметров и модели распределения гусениц дубовой листовертки под влиянием биотических факторов.- Науч. тр. Моск. лесотехн. ин-та, вып. 65. М., 1974.
Знаменский В. С, Лямцев Н. И. Влияние плотности популяции на качественные показатели динамики численности непарного шелкопряда.- Сб. науч. тр. ВНИИЛМ. Защита леса от вредителей и болезней. М., 1980.
Ивлиев Л. А., Синчилина Е. М. Хвойная волнянка в лесах Приамурья и ее роль в очагах сибирского шелкопряда.- В кн.: Экология насекомых Приморья и Приамурья. М., 1964.
Идрисова Н. Т. Материалы по биологии и экологии непарного шелкопряда в Башкирии.- В кн.: Насекомые - вредители лесов Башкирии. Уфа, 1977.
Иерусалимов Е. Н. Температурный режим и влажность воздуха в насаждении, поврежденном сосновым шелкопрядом.- Лесоведение, 1973, № 6.
Иерусалимов Е. Н. Компенсационные процессы в насаждении, поврежденном листогрызущими насекомыми.- В кн.: Фитофаги в растительных сообществах. М., 1980.
Ильинский А. И. Сосновый шелкопряд, его жизнь и меры борьбы с ним в лесах (на укр. языке). Киев, 1958.
Ильинский А. И. Непарный шелкопряд и меры борьбы с ним. М., 1959.
Ильинский А. И. (ред.). Надзор, учет и прогноз массовых размножений хвое- и листогрызущих насекомых в лесах СССР. М., 1965.
Исаев А. С, Хлебопрос Р. Г. Анализ динамики численности лесных насекомых на основе принципа стабильности подвижных экологических систем.- Журн. общ. биолог., т. XXXV, № 5. М., 1974.
Исаев А. С, Хлебопрос Р. Г., Кондаков Ю. П. Закономерности динамики численности лесных насекомых.- Лесоведение, 1974, № 3.
Исаев А. С, Ряполов В. Я. Анализ ландшафтно-экологической приуроченности очагов сибирского шелкопряда с применением аэрокосмической съемки.- В кн.: Исследование таежных ландшафтов дистанционными методами. Новосибирск, 1979.
Ионайтис В. П. Биология пилильщика Gilpinia hercyniae (Hymenoptera, Diprionidae) в Литовской ССР.- Зоолог, журн., т. 48, вып. 8. М., 1969.
Киреева И. М. Об изменчивости морфофизиологических особенностей популяции непарного шелкопряда (Porthetria dispar L.). в Нижнем Приднепровье.- Вестн. зоолог., 1975, № 1.
Киреева И. М. Прогнозирование массового размножения непарного шелкопряда.- Лесное хозяйство, 1978, № 4.
Киреева И. М. Морфофизиологическая структура популяции непарного шелкопряда в Нижнем Приднепровье.- Лесоведение, 1979, № 6.
Кириллов В. П. Массовое размножение соснового шелкопряда в ленточных борах Казахстана.- Тр. ин-та зоолог. АН Казахской ССР, т. VIII. Алма-Ата, 1958.
Кожанчиков И. В. Волнянки (Orgyidae).-Фауна СССР. Насекомые чешуекрылые, т. XII. М - Л., 1950.
Коломиец Н. Г. Звездчатый ткач-пилильщик. Новосибирск, 1967.
Коломиец Н. Г., Стадницкий Г. В., Воронцов А. И. Рыжий сосновый пилильщик. Новосибирск, 1972.
Кондаков Ю. П. Непарный шелкопряд (Porthetria dispar L.) в лесах Красноярского края.- В кн.: Защита лесов Сибири от насекомых-вредителей. М., 1963.
Кондаков Ю. П. Закономерности массовых размножений сибирского шелкопряда.- В кн.: Экология популяций лесных животных Сибири. Новосибирск, 1974.
Коников А. С. Приспособления хвоегрызущих насекомых к условиям среды. М., 1966.
Коников А. С. Регуляторы численности лесных насекомых. Новосибирск, 1978.
Крушев Л. Т. Микробиологический метод подавления очагов массового размножения хвое- и листогрызущих чешуекрылых. - Научн. тр. Моск. лесотехн. ин-та, вып. 65. М., 1974.
Крушев Л. Т., Марченко Я. И., Машнина Т. И. Опыт оценки энтомоцидной активности биопрепаратов по отношению к гусеницам соснового шелкопряда и сосновой пяденицы.- В кн.: Лесохозяйственная наука и практика, вып. 24. Минск, 1974.
Крушев Л. Т., Гримальский В. И. Биологическая профилактика массовых размножений хвоегрызущих вредителей сосны.- Лесное хозяйство, 1978, № 1.
Крушев Л. Т., Марченко Я. И. Опыт применения бактериальных препаратов против шелкопряда-монашенки.- Лесное хозяйство, 1981, № 4.
Ляшенко Л. И. Биология красноголового ткача в условиях лесостепной зоны.- Научн. тр. Моск. лесотехн. ин-та, вып. 38. М., 1971.
Малый Л. П. Биологические и экологические особенности звездчатого пилильщика-ткача в Белоруссии.- В кн.: Фауна и экология насекомых Белоруссии. Минск, 1969.
Малышева М.С. Сосновая пяденица и ее энтомофаги в условиях Савальско-го лесничества Воронежской обл. - Энтомолог, обозр., т. XI, вып. 3. Л., 1962.
Мартынова Г. Г. К биологии и экологии сосновой пяденицы в период вспышки массового размножения 1960-1966 гг.- Науч. тр. Моск. лесотехн. ин-та, вып. 26. М., 1969.
Марушина И. Г. Лунка серебристая (Phalera bucephala L.) в дубовых культурах Ростовской обл.- Науч. тр. Моск. лесотехн. ин-та, вып. 38. М., 1971.
Марушина И. Г. Динамика численности лунки серебристой в Волгодонском лесничестве Ростовской области.- Науч. тр. Моск. лесотехн. ин-та, вып. 65. М., 1974.
Мозолевская Е. Г., Голосова М. А. Новые данные о пяденицах-шелкопрядах.- Лесное хозяйство, 1961, № 7.
Мозолевская Е. Г., Новикова Л. К. Лесохозяйственное значение звездчатого ткача-пилильщика в Бузулукском бору.- Сб. работ Моск. лесотехн. ин-та, вып. 26. М, 1969.
Моисеенко Ф. П., Кожевников А. М. Прирост сосновых насаждений, поврежденных пилильщиками.- Лесное хозяйство, 1976, № 2.
Моравская А. С. Биология и некоторые закономерности изменения численности зимней пяденицы в Теллермановском лесу.- Тр. ин-та леса АН СССР, т. XVIII. М., 1960.
Новикова Л. К. К биологии звездчатого ткача-пилильщика.- Сб. работ Моск. лесотехн. ин-та, вып. 26. М., 1969.
Панкевич Т. П. Пилильщики - вредители сельского и лесного хозяйства Белоруссии. Минск, 1981.
Петренко Е. С, Дрянных Н. М. Биоценотические закономерности использования кормовой базы насекомыми-филлофагами в лесах Нижнего Приангарья.- В кн.: Экология питания лесных животных. Новосибирск, 1978.
Петренко Е. С, Корсакова Т. И., Дрянных Н. М. Характеристика процесса сезонного освоения листового аппарата древесных растений комплексом насекомых.- В кн.: Математический анализ компонентов лесных биогеоценозов. Новосибирск, 1979.
Плешанов А. С. Влияние серой лиственничной листовертки на прирост и плодоношение деревьев в очагах ее массового размножения.- В кн.: Анатомические, гистохимические и биохимические преобразования у лиственницы при повреждении насекомыми. Иркутск, 1972.
Покозий И. Т. Роль эпизоотии в динамике численности златогузки на востоке Украины.- Тр. Харьковского с. х. ин-та. Киев, 1966.
Покозий И. Т. Выживаемость златогузки в зависимости от биотических факторов и погоды в лесных насаждениях придонецкой части Украины.- Тр. Харьковского с. х. ин-та, т. 89. Харьков, 1969.
Прозоров С. С. Пихтовая пяденица Boarmia bistortata Goeze как массовый вредитель пихты сибирской. - Тр. Сибирск. лесотехн. ин-та, т. XI, вып. 3. Красноярск, 1955.
Прозоров С. С, Коршунова Л. М., Земкова Р. И. Античная волнянка (Orgyia antiqua L.) - вредитель лиственницы сибирской. - В кн.: Защита лесов Сибири от насекомых-вредителей. М., 1963.
Радкевич В. А. Экология листогрызущих насекомых. Минск, 1980.
Рафес П. М. Биогеоценологические исследования растительноядных лесных насекомых. М., 1980.
Романова Ю. С., Лозинский В. А. Кольчатый шелкопряд и борьба с ним. М, 1968.
Рожков А. С. Сибирский шелкопряд. М., 1963.
Рожков А. С. Массовое размножение сибирского шелкопряда и меры борьбы с ним. М., 1965.
Рожков А. С. (ред.). Вредители лиственницы сибирской. М., 1966.
Семевский Ф. Н. Прогноз в защите леса. М., 1971.
Семевский Ф. Н. Оптимизация поведения гусениц непарного шелкопряда Porthetria dispar L. при их распределении в кроне.- Журн. общ. биолог., т. 32, 1971, № 3.
Семевский Ф. Н. Исследования динамики численности непарного шелкопряда Porthetria dispar L. на низких уровнях плотности популяции.- Энтомолог, обозр., т. 52, вып. 1. Л., 1973.
Семевский Ф. Н., Ефремова В. А., Мишин А. С. Методика учета плотности популяций дубовой зеленой листовертки в кроне дуба.- Лесоведение, 1972, № 2.
Семевский Ф. Н., Семенов С. М. Динамика численности дубовой зеленой листовертки (Tortrix viridana L.) в Московской области.- Зоолог, журн., т. 37. М., 1978.
Степанова Р. К. Анализ смертности кольчатого шелкопряда в период депрессии численности.- В кн.: Насекомые - вредители лесов Башкирии. Уфа, 1977.
Трофимова О. В. Сосновая совка на юго-востоке лесостепи.- Науч. тр. Моск. лесотехн. ин-та, вып. 90. М., 1976.
Трофимова О. В., Трофимов В. Н., Орлинский А. Д. Влияние степени объедания на состояние насаждений, поврежденных сосновой совкой.- Защита леса, 1979, № 4.
Ханисламов М. Г., Латышев Н. К., Яфаева 3. Ш. Условия развития массовых размножений шелкопряда-монашенки в Башкирии.- В кн.: Исследования очагов вредителей леса Башкирии, т. II. Уфа, 1962.
Ханисламов М. Г., Гирфанова Л. Н, и др. Условия формирования резерваций и нарастания численности непарного шелкопряда в Башкирии.- В кн.: Исследования очагов вредителей леса Башкирии, т. П. Уфа, 1962.
Яфаева 3. Ш. Ивовая волнянка в Башкирии.- В кн.: Насекомые - вредители лесов Башкирии. Уфа, 1977.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'