Глава IX. Стволовые вредители и меры борьбы с ними
Общая характеристика
Стволовые вредители составляют большую экологическую группу насекомых, питающихся тканями стволов дерева; в фазе личинки ведут скрытый образ жизни. К ним относятся насекомые преимущественно из отряда жесткокрылых: семейства короеды, усачи, златки, долгоносики и др., а также рогохвосты (отряд перепончатокрылые), древоточцы и стеклянницы (отряд чешуекрылые).
Стволовые вредители обладают разной степенью активности. Одни из них нападают на деревья без видимых признаков ослабления, другие - только на очень ослабленные, почти утратившие свои жизненные функции, или поваленные деревья. В связи с этим еще в прошлом столетии возник спор о способности насекомых этой группы заселять здоровые деревья и о целесообразности названия их "вторичными вредителями". В настоящее время считается общепризнанным, что активность стволовых вредителей зависит от экологических условий, в которых они живут.
Стволовые вредители, относящиеся к различным семействам, сильно отличаются по своему строению и биологии. Поэтому каждому семейству дается отдельная характеристика. Объединяет всех стволовых вредителей сходная экология и прежде всего их взаимоотношения с древесными породами.
Заселение деревьев
Большинство стволовых вредителей - олигофаги и встречаются на нескольких родственных древесных породах. В пределах предпочитаемых древесных пород процесс выбора и заселения деревьев стволовыми вредителями определяется последовательным действием аттрактантов, оказывающих первичное привлечение, и феромонов, вызывающих вторичное привлечение (см. гл. IV). При полете насекомые ориентируются по запаху подходящих для заселения деревьев. Такие деревья обычно усиливают продуцирование аттрактантов в силу изменения их физиологического состояния.
После поселения первых насекомых на деревьях соответствующего физиологического состояния ими начинается усиленное выделение феромонов, что резко увеличивает привлекательность дерева. Эти насекомые получили название "первопоселенцев". Чем их больше, тем сильнее действие выделяемых феромонов и быстрее происходит дальнейшее заселение и освоение дерева вредителями.
Изменение физиологического состояния деревьев обычно связано, с нарушением их водного режима. У хвойных пород при этом уменьшается давление живицы, которая механически и токсически защищает деревья от нападения стволовых вредителей, изменяется давление лубяного слоя коры, а у лиственных пород - количество выделяемого сока. В целом можно сказать, что у ослабленных деревьев происходят глубокие изменения многих физиологических показателей. При этом многие деревья, мобилизуя свои внутренние резервы, восстанавливают нарушенное нормальное состояние и успешно отбивают атаки "первопоселенцев". Так, после низового пожара в сосновом лесу часто можно заметить смоляные воронки и попытки втачивания сосновых лубоедов на коре деревьев. Это следы неудавшейся атаки дерева насекомыми. У лиственных пород часто наблюдается зарастание начальных поселений усачей и златок в результате интенсивного каллюсования.
Заселенные стволовыми вредителями деревья отмирают различно. Это связано с характером ослабления дерева и последовательностью их заселения. Существует два основных типа ослабления деревьев: корневой и вершинный.
Все причины, ослабляющие корневые системы деревьев (низовые пожары, засуха, изменение уровня грунтовых вод, уплотнение почвы, корневая губка, опенок и др.), приводят к усыханию по корневому типу. В этом случае усыхание начинается с нижней части ствола, которая первой заселяется вредителями. Крона в это время часто бывает еще совершенно зеленой и верхняя часть дерева свободна от вредителей. Образуется характерный сухостой с зеленой кроной.
Под влиянием повреждения смоляным раком, хвоелистогрызущими насекомыми, газами и т. д. деревья начинают усыхать в области кроны. Крона может быть уже заселена насекомыми, в то время как нижняя часть дерева еще жизнеспособна. Такой тип ослабления называется вершинным.
Наряду с этими двумя типами ослабления деревьев в очагах стволовых вредителей встречается тип одновременного ослабления всего дерева, когда оно заселяется насекомыми сразу по всему стволу. Наконец, может происходить отмирание отдельных частей дерева в местах повреждений (ожоги, затески, морозобоины, раковые раны и т. д.) и заселение их вредителями. Такой тип получил название местного отмирания.
Экологические группировки стволовых вредителей обусловлены характером отмирания деревьев в очагах и временем их ослабления. В зависимости от времени ослабления для каждого типа отмирания выделяются еще подтипы: весенний и летний. Установленные типы отмирания дали возможность А. И. Ильинскому (1931, 1958) и его ученикам разработать обобщенные схемы формирования экологических группировок стволовых вредителей, облегчающие работы по лесопатологическим обследованиям, надзору и проектированию мер борьбы.
Очаги массового размножения
В случае массового заселения деревьев стволовыми вредителями в лесах образуются очаги. Условно к очагам относятся ослабленные древостои, где имеется более 10% заселенных вредителями деревьев.
В ослабленных насаждениях насекомые находят избыток пищи за счет потерявших жизнеспособность деревьев, на которых они поселяются. В результате происходит быстрый рост популяции стволовых вредителей. По мере роста численности вредителей незаселенных деревьев в насаждении все меньше. Когда все ослабленные деревья заселяются, начинает увеличиваться плотность поселения вредителей на дереве. Увеличение плотности сначала способствует лучшей выживаемости вредителей, а затем ведет к развитию конкуренции между ними, массовому появлению энтомофагов и болезней.
Очаги стволовых вредителей в насаждениях образуются вследствие засухи, зимних морозов, резкого нарушения уровня грунтовых вод или затопления, эрозии, массового повреждения хвое- и листогрызущими насекомыми, позвоночными животными, пожарами, молнией, ветром и снегом, дымом и газами, поражения грибными болезнями в результате нарушения в лесах санитарных правил, изреживания насаждений, уплотнения почвы и повреждения корневых систем при пастьбе скота и т. д.
Каждый очаг в своем развитии проходит несколько фаз. Обычно различают очаги возникающие, действующие и затухающие. Они отличаются по соотношению деревьев различных категорий и состоянию популяций стволовых вредителей.
В возникающих очагах преобладают ослабленные деревья, часть из которых заселяется вредителями. Действующие очаги характеризуются тем, что свежезаселенные вредителями деревья доминируют над отработанными (старый сухостой). В затухающих очагах встречается больше всего отмерших, уже отработанных насекомыми деревьев (или оставшихся пней) и очень мало ослабленных и свежезаселенных.
Очаг может действовать разное число лет. Это зависит от тех причин, под влиянием которых он возник, и от погодных условий. Различают временные очаги, или эпизодические, действующие от одного до нескольких лет, и хронические очаги, действующие много лет. Последние чаще всего приурочены к местам развития грибных заболеваний и древостоям, произрастающим в неблагоприятных условиях.
Очаги резко отличаются друг от друга в зависимости от вызывающих их причин. Даже в одинаковых по лесорастительным условиям и таксационным показателям насаждениях могут образовываться очаги разных видов стволовых вредителей и неодинаково развиваться.
Ниже рассматриваются наиболее часто встречающиеся типы очагов.
Гари. В лесах особенно распространены временные очаги, возникающие под влиянием лесных пожаров.
Заселение гарей стволовыми вредителями зависит от времени возникновения пожара, силы огня и размеров пожара, возраста поврежденных пожаром насаждений, а также от лесорастительных условий. Большое влияние на распространение стволовых вредителей оказывают также санитарное состояние гарей, запас вредителей в окружающих насаждениях и условия погоды.
По времени возникновения пожара гари делятся на весенние (апрель - май), летние (июнь - июль) и осенние (август). Чаще всего встречаются весенние гари, заселяющиеся вредителями в год пожара и представляющие наибольшую опасность в отношении размножения насекомых. Августовские гари обычно не заселяются в год пожара вредителями.
На гарях происходят очень сильные изменения. Деревья, ослабленные огнем, усыхают, повреждаются вредителями и гибнут. Реакция разных древесных пород на повреждения огнем различна. Деревья с толстой корой, глубокой корневой системой, высоко поднятой кроной и небольшой смолистостью меньше страдают от огня. Сосна, лиственница, дуб - огнестойкие породы, ель и пихта сильнее всех страдают от огня, сибирский кедр занимает промежуточное место.
Одним из важнейших признаков, характеризующих состояние устойчивости основных древостоев, поврежденных пожаром, является высота нагара на деревьях. При средней высоте нагара до 2,5 м отпад из древостоя не превышает 25% по запасу, при 2,5- 4,4 м - 50, а при 4,5 - 6,5 м - 70%. Нагар более 6,5 м сопровождается отпадом свыше 70% по запасу. Наиболее надежным признаком является состояние кроны (Галасьева, 1976).
Беглый низовой пожар не опасен для жизни насаждения. Только отдельные деревья теряют свои защитные свойства, становятся нежизнеспособными и заселяются вредителями. На таких гарях через пять лет после пожара суммарный отпад по запасу составит не более 5%, и очаги вредителей не образуются.
Устойчивый низовой пожар в средневозрастных и спелых насаждениях вызывает ожог корневых лап и корневой шейки деревьев, подсушку луба и просмоление водопроводящих сосудов, приводя к нарушению водоснабжения кроны. Смоловыделительная реакция падает прежде всего в нижней части деревьев, и многие из них в первые два-три года заселяются вредителями. На небольших гарях, размером до 5 га, максимум заселения деревьев стволовыми вредителями приходится на первый-второй год после пожара, на больших пожарищах - на третий-четвертый, а иногда и на пятый год. Очередность заселения и длительность пребывания стволовых вредителей на гарях зависят от времени их образования, лесорастительных условий и географического положения.
Влияние пожара на интенсивность отпада деревьев на гарях и их зараженность стволовыми вредителями увеличивается по мере роста захламленности насаждений. При этом большое значение имеют полнота и форма древостоев. Отпад уменьшается с увеличением полноты насаждений. Более низкая температура и высокая влажность воздуха, а также безветрие в полных древостоях ослабляют интенсивность пожара. В редких древостоях, в связи с иными фитоклиматическими условиями, обстановка для развития пожаров более благоприятная. Кроме того, в редких древостоях больше численность самых опасных стволовых вредителей.
Повальный пожар, обжигающий корни, стволы, сучья, приводит к вывалу части деревьев и значительному их обугливанию и поэтому существенного влияния на размножение стволовых вредителей не оказывает.
После притока вредителей к гари начинается обратный процесс - отток их и рассеивание в окружающих насаждениях, в которых возникают куртинные очажки короедов (в хвойных лесах) и златок (в лиственных лесах), а затем начинается повышенный отпад деревьев.
Чтобы предотвратить это явление, следует использовать гарь как ловчую площадь, быстрее и полнее ликвидировать на ней вредителей, не допуская их разлета в окружающие насаждения.
Грибные болезни. Размножение стволовых вредителей тесно связано с очагами грибных болезней. Непосредственной причиной гибели деревьев в очагах корневой губки и опенка обычно бывают стволовые вредители. Отмирание деревьев идет по комлевому типу. В лесной зоне преобладает весенний подтип заселения, доминируют сосновые лубоеды. В лесостепной и степной зонах ослабление больного дерева происходит за счет усиленной транспирации, которую не может удовлетворить работа поврежденной корневой системы. Поэтому чаще возникает летний подтип комлевого заселения, начинающийся с поселения синей сосновой златки и шестизубого короеда-стенографа. Затем к ним присоединяются черный сосновый усач и стволовая смолевка.
В еловых насаждениях, зараженных корневыми гнилями, массовое размножение стволовых вредителей наблюдается лишь в годы засух. Преобладает весенняя подгруппа видов - в основном типограф; ему сопутствуют двойник и гравер. Роль летней подгруппы (пушистый полиграф, еловая смолевка, усачи) зависит от состава, возраста и полноты насаждений и бывает обычно малочисленной. Деревья в еловых насаждениях чаще всего отмирают по стволовому типу, наблюдается переход от комлевого через стволовый к вершинному.
Смоляной рак чаще всего вызывает местное ослабление и отмирание частей и тканей дерева, обусловленный развитием раковой раны в кроне дерева. Стволовые вредители заселяют часть дерева, расположенную выше раковой раны. Вершина отмирает, но дерево продолжает еще долгое время жить. Если раковая рана расположена под кроной, ослабление и отмирание дерева происходит по вершинному типу. На такие деревья первым нападает вершинный короед, быстро достигая очень высокой численности за счет размножения на ветвях, которые, как правило, не убираются во время санитарных рубок. Совместно с ним поселяются малый сосновый лубоед, четырехзубчатый короед, смолевки, усач сосновых вершин и др.
В лиственных насаждениях существует тесная взаимосвязь между распространением ряда сосудистых и раковых заболеваний древесных пород и размножением стволовых вредителей. Так, очаги голландской болезни почти всегда являются и местами массового размножения ильмовых заболонников.
Засухи. После сильных засух возникают очаги узкотелых златок в дубравах, короеда типографа в ельниках, алтайского усача, лиственничной златки в лиственничниках.
На возникновение очагов стволовых вредителей в насаждениях, поврежденных хвое- и листогрызущими вредителями, указывалось ранее (см. гл. VIII). Обычно это бывает в хвойных насаждениях, особенно в местах размножения сибирского шелкопряда. По следам его повреждений идут черные усачи. Наиболее опасен черный пихтовый усач. Поселяясь на деревьях, лишенных хвои, он быстро увеличивает численность. Отрождающиеся имаго летят в соседние насаждения, где в процессе дополнительного питания ослабляют деревья и тем самым подготавливают кормовую базу для следующего поколения.
Антропогенные влияния. Систематическое возобновление насаждения порослью ведет к его вырождению. Любое такое насаждение ослаблено и в нем создаются условия для размножения стволовых вредителей.
Порослевые насаждения оказываются несравненно более зараженными большим дубовым усачом, чем семенные. Неудовлетворительное состояние грабовых насаждений и появление в них очагов грабовой узкотелой златки и грабового заболонника также связано с их порослевым происхождением.
Изреживание насаждений ниже нормальной густоты для данного местообитания, увеличение периметра опушек, чересполосные рубки ведут к увеличению освещенности, нарушению нормальной лесной среды и ослаблению древостоя. В таких древостоях, как правило, возникают очаги размножения многих стволовых вредителей. В лиственных древостоях пионерами являются златки, в еловых - короеды и усачи, в сосновых - синяя сосновая златка, короеды вершинный и стенограф, черный сосновый усач, в лиственничных - продолговатый короед.
Длительные очаги часто возникают также в насаждениях биологически неустойчивых, произрастающих в плохих лесорастительных условиях, или когда тип культур, древесные породы и их смешение не соответствуют этим условиям.
В ослабленных насаждениях стволовые вредители находят избыток пищи за счет потерявших жизнеспособность деревьев, на которых они поселяются. В результате происходит быстрый рост численности. По мере роста численности вредителей незаселенных ослабленных деревьев становится все меньше. Когда они все заселены, начинает увеличиваться плотность поселения вредителей на дереве. При этом сначала численность молодого поколения возрастает, а потом, при высокой и очень высокой плотности заселения дерева, начинает снижаться. В это время уменьшается длина маточных ходов, число отложенных в них яиц, увеличивается смертность личинок. Плотность поселения вредителей на дереве влияет и на эффективность их врагов.
Стволовые вредители имеют очень много врагов из мира насекомых, а также среди птиц и микроорганизмов. Они оказывают большое влияние на численность отдельных видов и часто регулируют ее. По мнению одних исследователей, регуляция численности осуществляется преимущественно хищниками (Харитонова, 1972), по мнению других,- паразитами (Гириц, 1973). Вопрос этот до конца не исследован. Среди хищников в ходах короедов реже других стволовых вредителей преобладают жуки-подкорники. Фауна паразитов довольно разнообразна. Часто встречаются хищные мухи. Короеды бывают заражены нематодами, микозами, редко бактериозами.
Меры борьбы
Борьба со стволовыми вредителями складывается из надзора за их массовым появлением и распространением, выполнения санитарных правил и химических мер борьбы.
Надзор
Надзор организуется во всех предприятиях лесного хозяйства и осуществляется по группам стволовых вредителей. В соответствии с общими принципами специальный надзор проводится в форме рекогносцировочного - в целях обнаружения массового размножения и площади очагов стволовых вредителей, и детального - для оценки динамики численности насекомых и их угрозы насаждениям.
Детальный надзор проводится методами лесопатологического обследования, а в хронических очагах дополняется ежегодными наблюдениями на стационарных пробных площадях, которые закладываются на 10 лет. При детальном надзоре уточняют первопричину ослабления насаждений, определяют их состояние, видовой состав и основные группировки стволовых вредителей, ведут учет показателей их динамики численности. По результатам полученного прогноза назначаются меры борьбы с вредителями.
При детальном надзоре на пробных площадях ведут пересчет деревьев по категориям состояния (здоровые, ослабленные, сильно ослабленные, усыхающие, свежий и старый сухостой), затем берут из числа свежезаселенных деревьев модели и на них определяют методом энтомологического анализа видовой состав стволовых вредителей, плотность популяции и коэффициент размножения для ведущих видов. Здесь же учитывается состояние популяции, наличие энтомофагов и болезней.
На основании данных надзора в сочетании с анализом метеорологических показателей и оценкой санитарного состояния насаждений составляют прогноз и на его основе проектируют мероприятия по борьбе с вредителями. Угроза предстоящего заселения насаждений стволовыми вредителями определяется по соотношению деревьев разных категорий (в основном заселенных к незаселенным, но сильно ослабленным) и коэффициенту размножения.
Санитарные правила
направлены на предупреждение массового размножения вредителей и болезней в лесах путем систематически осуществляемых санитарных рубок леса и поддержания установленного режима - уборки порубочных остатков и окорки древесины.
В лесу нужно вести систематическую уборку мертвого леса и валежа, выбирать свежезаселенные вредителями и усыхающие деревья, при необходимости проводить выборочные и сплошные санитарные рубки, В местах лесозаготовок необходима своевременная очистка мест рубок, вывозка, окорка или химическая защита лесопродукции, окорка или химическая обработка пней хвойных пород.
При назначении участков леса под санитарные рубки следует обращать внимание на то, что излишнее изреживание насаждений ведет к потере устойчивости деревьев и к их гибели. Необходимо стремиться к тому, чтобы после выборочной санитарной рубки полнота насаждения была не меньше 0,7, а в редких древостоях - 0,6. Под сплошные санитарные рубки отводятся участки, где заражено стволовыми вредителями и усыхает более 40% деревьев. Исключение составляют изреженные ранее, уже расстроенные насаждения.
Намечаемые под сплошную санитарную рубку насаждения должны быть предварительно обследованы комиссией (она назначается директором предприятия) и оформлена вся документация согласно требованиям "Санитарных правил в лесах СССР".
Одна из основных задач рубок ухода за лесом - улучшение санитарного состояния леса и повышение устойчивости древостоев против вредителей и болезней. Поэтому при рубках ухода за лесом в первую очередь вырубаются деревья, зараженные стволовыми вредителями, суховершинные, имеющие механические повреждения и предрасположенные к заселению вредителями, физиологически дряхлые.
Планирование рубок главного пользования
При планировании рубок леса нужно стремиться максимально сжимать периметр лесосек, так как по опушкам чаще всего возникают куртинные очаги стволовых вредителей, избегать чересполосных и кулисных рубок, учитывать направление господствующего ветра, в первую очередь разрабатывать горельники, буреломы, ветровалы и другие категории описанных выше очагов стволовых вредителей и древостои, где они могут возникнуть.
Из числа активных мероприятий по борьбе со стволовыми вредителями в лесном хозяйстве применяются: выборка свежезаселенных деревьев, выкладка ловчих деревьев и химическая борьба с насекомыми.
Выборка свежезаселенных деревьев
Выборка свежезаселенных деревьев проводится систематически во всех насаждениях, которым угрожает опасность размножения стволовых вредителей. Ее нельзя отождествлять с обычной выборочной санитарной рубкой, заключающейся в удалении засохших и фаутных деревьев.
На стволах свежезаселенных деревьев, подлежащих вырубке, лесная охрана ставит отметки, которые проверяются специалистами: техником, лесничим или лесопатологом. Деревья, заселенные стволовыми вредителями, распознаются по одному из следующих признаков: наличию буровой муки у основания стволов, по смоляным воронкам и мелким дырочкам - входным отверстиям насекомых - на стволах, по обильным потекам смолы по стволу, вздутиям на стволах, наполненным черноватой или бурой выступающей на поверхность жидкостью (бляхам), насечкам, сделанным усачами для откладки яиц, по изреженной листве, иногда по пожелтевшей кроне, поврежденным корням, темному камбию. В сомнительных случаях необходим тщательный осмотр со вскрытием участка коры, под которым должны находиться ходы насекомых.
Свежезаселенные стволовыми вредителями деревья следует вырубать в то время, когда под корой находятся личинки. Растягивать время вырубки нельзя, так как появившиеся молодые жуки могут при валке и ошкуровке деревьев остаться на земле и расползтись. Срубленные деревья нужно немедленно окаривать или обрабатывать ядохимикатами, что значительно проще и экономически выгоднее.
Выкладка ловчих деревьев
Выкладка целесообразна только при соблюдении санитарных правил и одновременной выборке свежезаселенных деревьев. При выкладке ловчих деревьев необходимо учитывать экологию соответствующих видов стволовых вредителей, географическое расположение насаждений, их лесорастительные условия, санитарное состояние, направление хозяйства, количество короедов. Ловчие деревья должны быть вовремя выложены, вовремя окорены и разработаны, в противном случае они превратятся в рассадник стволовых вредителей.
Число ловчих деревьев в том или другом участке леса должно соответствовать числу деревьев, заселенных стволовыми вредителями в прошлом году. Численность стволовых вредителей определяется на модельных деревьях при обследовании очагов. Модельные деревья берутся из числа свежезаселенных и на них определяются видовой состав стволовых вредителей, а численность ведущих видов - по числу маточных ходов (брачных камер) у короедов и по числу личинок (или их уходов в древесину) у усачей, златок и слоников. При высокой численности вредителей ловчих деревьев должно быть не более общего числа заселенных деревьев, при средней - не более половины, при слабой - не более четверти. В случае невозможности определить численность стволовых вредителей ловчие деревья выкладывают на основании материалов об отпуске прошлогоднего сухостойного (короедного) леса.
Существует несколько способов выкладки ловчих деревьев: путем оставления их на корню, искусственного ослабления или срубки и раскладывания целых деревьев с кроной, хлыстов или сортиментов. Для большей емкости деревья укладываются на подкладки толщиной 15 - 20 см. Ловчие деревья лучше выкладывать группами, а не в разброс по всему насаждению. Начинать выкладку их нужно за месяц до начала лёта короедов: в конце февраля - марте против первого поколения и в июне - июле - против второго.
Окорку ловчих деревьев следует проводить после отрождения основной массы личинок, но целесообразнее заменить химической обработкой 16%-ным концентратом эмульсии гамма-изомера гексахлорана. Обработка ловчих деревьев должна производиться перед началом лёта тех стволовых вредителей, с которыми ведется борьба в данном районе. В южных районах требуется повторная химическая обработка ловчих деревьев через 1,5 - 2 месяца. Если ловчие деревья заранее не обработаны, их можно опрыскивать в период массового окукливания и появления молодых жуков, однако при этом эффективность борьбы снижается.
Химическая борьба
Химическая борьба со стволовыми вредителями заключается в защите стволов ослабленных деревьев в период лёта вредителей, уничтожении их во время дополнительного питания, обработке свежезаселенных и ловчих деревьев, лесопродукции в лесу и на складах.
Химическая защита ослабленных деревьев заключается в обработке их ядохимикатами перед началом и во время лёта главнейших короедов, усачей и златок.
Лучшие результаты достигаются при опрыскивании стволов деревьев 3%-ными рабочими жидкостями 16%-ного концентрата минерально-масляной эмульсии гамма-изомера гексахлорана и 4%-ным раствором технического гексахлорана в дизельном топливе. При химической защите стволов деревьев нужно учитывать характер коры. При обработке толстой и переходной коры целесообразно снижать крепость применяемых рабочих растворов (до 2 - 4%), но увеличивать расход жидкости, так как трещины и щели коры на таких деревьях глубже. Расход жидкости в этом случае должен быть 0,6 - 2 л на 1 м2 поверхности коры. При обработке гладкой коры стволов и ветвей снижают расход рабочего раствора до 0,2 - 0,4 л на 1 м2, так как он непроизводительно стекает с ее поверхности; концентрацию раствора нужно увеличивать до 6-10%.
Успех химической обработки ослабленных деревьев, подвергавшихся нападению стволовых вредителей, в значительной мере зависит от соблюдения сроков защиты. Поэтому очень важно вести систематические фенологические наблюдения и хорошо знать время появления главнейших видов вредителей, встречающихся в районе защищенных объектов. Следует также обращать внимание на то, какую часть ствола дерева каждый из этих видов заселяет. Если заселяется комлевая часть ствола в области толстой коры, можно ограничиться обработкой только этой части. Гораздо сложнее защищать деревья, заселяющиеся вредителями с вершин. Обработка крон этими же препаратами часто дает неполноценный результат, однако возможна. Ее можно проводить для борьбы с ильмовыми заболонниками, большим сосновым лубоедом, черными усачами и др.
Наряду с защитой ослабленных деревьев в ценных насаждениях химическим путем можно уничтожать жуков на свежезаселенных деревьях перед их вылетом и в местах зимовок. Свежезаселенные деревья срубают и вместо их окорки производят химическую обработку таким же образом, как и стоящих на корню ослабленных деревьев. При этом ядохимикат затекает в трещины коры, входные отверстия насекомых и, просачиваясь под кору, уничтожает личинок, куколок и отрождающихся молодых жуков. Сохранившиеся жуки при вылете вступают в контакт с ядом на поверхности коры и тоже погибают.
Борьба в местах зимовки осуществляется путем опрыскивания поверхности почвы, корневых лап и оснований стволов деревьев, где зимуют многие короеды и слоники (расход жидкости 0,25 - 0,5 л на 1 м2).
Изложенные выше химические меры борьбы успешно применяются против скрытостволовых насекомых, проводящих часть своей жизни или всю жизнь под корой деревьев и лишь некоторое время неглубоко в древесине (короеды, златки, часть усачей, слоники). Против живущих в древесине стеклянниц и древесницы въедливой химический метод разработан пока недостаточно. Возможные приемы борьбы излагаются при описании указанных видов вредителей.
Обзор отдельных видов
Жесткокрылые (Coleoptera)
Короеды (Scolytidae)
Короеды образуют сравнительно немногочисленное семейство жуков, жизнь которых тесно связана с деревом. У них короткое тело цилиндрической формы с небольшой головой. Самый большой короед из встречающихся в СССР 300 видов - около 9 мм длиной, а самый маленький - 1 мм. Цвет жуков обычно коричневый, бурый или черный. Тело покрыто прочными кожистыми надкрыльями, под которыми имеются хорошо развитые перепончатые крылья, с помощью которых жуки летают. Яйца короедов белые, мелкие. Личинки мясистые, безногие, слегка изогнутые, с хорошо заметной темной головой, голые или слабоволосистые. Куколки белого цвета.
По внешним признакам короеды делятся на три резко отличающиеся между собой группы: лубоедов, заболонников и настоящих короедов (рис. 93).
У лубоедов задний конец тела выпуклый и закругленный, как это свойственно большинству других жуков.
Заболонники отличаются формой брюшка, косо срезанного от задних ног к вершине надкрылий, благодаря чему задний конец тела напоминает долото.
Настоящие короеды имеют на заднем конце тела глубокую впадину (тачку), окруженную зубцами и образующую подобие тачки или корзины. Число зубцов и форма их у разных видов различны, но для каждого вида постоянны.
Особенности строения тела короедов тесно связаны с их образом жизни. Почти всю жизнь короеды проводят под корой стволов и ветвей деревьев. Там они прокладывают ходы, имеющие формы определенных фигур. У каждого вида короеда имеется ход определенной формы.
Ходы бывают простые и сложные. Простые ходы состоят из одного канала, который прогрызается самкой и называется маточным ходом. Они бывают продольные и поперечные. Сложные ходы имеют несколько каналов и делятся на звездчатые с уклоном к продольному и поперечному направлениям и лучистые (рис. 94).
Каждый вид короеда всегда поселяется на определенной древесной породе или нескольких близких породах и занимает определенную часть дерева. Так, например, большой сосновый лубоед обычно заселяет дерево в нижней его части, там, где кора толстая, и очень редко встречается на вершине сосны, где кора тоньше, а малый сосновый лубоед, наоборот, заселяет дерево с тонкой корой и не селится под толстой.
Существует связь между формой ходов, местом поселения короедов на хвойных деревьях и строением смолоносной системы. В нижней части ствола, где меньше смолоходов, поселяются короеды, делающие продольные ходы, а в верхней части ствола, где смолоходов больше, встречаются короеды, прокладывающие поперечные и звездчатые ходы.
Лёт короедов обычно начинается весной и длится до середины лета. Первыми (в конце апреля) начинают летать сосновые лубоеды и другие короеды, живущие на сосне. Затем появляются еловые короеды и заболонники, живущие на лиственных породах.
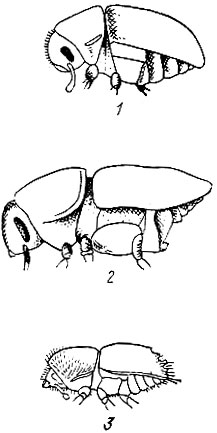
Рис. 93. Представители семейства короедов: 1 - лубоед, 2 - заболонник, 3 - настоящий короед
Короеды в период размножения создают семью. При этом одни виды короедов, обычно лубоеды и заболонники, имеют однобрачную (моногамную) семью, состоящую из самки и самца, а другие, преимущественно собственно короеды,- многобрачную (полигамную) семью, состоящую из одного самца и нескольких самок.
У однобрачных короедов самка прогрызает на дереве продолговатое входное отверстие и прокладывает под корой продольный или поперечный маточный ход. По обе стороны хода самка откладывает яйца в специально устроенные яйцевые камеры, заклеивая их небольшим количеством очень мелких опилок, уплотненных и склеенных выделениями из придаточных половых желез. Вылупившиеся из яиц личинки грызут личинковые ходы. Они постепенно расширяются по мере роста личинок и заканчиваются куколочными колыбельками, в которых личинки превращаются в куколку, а куколки - во взрослых жуков. Жуки отрождаются почти белыми, но постепенно приобретают свой нормальный цвет, прогрызают вылетные круглые отверстия и вылетают для дополнительного питания или на зимовку.
В многобрачной семье входное отверстие прогрызает самец. Под корой он устраивает неправильной формы полость - брачную камеру, в которой могут поместиться несколько жуков. В камеру последовательно проникают самки (от 2 до 12). После спаривания каждая самка начинает прогрызать свой маточный ход и откладывать яйца. Маточные ходы отходят от брачной камеры в разных направлениях.
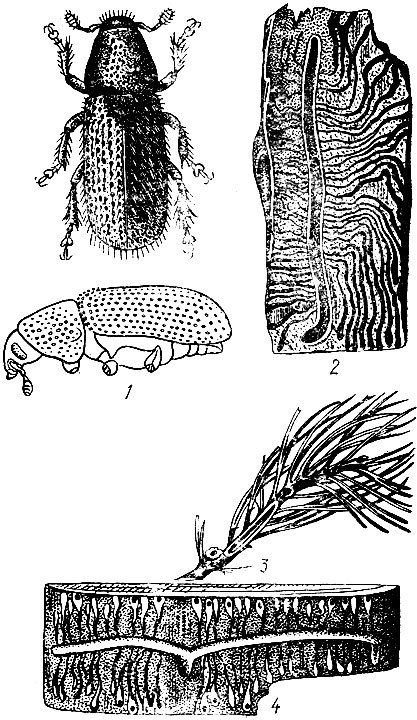
Рис. 94. Типы короедных ходов: 1 - простой прямой продольный ход, 2 - сложный продольный ход, 3 - звездчатый ход с уклоном к продольному направлению, 4 - простой поперечный ход, 5 - сложный поперечный ход, 6 - звездчатый ход с уклоном к поперечному направлению, 7 - лучистый ход, 8 - ход крифала, 9 - семейный ход, 10-12 - лестничные ходы в древесине
Если маточный ход направлен вверх по стволу дерева и находится против входного отверстия, буровая мука (опилки) высыпается через брачную камеру и это отверстие наружу. Из маточных ходов, направленных вниз или расположенных под углом, буровая мука сама высыпаться не может. Ее выбрасывает из хода самец с помощью тачки, расположенной на конце тела.
Среди короедов встречаются такие виды, которые прогрызают входное отверстие, заканчивающееся расширенным коротким маточным ходом, где самка откладывает яйца сразу одной или несколькими кучками (лубоед дендроктон). Отродившиеся личинки грызут совместный семейный личинковый ход, представляющий собой обширную, обычно заполненную смолой полость. Некоторые короеды откладывают яйца кучкой в конце маточного хода, но личинки грызут отдельные ходы в разные стороны (короеды крифалы). Главнейшие типы короедных ходов показаны на рис. 94.
Самые мелкие короеды (длина тела 1,2 мм) рода Crypturgus забираются в ходы других короедов и от их краев начинают прокладывать собственные, образующие густую сеть, ходы.
Короеды, обитающие в древесине (древесинники), имеют свои особенности. Самка обычно прогрызает маточный канал перпендикулярно оси ствола дерева. От маточного канала берут начало кормовые каналы, от которых у одних видов начинаются личиночные ходы, у других - личинки самостоятельных ходов не делают и пользуются только теми, которые сделаны самкой.
У древесинников имеется тесная связь с грибами, которые постоянно находятся в кишечнике жуков и с ними попадают в древесину. В древесине самки перед кладкой яиц производят "посев гриба". Освобожденные споры гриба попадают в благоприятные условия влажной древесины, тут же прорастают и образуют грибницу. Личинки питаются грибницей и за счет этого полностью обеспечены азотистыми веществами, что позволяет им не затрачивать энергии на проделывание длинных ходов в древесине.
Лёт и откладка яиц у короедов длятся около месяца. Фаза яйца продолжается 10 - 14 дней, фаза личинки - 15 - 20 дней, куколки - 10 - 14 дней. Таким образом, весь жизненный цикл завершается в 1,5 - 2 месяца, после чего наступает период дополнительного питания, которое необходимо для полного развития половой системы.
Дополнительное питание у большинства короедов проходит под корой дерева, где они выгрызают короткие ходы разнообразной формы, получившие название минных. Некоторые виды, например сосновые лубоеды, выгрызают внутреннюю часть молодых побегов, которые, не выдерживая своей тяжести, обламываются и падают на землю. Ряд заболонников питается сочным лубом в развилках веточек, а корнежилов - на стволиках молодых сосен.
Холодостойкость короедов тесно связана с характером их зимовки. Те из них, у которых всегда бывает одна генерация (сосновые и ясеневые лубоеды), зимуют в фазе жука у основания стволов, деревьях в коротких минных ходах в толще коры или в лесной подстилке. Благодаря снежному покрову они мало уязвимы во время низких температур. Молодые жуки, куколки и личинки, зимующие под корой деревьев в своих гнездах, различно относятся к низким температурам. Если такая зимовка обычна для вида, личинки выдерживают температуру до -30° С, если необычна, то многие погибают уже при -15° С.
Высокие летние температуры также могут быть причиной большой смертности личинок и куколок короедов. Часто под влиянием солнечных лучей в июне температура под корой ели поднимается выше верхнего теплового порога развития (обычно выше 40° С) короедов, и личинки их в массе погибают.
Короеды обладают различной степенью активности, однако, как правило, совершенно здоровые деревья не заселяют. Многие из них, кроме того, тонко реагируют на окружающие условия освещения, температуры и влажности, являясь типичными индикаторами определенных местообитаний.
В разных географических районах число видов короедов и их активность различны. Особенно разнообразна фауна короедов Кавказа и Дальнего Востока. Много эндемичных видов встречается в Средней Азии.
Меры борьбы с короедами, общие для группы стволовых вредителей, описаны выше.
Наибольшее распространение и хозяйственное значение имеют следующие виды.
На сосне.Большой сосновый лубоед (Blastophagus piniperda L.) (рис. 95). Жук длиной 3,5 - 4,8 мм, продолговатый, черно-бурый, блестящий, надкрылья пунктированы и на покатой части имеют две слабо углубленные бороздки. Летает в конце апреля - мае и первым заселяет ослабленные деревья в сосновых насаждениях разных возрастов, особенно на гарях и в очагах корневой губки. Самки протачивают под толстой корой нижней части сосен снизу вверх продольный одиночный маточный ход длиной от 3 до 23 см без брачной камеры. Ходы отпечатываются на заболони, и края их сильно засмолены. Личиночные ходы длинные, извивающиеся. Отрождающиеся в июне - июле молодые жуки выгрызают вылетные отверстия и улетают в кроны соседних деревьев, где вгрызаются в побеги текущего, реже прошлого года, и выедают сердцевину, в результате чего побеги обламываются. Один жук может повредить до семи побегов. Осенью жуки покидают крону и зимуют у основания стволов сосен, проделывая в толще коры короткие ходы. Генерация одногодовая.
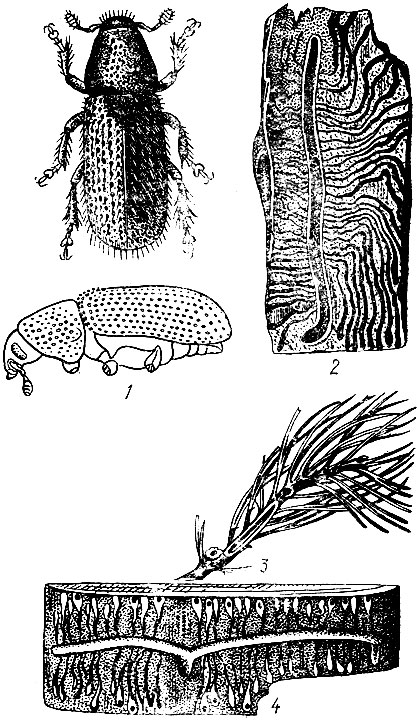
Рис. 95. Сосновые лубоеды: 1 - жук большого соснового лубоеда, 2 - ходы под корой, 3 - побег, поврежденный жуком при дополнительном питании, 4 - ходы малого соснового лубоеда
Малый сосновый лубоед (Blastophagus minor Hart.). Жук длиной 3,4 - 4,0 мм. Очень похож на предыдущий вид, но надкрылья обычно имеют красновато-бурый цвет и на их покатой части углубленные бороздки отсутствуют. Жуки летают на одну-две недели позднее, чем у большого лубоеда. Самки протачивают под тонкой корой в верхней части сосен поперечные, глубоко отпечатывающиеся на заболони маточные ходы, имеющие вид скобки. Длина маточного хода от 4 до 32 см. Короткие личиночные ходы направляются в обе стороны от маточного вдоль ствола дерева. Каждый из них заканчивается колыбелькой глубоко в заболони. Молодые жуки проходят дополнительное питание в кронах, выгрызая побеги, и зимуют в лесной подстилке. Генерация одногодовая.
Оба вида сосновых лубоедов тенелюбивы, встречаются повсеместно в самых разнообразных древостоях. Большой лубоед доминирует над малым в более влажных типах леса. Оба хорошо привлекаются на ловчие деревья.
На Дальнем Востоке кедру корейскому вредит широко распространенный дальневосточный кедровый лубоед (Blastophagus pilifer Spess.), напоминающий по образу жизни малого соснового лубоеда. Там же в горных лесах на саянской ели поселяется еще один представитель этого рода - дальневосточный еловый лубоед (Blastophagus puellus Rt.).
Оба вида имеют годовую генерацию. Большого экономического значения они не имеют, чем отличаются от европейских видов, наносящих большой вред.
Шестизубый короед (Ips sexdentatus Воеrn.) (рис. 96). Жук длиной 5 - 8, чаще 6 мм, коричневый, блестящий. На конце надкрылий тачка, на каждой стороне ее по шесть зубцов. Селится в нижней части сосен под толстой корой. От брачной камеры отходят один - три очень длинных (до 50, а изредка 70 см) широких (3 - 4 мм) маточных ходов, резко отпечатывающихся на заболони. Личиночные ходы короче маточных, слабо задевают заболонь, сильно расширяются в конце и заканчиваются куколочными колыбельками на внутренней поверхности коры.
Широко распространен в Европе. Встречается в горах, вредит пихте и ели на Кавказе, а в Сибири и на Дальнем Востоке - кедру. В Якутии, на Алтае, и в европейской части СССР - типичный вредитель сосны обыкновенной. Лёт обычно начинается в начале мая, однако высоко в горах и северных районах Сибири он наблюдается только в первой декаде июня и очень растянут. Молодое поколение жуков отрождается через 40 - 50 дней. Молодые жуки тут же приступают к дополнительному питанию, выгрызая звездчатой формы короткие ходы. Зимует в лесной подстилке или минных кодах под толстой корой. Генерация одногодовая, а в южных районах - двойная.
Вид светолюбивый, ксерофильный, заселяет ослабленные сосны в очагах корневой губки, на гарях, в изреженных насаждениях, в лесах, пострадавших от хвоегрызущих вредителей, особенно сибирского шелкопряда. Заселяет лесопродукцию на лесосеках, сильно размножается в местах выборочных рубок, хорошо идет на ловчие деревья.

Рис. 96. Вершинный и шестизубый короеды: 1 - вершинный короед, 2 - тачка вершинного короеда, 3 - ходы вершинного короеда, 4 - шестнзубый короед, 5 - тачка, 6 - ходы шестизубого короеда
Вершинный короед (Ips acuminatus Gyll.) (рис. 96). Жук длиной 2,2 - 3,7 мм, коричневый, блестящий, слабо волосистый; на вытянутой тачке три зубца. Лёт жуков происходит в начале мая. Селится жук в верхней части ослабленных сосен, где протачивает очень характерные ходы. От брачной камеры отходят один - восемь маточных ходов длиной от 5 до 50 см и шириной 2 мм. Маточные ходы забиты буровой мукой, личиночные ходы короткие, быстро расширяющиеся, редкие, глубоко отпечатываются на заболони. Генерация одногодовая, а на юге СССР - двойная. Вид исключительно светолюбивый, поселяется часто на соснах, зараженных смоляным раком, в изреженных насаждениях, особенна где велась подсочка или было размножение хвоегрызущих насекомых.
Четырехзубый гравер (Pityogenes quadridens Hart.). Жук длиной 1,5 - 2,3 мм, коричневый, тачка с каждой стороны имеет по четыре зубца. Заселяет вершины и толстые ветви сосен. По ходам и образу жизни напоминает вершинного короеда, но менее активен и светолюбив. Генерация обычно одногодовая, но может быть на юге двойная.
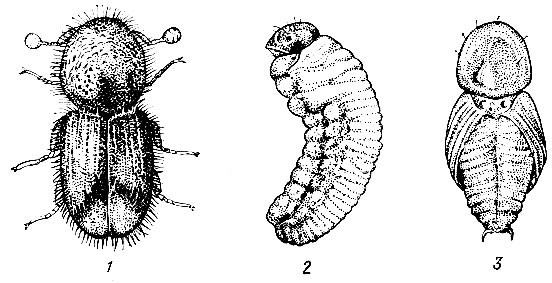
Рис. 97. Короед-типограф: 1 - жук, 2 - личинка, 3 - куколка
Короед пожарищ (Orthotomicus suturalis Gyll.). Жук длиной 2,5 - 3,5 мм, темно-бурый, блестящий, впадина на скате овальная, имеет по три зубца с каждой стороны, они сдвинуты внутрь впадины. Маточные ходы извилистые, резко отпечатываются на заболони, от брачной камеры отходит три - семь ходов, длина их до 3 см, ширина 1,5 мм. Личиночные ходы частые, длинные, извилистые. Селится на всех хвойных, предпочитает сосну и кедр. Лёт в мае, очень растянут. Молодые жуки отрождаются в. июле и проходят дополнительное питание, вбуравливаясь в древесину. Зимуют под корой, концентрируясь по 15 - 20 шт. в звездчатых ходах прикомлевой части ослабленных деревьев (Земкова, 1965). Генерация одногодовая. Активно влияет на усыхание древостоев, пройденных пожаром. Преобладает в жердняках, где заселяет деревья по всей их высоте.
Полосатый древесинник (Trypodendron lineatum Oliv.). Заселяет ослабленные деревья сосны, реже ели и других хвойных в области толстой коры одновременно с большим сосновым лубоедом, но прокладывает свои ходы в древесине. Генерация одногодовая, на юге (Крым, Кавказ) двойная, зимуют жуки в подстилке. Тенелюбивый, Гигрофильный вид, идет на ловчие деревья. Приносит большой технический вред и распространяет грибную болезнь - синеву древесины.
На тонких ветвях в вершинах крон сосен разного возраста селится целый ряд видов мелких короедов, которые при вершинном типе отмирания деревьев могут принести им значительный вред. Среди них очень распространены сибирский гравер (Pityogenes irkutensis Egg.), малый степной лубоед (Carpohoborus minimus Fabr.) и др.
На ели.Короед-типограф (Ips typographus L.) (рис. 97). Жук длиной 3,5 - 5 мм, темно-коричневый, отлогая впадина на скате надкрылий имеет с каждой стороны по четыре зубца, расположенных на равном расстоянии друг от друга; третий зубец самый крупный, утолщен на вершине. От брачной камеры в продольном направлении отходят один - четыре маточных хода длиной 10 - 15 см. Личиночные ходы частые, слегка извилистые, не задевают заболонь (рис.98).
Лёт в мае - июне. Жуки поселяются преимущественно в нижней и средней части стволов на старых и толстых, реже молодых, елях и деревьях других хвойных пород. Генерация одногодовая, на юге ареала - две генерации в год. Дополнительное питание - в местах развития. Зимуют жуки в минных ходах под корой или в лесной подстилке. Если развитие второго поколения задерживается, зимуют личинки и куколки в ходах и часто во время зимних морозов вымирают. Вид светолюбивый, пластичный.

Рис. 98. Ходы короеда-типографа
Типограф - опасный вредитель ели, заселяет ее во всех случаях ослабления, встречается в горах до 1800 м над уровнем моря, его массовые размножения неоднократно носили катастрофический характер.
Короед двойник (Ips duplicatus Sahib.). Очень похож на типографа, но немного меньше размером. На тачке с каждой стороны по четыре зуба, но два средних сближены и находятся на общем основании. Ходы немного уже и короче, чем у типографа, иногда слегка извилистые. Лёт начинается на несколько дней позднее, чем у типографа. Заселяет преимущественно более молодые и тонкие деревья, заходя на участки с тонкой корой. Вид светолюбивый, размножается в изреженных насаждениях, заселяет оставшийся подрост на лесосеках. Хорошо идет на ловчие деревья.
Еловый гравер (Pityogenes chalcographus L.). Жук длиной 2 - 2,9 мм, темно-бурый, с красновато-желтой узкой тачкой, снабженной с каждой стороны тремя зубцами. Ходы сложные, звездчатые. От брачной камеры отходят три - пять маточных ходов. Обычно сопутствует двум предыдущим видам, заселяя вершины и ветви деревьев разного размера и возраста. В южной части ареала ели может давать две генерации в год. Дополнительное питание в местах отрождения молодых жуков, там же они зимуют. Вид светолюбивый, пластичный, имеет очень широкое распространение. Кроме ели часто встречается и на сосне. Первым ослабленные деревья заселяет редко.
Большой еловый лубоед-дендроктон (Dendroctonus micans Kug.). Жук длиной 5,5 - 9 мм, темно-коричневый или черный. Короткий маточный ход сбоку расширен. Личинки грызут семейный ход в виде большой полости, залитый смолой и забитый опилками. Отрождение жуков происходит в августе. Они зимуют и на следующий год весной самки выгрызают маточные ходы и откладывают по 250 яиц. Зимуют личинки. Генерация обычно двухгодовая. Повреждает ель и сосну. В Грузии - опасный вредитель восточной ели, на юге Западной Сибири - сосновых культур, в европейской части СССР образует куртинные очаги в ельниках и заболоченных сосняках. Для биологического цикла дендроктона характерны сильно растянутая во времени фаза взрослого насекомого и личинки, которые можно обнаружить под корой деревьев в любое время года (Коломиец, Богданова, 1978).
Пушистый лубоед (Polygraphus polygraphias L.). Жук длиной 2,2 - 3 мм, черно-бурого цвета, покрыт чешуйками, отчего сверху надкрылья кажутся блестяще-серыми. Летает в мае - июне, имеет одну генерацию в год; предпочитает ельники жерднякового и среднего возраста, заселяет деревья с гладкой корой, начиная с 1 - 2 м высоты. Особенно распространен на гарях и в очагах корневой губки, в таежной части лесной зоны тяготеет к более изреженным, а в области широколиственных лесов к более густым насаждениям. Активно заселяет деревья при самом первоначальном ослаблении, на ловчие деревья идет не очень охотно.
Фиолетовый лубоед (Hylurgops palliatus Gyll.). Одинаково часто заселяет ель и сосну в области толстой и переходной коры; вид тенелюбивый и гигрофильный; широко распространен в лесной зоне, предпочитает бурелом и ветровал; первым растущие деревья не заселяет.
В таежной зоне на ели распространен целый ряд видов короедов. На тонких веточках в большом количестве размножается короед-микрограф (Pityophthorus micrographus L.), в средней части стволов - Хальцеходный лубоед (Xylechinus pilosus Ratz.), в нижней - короед автограф (Dryocoetes autographys Ratz.) и др.
В средней Азии на тяньшанской ели живет ряд короедов, отсутствующих на других хвойных породах. Они приносят большой вред и часто способствуют массовому отмиранию насаждений. Самый опасный и распространенный вид - горный киргизский короед (Ips hauseri Reitt.). Летает в мае, заселяет ослабленные старые и средневозрастные деревья, ветровал, южные опушки по склонам, недорубы на лесосеках. Это свето- и теплолюбивый вид, во многом напоминающий типографа и столь же опасный в условиях горных лесов.
На пихте. Сибирская пихта чаще всего заселяется короедами, живущими на ели. Типичным вредителем белокорой, цельнолистной и сахалинской пихты на Дальнем Востоке и о. Сахалине является белопихтовый полиграф (Polygraphus proximus Blandf.). Он активно заселяет пихту в очагах сибирского шелкопряда, в местах осыпей, оползней, буреломов и ветровалов, заселяет срубленные деревья и штабеля лесоматериалов. Лёт очень растянут и генерации спутаны; в Приморье, видимо, две, а в Хабаровском крае и горных лесах - одна. Дополнительное питание в местах отрождения жуков, которые зимуют.
На европейской, белой и кавказской пихтах широко распространен крючкозубый короед (Pityokteines curvidens Germ.). Он селится под толстой корой ослабленных и срубленных деревьев, образует очаги в местах эрозионных процессов, в расстроенных рубками и зараженных грибными болезнями лесах. Лёт в мае, генерация одногодовая, зимуют жуки в минных ходах, в толстой коре растущих деревьев. Приносит значительный вред в Карпатах. Там же распространен западный крифал (Cryphalus piceae Ratz.), а в Приморье и на о. Сахалин - японский крифал (С. piceus Egg.). Все крифалы - типичные вторичные вредители и редко первыми заселяют ослабленные деревья.
На лиственнице (сибирской и даурской). По всему ареалу лиственницы распространен продолговатый короед (Ips subelongatus Motsch.). Жук длиной 5 - 6 мм; тело очень вытянутое, тачка с четырьмя зубцами с каждой стороны. Типичный обитатель лиственничных лесов. Он прокладывает длинные ходы, напоминающие таковые у шестизубого короеда, в Нижней и средней части ствола, изредка под тонкой корой. Лёт жуков в мае - июне, генерация одногодовая, дополнительное питание в местах развития, зимуют жуки в минных ходах и в верхнем слое почвы под мхом. Вид пластичный, однако больше тяготеет к освещенным, хорошо прогреваемым местам, где нападает на ослабленные, но еще жизнеспособные деревья и неокоренные лесоматериалы зимней заготовки. Особенно большой вред приносит в очагах сибирского шелкопряда. Может поселяться на сибирском кедре и изредка на ели.
На лиственнице встречается заболонник Моравитца (Scolytus morawitzi Sem.) -единственный заболонник, живущий на хвойных породах, однако массового распространения он HeimeeT. Гораздо чаще встречается байкальский лесовик (Dryocpetes baicalicus Reit.), но большого значения он также не имеет. На лиственнице поселяются короеды с сосны и ели: шестизубый гравер, типограф и др.
На лиственных породах. Встречается много видов короедов. Однако большое хозяйственное значение имеют не все виды. Здесь будут рассмотрены Только наиболее обычные и вредные виды.
Березовый заболонник (Scolytus raizeburgi Jans.). Распространен по ареалу березы до Дальнего Востока. Заселяет нижнюю и среднюю часть стволов, проделывая под корой простые продольные ходы с множеством круглых отверстий вдоль маточных ходов, по которым хорошо отличать заселенные им деревья. Лёт жуков в мае, дополнительное питание в коре возле почек, кладка яиц в июне, зимует личинка в ходах, генерация одногодовая.
Вид пластичный, однако предпочитает селиться на деревьях, растущих единично или группами по опушкам леса, вблизи дорог, в изреженных насаждениях и парках. Заселяет сильно ослабленные и усыхающие деревья.
В Приморье широко распространен очень похожий на березового амурский заболонник (S. amurensis Egg.). Чаще всего он встречается в ясеневых заболоченных лесах с примесью маньчжурской березы, где заселяет ветровальные деревья березы, а иногда вызывает их вершинное отмирание, повреждая вершину и толстые ветви.
Дубовый заболонник (S. intricatus Ratz.) Рис. 99). Встречается по всему ареалу дуба в европейской части СССР и на Кавказе, где заселяет местные виды дубов (армянский и др.). Изредка поселяется на грабе, каштане, березе, клене и других породах, но вредителем их не числится. Заселяет преимущественно отмирающие молодые дубы по всему стволу, а на более старых деревьях избегает толстой коры и заселяет их среднюю часть, вершины и ветви. Маточные ходы поперечные, простые, короткие.

Рис. 99. Ходы дубового заболонника
Лёт жуков в июне, после чего они в течение 10 - 12 дней дополнительно питаются в кронах совершенно здоровых дубов. Для этого жуки внедряются в тонкие концевые веточки в местах их сочленений. Первоначально они делают поверхностные погрызы и лишь спустя несколько дней полностью уходят в глубь ветви, делая на ней ход длиной около 0,5 см. Яйцекладка во второй половине июня - июле. Зимуют личинки, они окукливаются весной следующего года в конце апреля - начале мая. Генерация одногодовая.
Во время дополнительного питания жуки очень часто разносят инфекцию сосудистого заболевания дуба (Ceratocystis), заражая им по мере разлета все новые и новые деревья (Эдельман и Малышева, 1959). Вне очагов заболевания это довольно пассивный вредитель.

Рис. 100. Ходы ильмовых заболонников: слева - струйчатого, справа - разрушителя
Ильмовые породы. Много видов заболонников и лубоедов встречается на ильмовых. Массовое размножение периодически наблюдается в степной и лесостепной зонах. Заболонники переносят инфекцию голландской болезни (Graphium ulmi) при дополнительном питании. Дополнительное питание жуков проходит в сочленениях тонких веточек так же, как у дубового заболонника. Зараженные голландской болезнью деревья теряют устойчивость и заселяются заболонниками, хотя внешне выглядят еще совершенно здоровыми. При этом происходит куртинное усыхание ильмовых, обусловленное небольшим радиусом разлета заболонников, обычно равным 70 - 130 м.
Самыми распространенными видами заболонников, переносящих голландскую болезнь, являются: заболонник-разрушитель (Scolytus scolytus F.), струйчатый заболонник (S. multistriatus March.) (рис. 100), Оба заболонника заселяют берест, вяз и ильм разного возраста, преимущественно старше 8 - 10 лет.
Заболонник-разрушитель тяготеет к нижней части стволов, а струйчатый - к средней и верхней частям, заселяя также ветви.
Заболонник-разрушитель в степной части предпочитает берест, на котором развиваются две полные и частичная третья генерации. На вязе вылет молодых жуков разрушителя задерживается на две-три недели. Заболонник струйчатый на бересте развивается сходным образом, а на более предпочитаемой им породе - вязе - бывает только две генерации. Лёт жуков, заселение ими деревьев и развитие молодого поколения у обоих видов очень растянуты. В период с мая по сентябрь одновременно можно встретить личинок, куколок, молодых жуков и начало новых поселений. Севернее число генераций в год сокращается до одной, а на Кавказе и в Средней Азии увеличивается до четырех. В ряде областей (юго-восток европейской части СССР) заболонник-разрушитель на вязе замещается близко родственным заболонником морщинистолобым (Scolytus sulcifrons Rey.).
В верхней части стволов и на ветвях ильмовых селятся заболонник пигмей (S. pygmaeus F.) и заболонник Кирша (S. Kirschi Seal.). Они часто заселяют ильмовые в байрачных лесах, по балкам, в пойменных лесах и полезащитных полосах и также являются переносчиками голландской болезни. Заболонник пигмей развивается аналогично струйчатому, а зоболонник Кирша имеет только одну генерацию в год, лёт жуков в июне - июле. На Северном Кавказе, в Крыму и некоторых других районах распространен заболонник Зайцева (S. Zaitzevi But.), биолоия которого сходна с заболонником Кирша.
Наряду с заболонниками деревья заселяют ильмовый лубоед (Pteleobius vittatus Fabr.) и лубоед Краатца (P. Kraatzi Eichh.). Они появляются в апреле - мае, вылет молодых жуков в августе. Жуки зимуют в толще коры комлевой части стволов растущих деревьев.
Ясень. Большой вред ясеню наносят ясеневые лубоеды. Из них наиболее распространен и опасен малый (пестрый) ясеневый лубоед (Hylesinus fraxini Panz.). Он заселяет ясени разного возраста, преимущественно молодые и средневозрастные, в пределах европейской части СССР. При массовом размножении жуки заселяют внешне совершенно здоровые деревья, часто вызывая усыхания древостоя. Лёт в мае (на юге с середины апреля), ходы под тонкой и средней корой поперечные в виде фигурных скобок. Генерация одногодовая. Дополнительное питание в минных ходах на тонких частях ствола, а зимовка в таких же ходах в толстой коре, в одних и тех же местах из года в год. В результате возникают болезненные наросты в виде розеток.
В лесостепной и степной зонах также часто встречаются большой ясеневый лубоед (Hylesinus crenatus Fabr.), там же, и на Кавказе - масличный лубоед (Н. oleiperda F.), а в лесах Дальнего Востока - пестрый уссурийский лубоед (Н. eos Spess.) и черный лубоед (Н. laticollis Blandf.).
На плодовых деревьях повсеместно распространен морщинистый заболонник (Scolytus rugulosus Ratz.). Он повреждает также черемуху, боярышник, рябину, кизил; имеет ряд подвидов, распространенных на Кавказе и в Средней Азии. Нападает на ослабленные деревья, заселяет стволы с толстой корой, а на старых деревьях занимает их среднюю и верхнюю части и сучья. Генерация одногодовая, на юге - двойная, дополнительное питание в коре у основания почек.
Большой вред лиственным породам наносят короеды древесинники. Самый обычный из них лестничный лиственный древесинник (Trypodendron signatum 01.), внешне мало отличается от хвойного древесинника, ведет такой же образ жизни, но заселяет всегда только лиственные породы, особенно дуб, березу и ольху. Широко распространены также непарные древесинники, получившие свое название вследствие разницы между самцами и самками. Самки откладывают яйца кучкой; личинки грызут совместный семейный ход или расползаются по маточным ходам. Наиболее просто ходы устроены у многоядного древесинника (Xyleborus saxeseni Ratz.), нападающего на многие лиственные породы (дуб, бук, ольху, граб, лещину и др.), а на Дальнем Востоке и на хвойные. Лёт в конце мая - июне. Генерация одногодовая. Заселяет ослабленные деревья (рис. 101).
У западного непарного древесинника (Xyleborus dispar Fabr.) ход построен иначе, чем у других непарных древесинников. Самка сначала точит канал перпендикулярно поверхности ствола на 3 - 6 см, где ход поворачивается по годичному кольцу то в одну, то в другую сторону, кольцуя стволик. От этого первичного хода самка откладывает кучками яйца. Лёт жуков в июне; зимуют в ходах молодые жуки. Генерация одногодовая. Вид многоядный, особенно сильно повреждает дуб, бук, клен, березу и плодовые. В условиях степной зоны считается очень вредным видом, встречается также в Сибири и на Кавказе.
Все древесинники одновременно и технические вредители, борьба с ними обязательна (гл. X).
Усачи (Cerambycidae)
Семейство усачей объединяет на земном шаре около 17 тыс. видов жуков, из которых у нас в СССР живет только 1500 видов. Усачи питаются растениями, причем большая часть усачей живет за счет деревьев и кустарников и называется дровосеками.
Размеры жуков усачей колеблются от 3 до 60 мм. Тело удлиненное, чаще всего покрыто волосками. Ноги длинные, голени с шипами, лапки 4-члениковые. Голова свободная. Усики длиннее половины тела и часто превосходят его в 1,5 - 2 раза. Всем дровосекам присуща способность "закидывать усики на спину", т. е. загибать их назад, что не могут делать другие жуки. Надкрылья покрывают все брюшко; изредка надкрылья бывают сильно укорочены и брюшко остается частично неприкрытым (коротконадкрылые усачи p. Molorchus и др.). Большинство усачей способно издавать скрипучий звук при трении среднегруди о переднегрудь. Личинки усачей снабжены небольшими по размеру, но хороша приспособленными для разгрызания древесины челюстями. С их помощью они прогрызают длинные и широкие ходы в древесине. Взрослые личинки цилиндрические или слегка уплощенные, белые. Голова и челюсти твердые, коричневого цвета. Передний конец тела более широкий за счет расширенной передней груди. На члениках имеются особые площадки - "мозоли", упираясь которыми личинки передвигаются в своих ходах.
Личинки усачей сильно разнятся по строению тела, размерам головы, отсутствию или наличию ног, однако на первый взгляд все они похожи друг на друга, имеют сходство в основных чертах строения и легко отличимы от личинок других стволовых вредителей.
Лёт дровосеков проходит в разное время и очень растянут, так как условия развития их в дереве весьма изменчивы и зависят от его состояния, возраста, местообитания и разнокачественности отдельных тканей, которыми питаются личинки. Ряд видов летает рано весной, одновременно с сосновыми лубоедами, большинство в июне - июле, у некоторых усачей лёт затягивается до конца августа.
Календарные сроки лёта отдельных видов обусловлены погодными условиями года и географическим районом.
Самки откладывают белые продолговато-овальные яйца в трещины и щели коры и древесины или в углубления, выгрызаемые в коре (насечки). Развитие яйца длится 10 - 20 дней. Вышедшие из яиц личинки начинают грызть в лубе ходы.
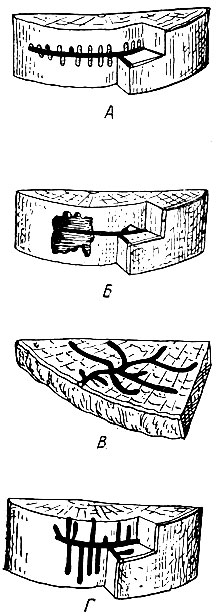
Рис. 101. Ходы древесинников. А - лестничного; Б - семейного, В - короеда-монографа, Г - непарного короеда
По образу жизни личинок можно разделить на несколько групп:
личинки всю свою жизнь проводят под корой, где грызут ходы и окукливаются;
личинки большую часть жизни проводят под корой, выгрызают длинные ходы, а перед окукливанием уходят в древесину и проделывают небольшой крючковидной формы ход;
личинки живут под корой недолго, выгрызают небольшую площадку в лубе и затем проделывают в древесине длинные ходы, подводя их перед окукливанием к поверхности древесины; такой ход после вылета жука имеет скобовидную форму (рис. 103).
Личинки всю свою жизнь проводят в древесине. У большинства личинок дровосеков ходы округло-овальные. Личинки обычно зимуют один-два раза и весной окукливаются. Длительность развития личинок может изменяться в зависимости от состояния древесины и условий питания.
Перед окукливанием личинка почти всегда устраивает особую колыбельку, т. е. расширяет конец хода. В колыбельке личинка окукливается. Если личинка проделывает крючковидный ход, то она переворачивается перед окукливанием головой к выходу. В том случае, если личинка прогрызает скобовидный ход, она не переворачивается, а молодой жук разгрызает оставшееся пространство. Перед окукливанием личинка обычно отделяет колыбельку от остального хода пробкой из опилок. Развитие куколки длится 10-12 дней.
Генерация усачей различная. У многих видов она одногодовая, у других длится два-три года. На длительность генерации влияют условия питания личинок. При неблагоприятных условиях генерация затягивается на несколько лет.
После вылета молодые жуки у многих видов проходят дополнительное питание сочным лубом молодых побегов в кронах (черные усачи), выгрызают ткани листа (осиновые скрипуны) или питаются пыльцой цветков (большее число видов).
Среди усачей преобладают олигофаги, питающиеся рядом близких по своему происхождению древесных пород. Все усачи делятся на вредителей хвойных и лиственных древесных пород. С хвойных пород на лиственные усачи переходят очень редко, хотя такие случаи известны. Так, черный большой хвойный усач в Восточной Сибири развивается на пихте, ели и березе. Среди усачей одного рода очень часто одни виды связаны с хвойными, а другие с лиственными породами.
В пределах хвойных и лиственных пород усачи также отдают предпочтение известным видам древесных пород. Так, серый длинноусый усач и усач сосновых вершин очень редко развиваются на ели, а блестящегрудый усач - на сосне. Такая же картина наблюдается и при питании на лиственных породах. Переход с одной породы на другую часто ведет к задержке в развитии, сроках вылета, плодовитости, изменяет величину тела. Предпочтение той или иной древесной породе зависит от разных географических районов.
Каждый вид усача занимает на дереве определенный район поселения (корни, тонкие ветви, часть ствола с грубой или тонкой корой) и редко изменяет своим привычкам.
Некоторые усачи очень активны и заселяют внешне совершенно здоровые деревья (большой и малый осиновые усачи и др.). Большинство нападает на ослабленные деревья. Среди усачей встречаются свето- и теплолюбивые формы и тенелюбы, большинство видов обладает большой биологической пластичностью.
Изменение численности усачей зависит от комплекса факторов, рассмотренных выше для всей группы стволовых вредителей.
Обилие видов усачей не позволяет в кратком курсе дать их сколько-нибудь удовлетворительный обзор. Поэтому ниже приводится характеристика только самых распространенных и вредных видов.
Черные хвойные усачи (p. Monochamus). Крупные насекомые, тело у них более или менее вытянуто. Оно обычно блестящее, черное или смоляно-черное. Надкрылья длинные, в большинстве случаев сильно вытянутые, слегка суженные к концу, обычно закругленные, с грубой скульптурой и густыми более светлыми волосками. Усики более или менее тонкие, в 1,5 раза длиннее тела, с сильно утолщенным одним члеником.
Личинки белые, безногие, голова черная, тело к концу несколько сужена. Размеры личинок зависят от вида и достигают у пихтового усача 4 - 6 см. Они сначала выгрызают большие неправильной формы площадки под корой, а затем углубляются в древесину, где делают очень крупные, скобообразные ходы. Так, у пихтового усача длина вертикальной части хода 15 см, общая длина хода 30 - 40 см, а ширина 1 - 2 см. Лётное отверстие 1 - 1,2 см.
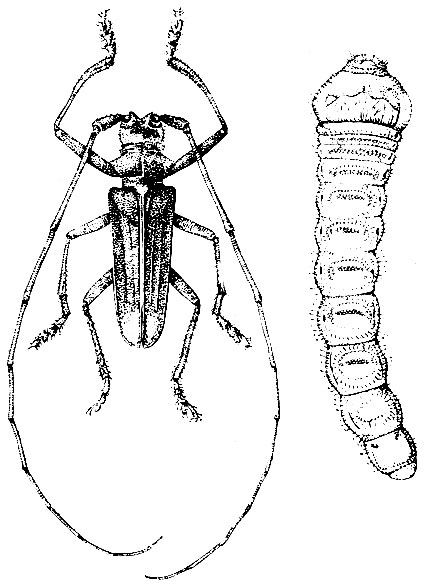
Рис. 102. Пихтовый усач и его личинка
Все черные усачи проходят дополнительное питание в кронах деревьев, повреждая побеги и ветви.
В лесах СССР распространены следующие виды черных усачей: черный большой хвойный усач (М. urussovi Fich.), черный сосновый усач (М. galloprovincialis Gelb.), черный малый хвойной усач (М. sutor L.), бархатно-пятнистый черный хвойной усач (М. saltuarius Gelb.) и черный хвойный крапчатый усач (М. impluviatus Motsch). Последние два вида встречаются только в лесной зоне азиатской части СССР, остальные повсеместно.
Черный большой хвойный усач (Monochamus urussovi Fisch.). Наибольший вред приносит в лесах Сибири и на Дальнем Востоке, размножаясь в огромных количествах в очагах сибирского шелкопряда и пихтовой пяденицы, на гарях, а также на лесных складах и в местах крупных лесозаготовок. В европейской части СССР этот вид широко распространен в северной части лесной зоны и сравнительно малочислен в ее южной части (рис. 102).
Массовый лёт жуков в лесах Сибири начинается при среднесуточной температуре выше 13° С, при температуре более 20° С интенсивность лёта усиливается, а при ее падении ниже 10° С - уменьшается. Обычно лёт наступает в третьей декаде июня, бывает массовым в первых двух декадах июля и заканчивается в середине сентября. На о. Сахалин лёт начинается почти на месяц позднее (Криволуцкая, 1961), а в южной части лесной зоны - на 1 - 1,5 недели раньше.
Жуки живут около двух месяцев (по Прозорову, в среднем 51 - 52 дня) и в течение этого времени проходят дополнительное питание в кронах деревьев разного возраста (начиная с 10 - 12 лет). Обычно жук выбирает тонкую ветку, помещается вдоль нее, скусывает имеющиеся хвоинки и затем начинает соскабливать кору, оголяя древесину полосой по длине ветви на 1 -10 см.
Откладка яиц начинается через 12 - 20 дней после появления первых жуков, а еще через 10 - 12 дней достигает максимума. В южной части лесной зоны европейской части СССР откладка яиц начинается через 8 - 12 дней после появления первых жуков. Для откладки яиц самка выгрызает в коре узкую щель - насечку, в которую с помощью яйцеклада вводит на глубину 2 - 3 мм одно, реже два яйца. Средняя плодовитость одной самки 14 яиц, максимальная 33. Фаза яйца длится 13 - 29 дней; для его развития нужна сумма температур около 250° С.
Вышедшая из яйца личинка имеет длину тела около 3 - 5 мм и грызет ход в толще коры, а далее в заболони и зимует в первом или во втором возрасте. Во втором возрасте она расширяет ход под корой и углубляется в древесину до 5 см. Вторая линька происходит в июне следующего года. В это время личинка еще дальше проникает в глубь ствола, все время очищает ходы и периодически возвращается для питания под кору. В четвертом возрасте личинки уже редко посещают подкоровое пространство. В последнем - пятом возрасте, который наступает после четвертой линьки осенью второго года или весной на третий год после фазы яйца, личинка уже не возвращается под кору, а заканчивает свой ход на расстоянии 1,5 - 2 см от поверхности ствола и в конце его устраивает куколочную колыбельку, в которой на третий год превращается в куколку (рис. 103). Фаза куколки длится 25 - 26 дней. Генерация двухгодовая, но при благоприятных условиях развития часть популяции может завершать жизненный цикл в один год.
Черный большой хвойный усач может заселять все хвойные породы тайги, но предпочитает пихту, а в лесной зоне европейской части СССР - ель. Кроме того, в условиях Забайкалья и в Монголии он поселяется на березе, где успешно завершает свое раз-питие, включая дополнительное питание (Тальман, 1940; Гречкин, 1960).
Жуки усача светолюбивы и в первую очередь селятся в окнах, по опушкам и в изреженных насаждениях, однако при массовом размножении эти особенности стираются и жуки распространяются независимо от освещения. Они заселяют поваленные и стоящие деревья, но плотнее - первые.
В очагах сибирского шелкопряда в первую очередь заселяется пихта, затем ель и сибирский кедр. На лиственнице усач встречается реже и играет второстепенную роль. Он предпочитает деревья с диаметром толще 24 см, а деревья тонкие (8 - 12 см) избегает (Катаев, 1959). Заселяется преимущественно нижняя и средняя часть ствола дерева, где развитие идет несколько быстрее, а смертность личинок меньше. В целом же у усача наблюдается очень большая эмбриональная смертность и гибель до 50% личинок (Лонщаков, Маслов, Мишель, 1958), хотя деятельность энтомофагов сравнительно малоактивна. Много личинок истребляется дятлами, особенно желной (Прозоров, 1958).
Черный сосновый усач (Monochamus galloprovincialis Germ.) (рис. 104). Опасный вредитель сосновых лесов в южной части лесной зоны, лесостепи и степной зоны европейской части СССР, ленточных боров Западной Сибири и Казахстана. Он размножается в оча- гах корневой губки, на гарях, в очагах хвоегрызущих насекомых, в сосняках, сильно ослабленных засухой, подкорным сосновым клопом, в местах лесозаготовок и на складах древесины.
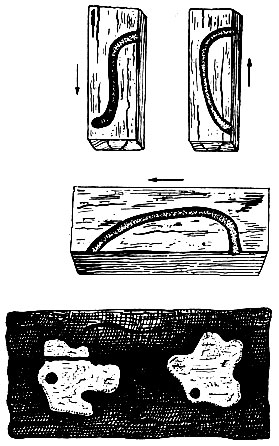
Рис. 103. Схема ходов пихтового усача
Лёт жуков начинается в первой декаде июня, в начале июля 90% их покидает древесину (Кузнецова, 1956). Жуки выходят неполовозрелыми и проходят дополнительное питание на ветвях сосен, обгладывая свежую тонкую кору. Жуки живут до 70 дней, но уже по прошествии 5 - 7 дней самки начинают откладку яиц в насечки. Личинки появляются в середине июля. Они питаются корой, лубом, заболонью и верхними слоями древесины. В начале августа личинки углубляются в древесину. Как и у пихтового усача, личинки на протяжении всего развития периодически выползают из ходов в подкоровое пространство для питания лубом и заболонью. В связи с этим они очищают и расширяют свои ходы, иногда проделывают дополнительные отверстия для выбрасывания "опилок". Конец хода в древесине личинка не доводит на 1 -1,5 см до поверхности и в конце его устраивает кукольную колыбельку, где зимует. Окукливание происходит в мае. Генерация одногодовая, но часть личинок развивается по двухгодовому циклу.
Жуки усача светолюбивы и предпочитают изреженные, хорошо прогреваемые насаждения. В смешанных насаждениях численность усача резко падает. Он селится по всему стволу, при этом в комлевой части больше отрождается самок, а в верхней - самцов.
Биология остальных черных хвойных усачей очень сходна с вышеописанными двумя ведущими видами. Они также летают с конца июня до августа и развиваются ни одногодовому циклу, заселяя различные хвойные породы.
Усачи тетропиумы (p. Tetropium). Отличаются меньшими размерами и уплощенным телом жуков. Усики достигают половины тела, переднеспинка в длину почти такая же, как и в ширину, надкрылья едва выпуклые, умеренно длинные, параллельные, обычно гораздо шире переднеспинки, на вершине закругленные, черные или каштановые; тело черное.
Для личинок характерно наличие коротких ног; они желтовато-белые с более темными переднеспинкой и головой, челюсти черные, голова почти сердцевидная, сверху с продольной бороздкой посредине. Размеры личинок зависят от вида и достигают 20 мм. Они выгрызают площадки под корой, а затем делают крючковидный ход в древесине, где окукливаются.
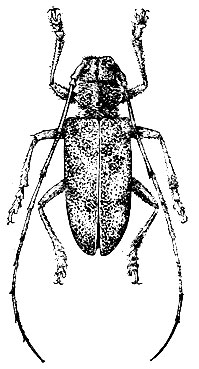
Рис. 104. Черный сосновый усач
В лесах СССР распространены следующие виды: блестяще-грудый (Т. casianeum L.) и матовогрудый (Т. fuscum F.) еловые усачи, лиственничный дровосек Габриэля (Т. gabrieli Weise.), тонкоусый еловый усач (Т. gracilicorne Reitt.) и семиреченский еловый усач (Т. staudingeri Pic). Образ жизни у всех этих видов имеет много общего. Все они повреждают хвойные породы и являются активными вредителями, нападая первыми на ослабленные деревья в очагах хвоегрызущих насекомых, опенка и корневой губки, заселяя по опушкам ослабленные деревья, ветровал и бурелом, древесину в местах лесозаготовок.
Блестящегрудый еловый усач (Tetropium castaneum L., p. 105). Распространен в СССР повсеместно. Лёт в мае - июне; самка откладывает яйца в щели коры деревьев, личинка выгрызает под корой широкие ходы неправильной формы, глубоко задевающие заболонь, и через 20 - 25 дней уходит, делает крючковатый ход в Древесину на глубину 2 - 4 см. В этом ходе она зимует, а весной поворачивается к выходу и окукливается. Генерация одногодовая.
Усач поселяется на деревьях ели разного диаметра, в различных экологических условиях, преимущественно в затененных местах, заселяет комлевую часть стволов, а в Сибири, в очагах сибирского шелкопряда, поселяется раньше пихтового усача и занимает весь ствол, повреждая кроме ели еще сибирский кедр. На других хвойных встречается редко.
Алтайский лиственничный усач (Xylotrechus altaicus Gelb.). Жук длиной 11 - 23 мм. Самки крупнее самцов. Тело вытянутое, узкое, бурого цвета, ноги длинные, переднеспинка массивная, почти шаровидная. Надкрылья серовато-бурые. Распространен от Урала до Тихого океана.

Рис. 105. Блестящегрудый усач и его ходы
Массовый лёт в июле. Жуки не питаются. Самки интенсивно откладывают яйца в первые пять - шесть дней, а через две недели прекращают кладку совсем. Они помещают яйца в щели коры поодиночке, преимущественно на южную сторону деревьев, размещая их по всей высоте ствола. Плодовитость одной самки - 50 - 102 яйца (Рожков, 1981). Фаза яйца 13 - 16 дней. Вылупившаяся личинка безногая, белая, с рыжеватым оттенком. Длина взрослой личинки около 32 мм. Личинка проходит пять возрастов. После выхода их яйца личинка вгрызается в кору, разрушает луб, зимует в коре. Весной под корой прокладывает ход по окружности ствола. и в июле - августе углубляется в древесину, где зимует вторично. После второй зимовки личинка подходит к периферии ствола, устраивает куколочную колыбельку и окукливается. Фаза куколки 16 - 20 дней, генерация двухгодовая.
Прокладываемые личинками в наружных слоях заболони ходы очень характерны. Они имеют поперечные направления и не пересекаются даже при очень плотном расположении, на всем протяжении забиты опилками. Поврежденные деревья очень характерны и распознаются по описанным выше ходам (рис. 106).

Рис. 106. Ходы алтайского усача под корой лиственницы
Вид светло- и теплолюбивый. Очаги возникают в первую очередь в старых, изреженных насаждениях, поврежденных пожаром или хвоегрызущими вредителями. Это физиологически активный вид. Он заселяет деревья при незначительном снижении ими резистентности. Достигая высокой численности в очагах размножения, он поселяется и на здоровых деревьях. Характерной особенностью усача является образование новых очагов за счет миграции. По утверждению А. С. Рожкова (1981), лиственничный усач имеет очень большое хозяйственное значение. Он самый опасный вредитель лиственницы. Другим древесным породам не вредит - строгий монофаг.
Серый длинноусый усач (Acanthoclnus aedilis L.). Тело плоское, светло-бурое, подкрылья серые с темными перевязями. Длина 13 -20 мм. Усики в 1,5 - 3 раза длиннее тела. Самый обычный обитатель сосновых лесов. Повсеместно встречается в большом количестве, но нападает, как правило, только на усыхающие и поваленные деревья, пни, ветровал, бурелом. Древесине не наносит вреда, так как личинка грызет широкие ходы неправильной формы только в коре и лубе. Личинка безногая, белая, длина 30 - 34 мм.
Лёт усача начинается очень рано, обычно в конце апреля - мае, он сильно растянут, и жуков можно встретить до августа месяца. Развитие идет быстро, и осенью под корой в овальных колыбельках находятся молодые жуки. При более позднем отрождении личинок они зимуют, и молодые жуки появляются только весной. Генерация одногодовая.
При большой численности личинки усача разъедают все подкоровое пространство, забивая его спрессованными опилками бурого цвета, и тем самым препятствуют поселению других стволовых вредителей, особенно короедов.
В лесах Сибири и на Дальнем Востоке повреждают хвойные породы и другие представители p. Acanthoclnus: сибирский серый длинноусый усач (A. carlnulatus Gelb.) и малый серый длинноусый усач (A. griseus F.).
Рагий ребристый (Rhagium inquisitor L.) сопутствует серому длинноусому усачу, имеет такой же цикл развития. Его личинки с головой, окрашенной в яркий оранжево-коричневый цвет, живут и окукливаются под корой сухостойных деревьев хвойных пород. Этот вид очень широко распространен, но вреда не приносит. Другие виды этого рода (Rh. mordax Dg. Rh. sycophanta Schr.) развиваются за счет лиственных пород и тоже практически безвредны.
В комлевой части деревьев хвойных пород и свежих пнях живет три вида широко распространенных усачей: бурый комлевой усач (Criocephalus rusticus L.), черный ребристый усач (Asemum striatum L.) и короткоусый усач (Spondylis buprestoides L.). Заметный вред наносит только первый из них. Два других приносят скорее пользу, ускоряя разрушение пней и биологический круговорот в экосистемах с доминированием двойных (преимущественно сосны) пород. В литературе, однако, имеется ряд указаний на вред, наносимый личинками этих усачей в сухих сосновых борах.
Бурый комлевой (или деревенский) усач (Criocephalus rusticus L.) приносит технический вред, а также участвует в комплексе весенней фенологической группировки стволовых вредителей, заселяющих ослабленные различными факторами (пожары, корневая губка и т. д.) деревья. Это крупный жук рыже-бурого цвета, длина 10 - 27 мм, усики короткие. Жуки ведут ночной образ жизни, охотна летят на свет в дома. Лёт в июне - июле. Самки откладывают яйца в пни, корни и нижнюю часть отмирающих сосен, реже других хвойных пород.
Личинка желтовато-белого цвета с черно-бурыми челюстями, длиной до 33 мм. Она сначала живет под корой в области толстых корней или прикорневой части ствола, затем уходит в древесину и прокладывает продольные ходы, забивая их буроватой мукой. В последнем возрасте личинка устраивает колыбельку для окукливания и выгрызает выход к боковой поверхности, забивая eго крупной буровой мукой. Фаза куколки длится три-четыре недели. Отрождающиеся жуки в дополнительном питании не нуждаются и сразу приступают к спариванию. В зависимости от субстрата генерация длится один - три года. Предпочитает сосну. Часто встречается в древесине холодных построек, неокоренных бревнах, телеграфных столбах.
Лиственные породы повреждают многие виды усачей. Ниже приводятся главнейшие из них.
Большой дубовый усач (Cerambyx cerdo L.) повреждал дубовые леса на Украине (к западу от Днепра) и особенно в Крыму, Грузии и Краснодарском крае. Очень красивый большой жук длиной до 6 5мм (рис. 107. Теперь стал редким видом и нуждается в охране.
Лёт жуков с половины мая до августа. Самка откладывает яйца по одному в трещины коры, всего до 100 яиц. Фаза яйца длится 10 - 15 дней. Личинка первый год грызет ход под корой, после зимовки углубляется в древесину, где проделывает неправильный канал шириной до 3 см, вновь зимует и на третий год окукливается в конце хода. Из куколки в июле - августе отрождается жук, но покидает дерево только весной и дополнительно питается соком дуба. Генерация трехгодовая.
Вид светолюбивый, поселяется на южных опушках, в изреженных, преимущественно старых насаждениях порослевого происхождения, где заселяет в первую очередь хорошо освещенные самые толстые, внешне вполне жизнеспособные дубы.
На Кавказе наряду с дубовым распространен похожий на нега большой плодовый усач (Cerambyx dux Fald.), повреждающий дуб, бук и плодовые породы, а в дубравах лесостепи - малый дубовый усач (С. scopolli Fussl.). Он повреждает кроме дуба бук, граб, ясень, клен, ильмовые и плодовые, но очагов обычно не образует и хозяйственное значение невелико.
Пестрые дубовые усачи (Plagionotus arcuatus L., P. detritus L.). Очень широко распространены в границах ареала дуба в европейской части СССР и на Кавказе. Издали немного по раскраске напоминают ос. Тело черное с дуговидными желтыми полосами у одного вида (P. arcutus L.) и широкими желтыми перетяжками - у другого (P. detritus L.). Массовый лёт в июне, откладка яиц в щели коры по всему стволу, личинки грызут длинные продольные ходы, расширяющиеся по мере их роста и глубоко задевающие заболонь. На стоящих дубах они направлены снизу вверх, а на лежащих деревьях и бревнах имеют неопределенное направление. Через 30 - 40 дней после выхода из яиц личинки прогрызают овальные отверстия и уходят в древесину на глубину 2-4 см, затем круто, почти под прямым углом, загибаются вниз и грызут ход вдоль волокон древесины длиной до 3 - 5 см. В конце таких ходов личинки зимуют, заткнув горизонтальную часть хода пробкой из огрызков древесины, а весной расширяют ход, переворачиваются головой к выходу и окукливаются. Фаза куколки продолжается около 20 дней. Молодые жуки расширяют входные отверстия. Генерация одногодовая. Эти виды способствуют отмиранию часто еще жизнеспособных деревьев и наносят большой технический вред, приводя в негодность дубовую древесину. Пластичные виды встречаются в самых разнообразных условиях.
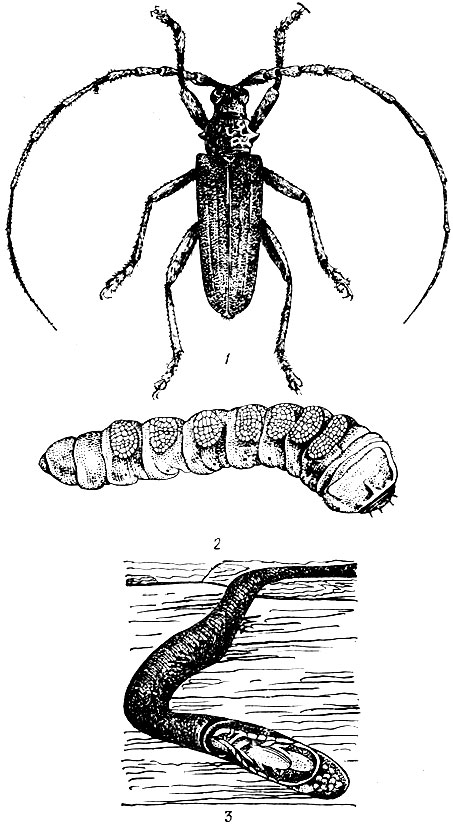
Рис. 107. Большой дубовый усач: 1 - жук, 2 - личинка, 3 - ходы
Желтопятнистый глазчатый усач (Mesosa myops Dalm.) распространен повсеместно, включая и Дальний Восток. Он повреждает большинство лиственных пород, особенно дуб. Лёт с июня до конца августа; личинка делает длинные ходы под корой, где и окукливается, зимуют жуки, дополнительное питание на коре стволов и ветвей, генерация одногодовая. Очень пластичный вид, встречается в самых различных экологических условиях.

Рис. 108. Большой осиновый скрипун
На ослабленных и поваленных дубах поселяется еще целый ряд усачей, личинки которых прогрызают довольно глубокие ходы в древесине. Эти усачи открывают ворота грибной инфекции и сильно снижают технические качества древесины. Главными видами являются: краснокрылый усач Келера (Purpuricenus kaehleri L.), красный дубовый дровосек (Phyrrhidium sanguineum L.), усач антилопа (Xylotrechus antilope Schonh.). Они летают с конца мая до июля. Личинки зимуют в древесине. Генерация одногодовая. Жуки заселяют деревья разного диаметра, однако предпочитают дубы средней толщины, молодые, средневозрастные.
Большой осиновый усач (Saperda carcharias L.). Этот вид широко распространен в европейской части СССР и Сибири (рис. 108).
Жук длиной 21 - 28 мм. Светло-коричневый или серый благодаря покрывающим его волоскам. У самцов надкрылья суживаются кзади, у самок они почти параллельны. Последние членики усиков в черных колечках. Лёт с конца июня до сентября (пик в июле). Жуки проходят дополнительное питание, выгрызая в листьях осины и тополей округлые дыры, а на побегах, тонких стволиках и ветвях - поперечные щелевидные погрызы коры. После спаривания самки делают в комлевой части растущих деревьев насечки и откладывают по одному яйцу. Плодовитость одной самки 50 - 60 яиц.
Личинка выгрызает сначала в заболони неправильной формы полость и затем углубляется в древесину, где прогрызает длинный (до 1,5 м) вертикальный ход, в котором зимует вторично, а весной третьего года делает боковой лётный ход и окукливается в верхнем конце вертикального хода на предварительно подготовленной пробке из волокнистых стружек. Молодой жук, разрушив пробку, пробирается в боковой ход и прогрызает круглое лётное отверстие, через которое выходит наружу. Генерация чаще всего двухгодовая, но может затягиваться до трех-четырех лет.
В районах севернее Москвы зимуют чаще всего яйца, изредка личинки первого возраста (Павлинов, 1965), в более южных местах личинка успевает прогрызть ход в виде полости под корой. Направление, конфигурация и размер этих ходов зависят от возраста дерева. Наиболее извилистые и крупные полости встречаются на молодых деревьях. При этом личинка часто прогрызает в коре несколько отверстий, из которых выбрасывает опилки. Позднее личинки на всех деревьях начинают вгрызаться в заболонь. При этом направление ходов может быть различным. Чаще личинки вгрызаются в тангентальном направлении, постепенно поднимаясь вверх. Однако при откладке яиц непосредственно у корневой шейки ходы сначала идут вниз. В дальнейшем личинка поднимается вверх и начинает грызть типичный сердцевинный ход. Нижняя часть хода к этому времени обычно бывает плотно забита опилками. Длина вертикального хода различна. В среднем она составляет 30 - 40 см. На старых деревьях длина ходов может достигать 1 - 1,5 м.
Взрослая личинка достигает в длину до 42 мм (в первом возрасте около 6 мм). Она беловато-желтая, безногая, в редких волосках.
Личинка перед окукливанием прогрызает боковой ход к поверхности ствола примерно в центре вертикального хода. Окукливание происходит на плотной пробке из опилок в верхней части хода. Фаза куколки около 15 дней. Отродившийся жук прогрызает круглое отверстие, которое быстро зарастает. Генерация в северной части ареала трех- и четырехгодовая (Павлинов, 1965). Для южной части ареала наряду с четырехгодовой (Петрова, 1956) указывается двухгодовая (Гречкин, 1960).
При большой плотности заселения деревьев усач вызывает их постепенное усыхание. Одновременно он наносит и технический вред, так как источенная личинками древесина полностью теряет свои технические свойства. Кроме того, от ходов личинок и надрезов жуков на ветвях интенсивно распространяется краснина.
Малый осиновый усач (Saperda populnea L.). Наносит большой вред молодым осинам и тополям. Лёт в мае - июне. Генерация двухгодовая. Поврежденные ветви и стволики, внутри которых развивается личинка, хорошо заметны по галлам, образующимся в местах откладки яиц и внедрения внутрь побега личинок (рис. 109).
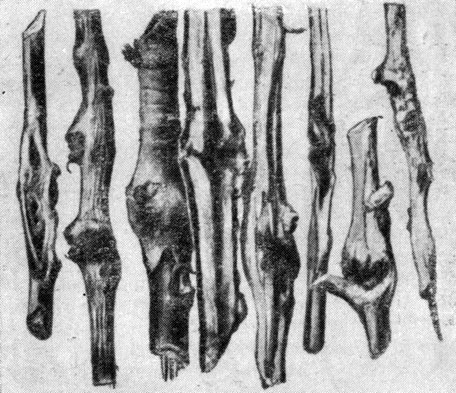
Рис. 109. Повреждения малого осинового скрипуна
Серый осиновый усач (Xylotrechus rusticus L.). Это самый распространенный и многочисленный обитатель лиственных лесов. Лёт жуков растянут, начинается в мае и продолжается до середины августа с пиком в июне. Самки откладывают яйца в щели и трещины коры стоящих ослабленных и поваленных деревьев, а также различных лесоматериалов.
Личинка сначала грызет ход под корой, глубоко задевая заболонь. Ход извилистый, иногда с расширениями и отрогами, весь забит буровой мукой. В наружных слоях древесины он обычно идет более или менее параллельно поверхности древесины, затем углубляется наискось, в тонких деревьях нередко доходя до противоположной стороны. На более толстых стволах ходы загибаются и направляются к поверхности древесины. В конце развития личинка подходит близко к поверхности древесины и там окукливается. Генерация двухгодовая, не может завершиться и в один год.
Мраморный узорчатый усач (Saperda scalaris L.). Жук имеет зеленоватую окраску с черными пятнами на надкрыльях, образующими мраморный рисунок. Длина жука 12 - 20 мм. Заселяет березу, осину, ольху и другие лиственные породы. Самка откладывает яйца в углубления, которые выгрызает в трещинах коры. Плодовитость 10 - 30 яиц (Трофимов, 1980). Личинка точит извилистый ход под корой, затем углубляется в древесину, где делает крючковатый короткий ход. Генерация одногодовая, однако ряд авторов указывают двухгодовую (Трофимов, 1980).
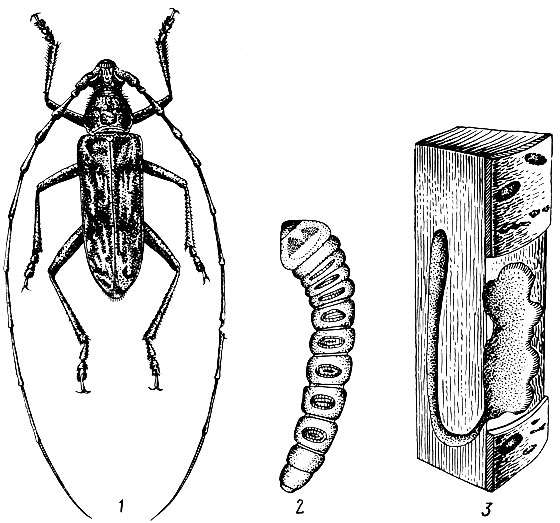
Рис. 110. Городской усач: 1 - жук, 2 - личинка, 3 - повреждение
Городской, или узбекский, усач (Aeolesthes sarta Sols.). Широко распространен в Средней Азии, где наносит огромный вред многим древесным породам, особенно произрастающим в городских условиях тополям, платану, белой акации, ореху грецкому и др. Жук длиной 28 - 47 мм, темно-серо-коричневый, с отливающим серебром волосяным покровом на надкрыльях (рис. 110). Лёт жуков с конца апреля до середины июня. Самки откладывают по одному - три яйца в щели и углубления коры стволов деревьев всех возрастов, чаще старых. Всего одна самка откладывает до 270 яиц. Отродившиеся личинки вгрызаются под кору, сначала питаются лубом, затем прокладывают извилистые ходы, сильно задевающие заболонь, и переходят в неправильно овальные полости, резко врезающиеся в луб и пробку коры. Осенью личинки углубляются в древесину и там зимуют, а на следующий год продолжают ход. Сначала он прокладывается вниз, потом резко крючкообразно изгибается и направляется вверх, параллельно поверхности ствола. Молодые жуки появляются в конце лета и зимуют в ходах. Генерация двухгодовая.
Городской усач нападает на внешне здоровые, еще вполне жизнеспособные деревья и постепенно приводит их к гибели. Он встречается в долинах и горных лесах до высоты 1800 м над уровнем моря.
Златки (Buprestidae)
Семейство златок объединяет жуков различной величины (от 3 до 80 мм), преобладающее большинство которых живет в тропических странах. В СССР фауна златок особенно разнообразна на Кавказе и в Средней Азии. В европейской части СССР встречается всего около 180 видов. В основном они связаны с древесно-кустарниковой растительностью. Многие виды - опасные вредители древесных пород, особенно в степной зоне, где они являются одними из основных стволовых вредителей.
Жуки имеют плоское, удлиненное, суженное к концу тело, металлически-блестящее, с ярко окрашенными твердыми элитрами. Голова маленькая, ноги короткие, лапки пятичлениковые, усики 11-члениковые, пильчатые. Форма тела и хорошо развитые задние крылья способствуют быстрым и дальним полётам златок и их распространению по территории.
Жуки златки исключительно свето- и теплолюбивы. Они летают, спариваются и откладывают яйца только при ярком солнечном свете, тяготеют к хорошо освещенным и прогреваемым местообитаниям.
Самки откладывают яйца в трещины и щели коры или на ее гладкую поверхность с освещенной, обычно южной части стволов деревьев. Иногда кладки яиц на стволах деревьев они заливают жидкостью, выделяемой специальными железами. Жидкость мгновенно застывает, и на стволах деревьев образуются многочисленные белые колпачки, под которыми находятся яйца (зеленая узкотелая златка и ряд других видов p. Agrilus). Небольшая группа видов златок откладывает яйца на листьях, которые затем минируются личинками (p. Trachys). Наконец, встречаются златки, самки которых откладывают яйца в землю около корней деревьев. Вылупившиеся личинки отыскивают ближайший корень и начинают им питаться, прогрызая длинные извилистые ходы по направлению к поверхности (златки p. Capnodis).
Личинки златок сильно удлиненные, неокрашенные, желтовато-белые, безногие, слепые, с характерно расширенным и уплощенным сверху и снизу переднегрудным сегментом, несущим сверху одну или две сходящиеся впереди бороздки. Голова маленькая, темная, втянутая в переднегрудь. Личинки златок сухие на ощупь и могут выносить высокие температуры под корой дерева (до 48°С), мирясь с большой сухостью субстрата и воздуха, что способствует их выживанию в межвидовой борьбе с личинками других стволовых вредителей.
По образу жизни личинки златок делятся на несколько групп. Одни из них все развитие проходят под корой, питаясь лубом и заболонью, другие заканчивают развитие в древесине, третьи - почти все время питаются в древесине.
Под корой личинки прогрызают плоские с острыми краями, извилистые, постепенно расширяющиеся ходы, плотно забитые пескообразной волнистой буровой мукой. Иногда ход несколько раз пересекается и образует в конце характерный клубок (зеленая узкотелая златка). Чаще всего ходы имеют поперечное направление и сначала проходят в коре и лубе, не задевая заболони. На хвойных породах это позволяет златкам первыми заселять деревья, так как их ходы при этом почти не нарушают систему смолоходов (синяя сосновая златка). В древесине ходы бывают короткие в виде крючка (например, у p. Chrysobothris). Технический вред древесине приносят только несколько видов златок, прокладывающих длинные ходы в старых пнях, столбах и бревнах (p. Buprestis). Ряд видов живет в корнях древесных пород, произрастающих в пустыне (джузган, гребенщик, саксаул и др.), истачивая их во всех направлениях.
Личинки обычно зимуют один-два раза и весной окукливаются в куколочковых колыбельках. Из куколок через две-три недели выходят молодые жуки. Они прогрызают лётное отверстие, имеющее форму более или менее вытянутого иногда очень узкого эллипса. Одна сторона, соответствующая спинке жука, более плоская, другая, соответствующая его брюшной поверхности, более выпуклая.
После вылета молодые жуки у многих видов проходят дополнительное питание на цветках и листьях. Генерация у златок чаще всего одно- и двухгодовая.
Среди златок преобладают вредители лиственных пород, фауна хвойных пород сравнительно бедна видами. Каждый вид предпочитает какую-нибудь одну или несколько близких между собой древесных пород и заселяет определенную часть ствола дерева, ветви или корни. Так, большинство мелких антаксий (p. Anthaxia) заселяет ветви и вершину стволов, а дицерки (p. Dicerca) поселяются в нижней части деревьев.
Многие виды златок очень активны и нападают на относительно здоровые деревья, заселяя их раньше усачей и короедов. Для своего размножения они выбирают изреженные, хорошо прогреваемые насаждения, произрастающие в ксерофильных условиях, в первую очередь опушки, кулисы, недорубы, группы семенников на лесосеках, полезащитные полосы и посадки без бокового отенения второго яруса и подлеска.
На хвойных породах самыми распространенными и вредными видами являются синяя сосновая златка, лиственничная шеститочечная, златка, четырехточечная еловая златка, златка пожарищ, ребристая бронзовая златка, арчовая златка.
Синяя сосновая златка (Phaenops cyanea F.). Жук 8 - 11 мм, с плоским удлиненным телом. Окраска нижней части тела зеленая, верхней сине-зеленая или темно-синяя с металлическим отливом (рис 111) Лёт в июне - июле. Самка откладывает яйца в трещины коры. Через три - пять дней выходят личинки. Они прогрызают длинные извилистые ходы, кольцующие дерево, и зимуют в толще коры, свернувшись в подковку.
Окукливаются личинки в мае следующего года. Фаза куколки длится 10 - 15 дней, генерация одногодовая.
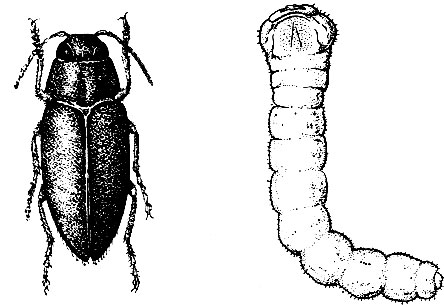
Рис. 111. Синяя сосновая златка и ее личинка
Златка первой заселяет ослабленные, но еще вполне жизнеспособные сосны с изреженной кроной и желтеющими кончиками хвоинок, в возрасте 20-80 лет. Заселение начинается с южной стороны дерева от высоты 1 - 1,5 м и захватывает всю среднюю часть ствола до места прикрепления кроны. Она предпочитает изреженные сухие сосняки. Особенно интенсивно размножаются в очагах корневой губки и на пожарищах, в сосновых культурах после двухлетней весенней засухи.
Личинки златки активно истребляются малым пестрым дятлом и пищухой, энтомофаги большой роли в колебаниях численности не играют. Примесь ели в сосняках резко снижает численность златки.
Лиственничная шеститочечная златка (Phaenops guttulata Gelb.). Жук длиной 7-11,5 мм, удлиненно-овальный, черный, с бронзовым отливом; на надкрыльях три пары светло-желтых пятен. Лёт в июне - июле. Самка откладывает яйца в трещины коры деревьев сибирской и даурской лиственниц. Зимуют личинки под корой и в мае окукливаются. Молодые жуки питаются хвоей лиственницы.
Златка поселяется на сравнительно мало ослабленных, еще жизнеспособных деревьях разного возраста. Заселяется наиболее прогреваемая часть дерева от высоты 3 - 5 м. Предпочитает изреженные насаждения, поврежденные хвоегрызущими насекомыми, гари, лесосеки. Распространена по всему естественному ареалу сибирской и даурской лиственниц.
Златка пожарищ (Melanophila acuminata Deg.). Жук длиной 6,5 - 13 мм. Одноцветно угольно-черный, удлиненный, сзади клиновидно сужен. Лёт в июне - июле. Самки откладывают яйца в трещины коры в нижней и средней части стволов. Личинки точат длинные ходы, как у предыдущих видов, зимуют и окукливаются в древесине. Генерация одногодовая. Встречается на гарях. Предпочитает средневозрастные деревья ели, реже сосны и других хвойных пород. В Сибири часто заселяет березу (Рожков, 1966). Вид очень свето- и теплолюбивый, широко распространен во всей лесной зоне.
Ребристая бронзовая златка (Chrysobothris chrysostigma L.). Распространена в лесной зоне и по образу жизни очень напоминает предыдущий вид.
Четырехточечная златка (Anthaxia quadripunctata L.). Жук длиной 4 - 7 мм, матовый, черно-бронзового цвета, на грудном щите четыре ямки. Лёт в июне, самки откладывают яйца в трещины коры средневозрастных елей, в средней и нижней части ствола, преимущественно с южной или незатененной стороны. Личинки протачивают под корой длинные, извилистые, постепенно расширяющиеся с острыми краями плоские ходы, набитые пестрой бурой мукой, и в них зимуют. Перед окукливанием они углубляются в древесину. Молодые жуки проходят дополнительное питание на желтых цветках одуванчика и других сложноцветных. Генерация одногодовая.
Распространена во всей лесной зоне, в лесостепи встречается разновидность этой златки, повреждающая сосну.
Арчовая златка (Anthaxia conradti Sem.). Жук длиной 4 - 7 мм, темно-бронзовый, широкий, плоский. Лёт в апреле - мае, часто затягивается до июля (Махновский, 1966). Самка откладывает яйца в трещины и под чешуйки коры веток на ослабленных деревьях и порубочных остатках арчи. Личинки грызут длинные, извилистые, постепенно расширяющиеся и слабо задевающие древесину ходы. Они зимуют, весной превращаются в куколок. Затем через декаду в жуков. Молодые жуки питаются на цветках одуванчика, затем шиповника. Генерация одногодовая (по Махновскому, на растущих деревьях - двухгодовая).
Арчовая златка предпочитает изреженные, хорошо освещенные места, свето- и теплолюбива, распространена по всему ареалу арчи.
На лиственных породах встречается очень много видов златок. Ниже описываются самые распространенные из них.
Зеленая узкотелая златка (Agrilus viridis L.) (рис. 112). Жук длиной 6 - 9 мм, с узким, более выпуклым снизу телом, металлически-зеленого или синего цвета. Лёт жуков в июне. Самки откладывают яйца кучками на гладкую кору стволов и ветвей. В одной кучке 7 - 11, максимум 20 яиц. Самка заливает яйца выделениями из придаточных половых желез, вследствие чего на стволах образуются выпуклые белые щитки диаметром 2 - 3,5 мм.
Выходя из яиц, личинки вгрызаются под кору и прокладывают забитые буровой мукой ходы. Каждая личинка делает самостоятельный ход, но в зависимости от состояния дерева ходы личинок или свободно расходятся в стороны, или образуют овальный клубок, расположенный вдоль ствола или ветви (рис. 113). Осенью личинки углубляются в поверхностные слои древесины и устраивают куколочные колыбельки, в которых зимуют. Весной личинки окукливаются, и вскоре появляются молодые жуки, которые проходят дополнительное питание на листьях деревьев. Генерация повсеместно одногодовая.
Очаги образуются в изреженных молодых насаждениях на бедных и сухих почвах, по южным опушкам, в полезащитных полосах ажурной конструкции и т. д.
Наибольший вред златка приносит тополям, березам и кленам, а на западе - буку. По внешнему виду и образу жизни очень похожи на зеленую узкотелую златку некоторые другие узкотелые златки.
Все узкотелые златки летают в июне, зимуют в фазе личинки, имеют однолетнюю генерацию, жуки питаются листьями деревьев, на которых живет потомство. Они заселяют молодые деревья или вершины и ветви более старых, светолюбивы и предпочитают порослевые изреженные насаждения, расстроенные рубками, "кулисы" и южные опушки, узкие полезащитные полосы ажурной конструкции, посадки без бокового отенения. Узкотелые златки - опасные физиологические вредители степных лесов, особенно в годы засух, а меры борьбы с ними довольно затруднены.
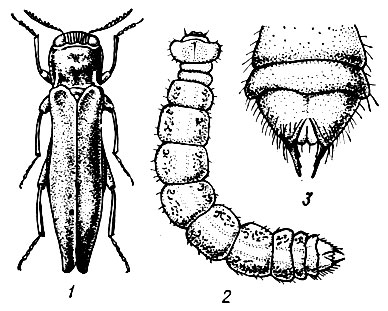
Рис. 112. Зеленая узкотелая златка: 1 - жук, 2 - личинка, 3 - анальный сегмент личинки
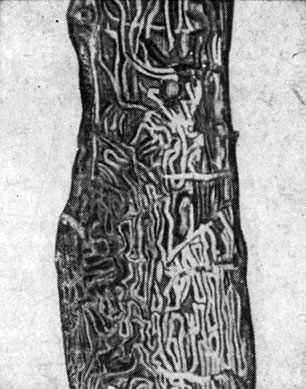
Рис. 113. Ходы зеленой узкотелой златки
Узкотелые златки различаются по откладке яиц. Подобно зеленой узкотелой златке, яйца под колпачок откладывает грабовая узкотелая златка (Agrilus olivicolor Ksw.), широко распространенная в районах произрастания граба, березовая узкотелая златка (A. betuleti Rtrb.), наносящая вред молодым березам в полезащитных полосах лесостепной зоны, и др.
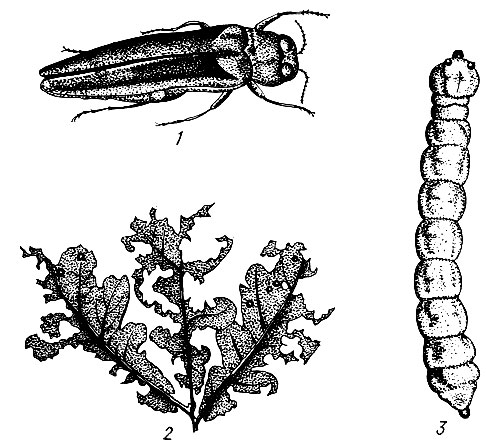
Рис. 114. Дубовая двупятнистая узкотелая златка: 1 - жук, 2 - листья дуба, поврежденные жуками во время дополнительного питания, 3 - личинка
В отличие от этих видов узкотелые златки, поселяющиеся на дубе, откладывают яйца в трещинах коры по одному, обычно на недалеком расстоянии друг от друга. Они являются опаснейшими вредителями дубрав в степной и лесостепной зонах в сухие климатические периоды. Широко распространены в дубравах шелковистая узкотелая златка (Agrilus hastulifer Rtrb.), вершинная дубовая узкотелая златка (A. angustulus 111.), двупятнистая узкотелая дубовая златка (A. biguttatus F.), удлиненная узкотелая златка (A. sulcicollis Lac.) и малая дубовая узкотелая златка (A. obscuricollis Ksw.).
Эти златки предпочитают молодые и средневозрастные порослевые дубы, а на старых заселяют только толстые ветви и вершину ствола. Исключение составляет двупятнистая златка, личинки которой живут под толстой корой свежих пней и в комлевой части живых старых дубов не выше 2 - 5 м.
Все они имеют одногодовую генерацию (только у двупятнистой златки в северных районах двухгодовая), питаются в фазе жука листьями дуба (рис. 114), хорошо летают и отличаются исключительным свето- и теплолюбием.
Бронзовая дубовая златка (Chrysobothris affinis Fobr.) менее активна, чем узкотелые златки, заселяет уже сильно ослабленные дубы, преимущественно ветровал, бурелом и лесопродукцию совместно с пестрыми усачами. Летает в июне. Откладывает яйца в трещины толстой коры. Личинки точат длинные продольного направления ходы, забитые буровой мукой, и затем углубляются в поверхностные слои древесины, где зимуют, а весной окукливаются. Генерация одногодовая, в лесной зоне бывает двухгодовая.
Осиновая златка (Poecilonota variolosa Payk.). Повреждает осину и тополя на юго-востоке. Лёт в мае - июне, генерация двухгодовая.
Тополевая пятнистая златка (Melanophila picta Pall.).
Опасный вредитель тополей в Средней Азии, Казахстане, на Кавказе и юго-востоке европейской части СССР (рис. 115). Лет жуков в мае - июне (в южных районах Средней Азии с конца апреля). Во время дополнительного питания они объедают края листьев, черешки и молодые побеги. Самка отладывает яйца в щели и углубления коры по одному, реже два-три в одно место. Развитие яйца длится 8 - 10 дней. Личинки прокладывают под корой извилистые ходы, заполненные буровой мукой (длина ходов 12 - 15 см). На тонких стволиках ходы собраны в клубки. Осенью личинки уходят неглубоко в древесину, устраивают куколочную колыбельку и в ней зимуют, а весной окукливаются. Генерация одногодовая.
Рис. 115. Тополевая пятнистая златка
Вид очень пластичный, свето- и теплолюбивый, активный. Встречается везде, где произрастают тополя, нападает на деревья всех возрастов, ветровал, лесопродукцию и пни. Особенно сильно вредит молодым тополям на плантациях и посадках, повреждает нижнюю часть стволиков и черенки.
Меры борьбы: в насаждениях, зараженных златкой, следует проводить санитарные рубки и вырубки свежезаселенных деревьев с последующей их химической обработкой. При производстве культур - обработка черенков жижей с гексахлораном, на плантациях обмазка и опрыскивание стволиков 2 - 4%-ной рабочей эмульсией 16%-ного концентрата гамма-изомера гексахлорана.
В условиях Средней Азии большую роль играют агротехнические мероприятия (рыхление почвы, уход за черенками), резко снижающие вредную деятельность златки.
Целый ряд видов Capnodis - серьезные вредители древесных пород (особенно плодовых) в Средней Азии, Казахстане, степной зоне РСФСР, на Кавказе и в Крыму. Эти крупные златки летают с апреля почти все лето. Яйца откладываются в почву около корней или прямо на корни и у основания стволов. Личинки повреждают корни и нижнюю часть стволов, вытачивая длинные широкие ходы, часто длиной до 2 м. Они живут два-три года. Генерация двух- и четырехгодовая. Жуки зимуют под опавшей листвой и в кучах мусора. Во время дополнительного питания наносят большой вред, обгладывая черенки и побеги; листья осыпаются на почву, часто покрывая ее сплошным слоем.
Слоники-смолевки (Pissodes)
Слоники-смолевки (Pissodes) имеют округленную головотрубку, примерно в середине которой прикреплены усики. Она длиной с переднеспинку, мало изогнута; усиковая бороздка прямая, идет к нижнему краю глаз. Плечи надкрылий не выступают, голени на конце с крючком. Щиток круглый, большой, в светлых чешуйках. Надкрылья в пятнах, образуемых чешуйками. Личинки белые с желто-бурой головкой, слепые, безногие, изогнутые. К этому роду относится несколько видов жуков, которые наносят большой вред хвойным насаждениям в возрасте 15 - 40 лет, а иногда и старше. Они тонко реагируют на малейшее ослабление дерева и поселяются на различных частях ствола. Самка откладывает яйца в кору по несколько штук. Личинки грызут под корой извивающиеся и постепенно расширяющиеся в разные стороны от места откладки яиц ходы. На тонких стволиках направление ходов продольное, а на более толстых они расходятся звездообразно. Типичный ход смолевки изображен на рис. 116.
Личинки окукливаются в заболони, где делают продолговатое углубление (колыбельку), которое закрывают мелкими стружками. Окукливание обычно происходит в конце лета и вскоре из куколки выходит жук, прогрызающий очерченную круглую лётную дыру. Жуки обычно зимуют в лесной подстилке и под корой старых пней, а весной приступают к размножению. Они проходят дополнительное питание лубом в области тонкой коры деревьев или на молодых побегах и ветвях. Генерация у всех видов одногодовая.
В предыдущих главах были рассмотрены слоники-смолевки, вредящие шишкам (Pissodes validirostris) и молодым культурам сосны (Pissodes notatus).
Более взрослые насаждения повреждают следующие виды.
Сосновая вершинная смолевка (Pissodes piniphilus Hrbst.). Распространена в чистых сосновых насаждениях 15 - 40 лет. Жуки летают в июне - июле и откладывают яйца по 1 - 5 шт. под тонкой корой в верхней части сосен. Личинки прокладывают между корой и лубом извилистые, постепенно расширяющиеся ходы. На более толстых деревьях ходы образуют звездообразную фигуру. Личинки зимуют и весной окукливаются в заболони. Нападает на еще жизнеспособные сосны, вызывая их отмирание. Очаги возникают в местах массового снеговала и снеголома, при интенсивном изреживании густых насаждений.
Стволовая смолевка (Pissodes pini L.). Поселяется в области переходной коры средневозрастных сосен (рис. 116). Образ жизни такой же, как у предыдущего вида. Очень распространенный вид, сопутствует синей златке, сосновым лубоедам.
Еловая смолевка (Pissodes harcyniae Hrbst.). Нападает на внешне здоровые деревья и является неизменным спутником корневых заболеваний деревьев (опенок, корневая губка). Лёт жуков и кладка яиц очень растянуты (с конца мая до июля). Личинки образуют типичные извилистые звездообразные ходы, оканчивающиеся куколочными колыбельками. Окукливание осенью или весной. Молодые жуки дополнительно питаются в верхней части стволов, вызывают смолотечение и сильно ослабляют деревья.
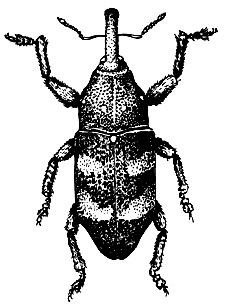
Стволовая смолевка
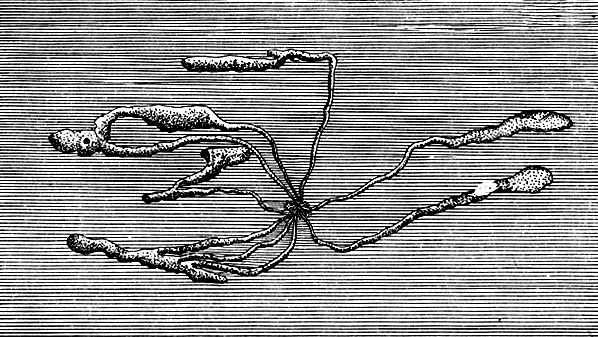
Рис. 116. Стволовая смолевка и ее повреждения
Пихтовая смолевка (Pissodes piceae 111.). Повреждает кавказскую и белую пихту. Лёт в мае, начале июня, растянут. Откладка яиц в местах с поврежденной корой. Зимуют личинки и жуки. Предпочитает область толстой коры. Дополнительное питание жуки проходят на подросте пихты, выедая площадки в коре.
Лиственничная смолевка (Pissodes insignitus Boh.). Повреждает все виды лиственниц в Сибири. Генерация одногодовая.
Перепончатокрылые (Hymenoptera)
Рогохвосты (Siricidae)
Рогохвосты имеют длинное цилиндрическое тело, сзади заостренное, у самок с выдающимся, иногда длинным яйцекладом. Личинки беловатые, цилиндрические, слегка S-образно изогнутые, сплюснутые с брюшной стороны, с тремя парами рудиментарных грудных ног и с острым ступенчатым отростком на заднем конце брюшка. При помощи яйцеклада самка просверливает кору и откладывает яйца по 1 - 3 шт. в одно место полосой вдоль ствола. Личинки проделывают в древесине круглые в поперечном разрезе ходы, туго забитые пылевидной буровой мукой. Поврежденные рогохвостом деревья обнаруживаются по круглым, типичным летным отверстиям. Летают рогохвосты с июня до сентября. Генерация одно- и двухгодовая. Дополнительного питания не проходят.
Все рогохвосты приносят технический вред; кроме того, многие виды очень активны и могут нападать на внешне здоровые деревья, выбирая стволы с механическими повреждениями. Скрытый образ жизни делает их малодоступными для изучения и потому биология ряда видов известна еще недостаточно.
Самые распространенные виды на хвойных породах - большой хвойный (Urocerus gigas L.), синий (Sirex juvencus F.), фиолетовый (S. noctilio F.), черно-синий (S. ermak Sem.) и черный (Xeris spectrum L.) рогохвосты.
Большой хвойный рогохвост (рис. 117) повреждает ель и сосну, а в лесах Сибири - ель, пихту, сибирский кедр и лиственницу. Генерация двухгодовая, на юге может быть одногодовая. Вид экологически пластичный, поселяется на ослабленных деревьях в комплексе с усачами и златками.
Синий, фиолетовый и черный рогохвосты широко распространены в лесах СССР. Они повреждают сосну, ель, пихту, реже лиственницу. Лёт в середине лета, генерация одно- и двухгодовая.
Черно-синий рогохвост - типичный обитатель сибирской тайги. Лёт в июле - августе, генерация двухгодовая. Повреждает все хвойные, гигрофил (Строганова, 1968).
На лиственных породах биология рогохвостов изучена недостаточно. Поэтому часто недооценивается их значение как активных стволовых и технических вредителей. Наиболее известен березовый рогохвост (Tremex fuscicornis L.). Это самый крупный вид, живущий на лиственных породах (длина тела 30 - 40 мм). Он летает в августе - сентябре, личинка проделывает в древесине сложные ходы и окукливается недалеко от поверхности. Генерация двухгодовая. Часто заселяет березу совместно с зеленой узкотелой златкой. Вид экологически пластичный, его очаги встречаются в полезащитных лесополосах, в заболачивающихся березняках, в котловинах выдувания песков и т. д. Кроме березы повреждает изредка иву, осину и ильмовые.
Лиственные породы повреждаются также рядом ксифидрий (Xiphydriidae) - перепончатокрылых насекомых, очень близких к рогохвостам и обычно носящих это же название. Среди них особенно распространены ольховая и дубовая ксифидрий.
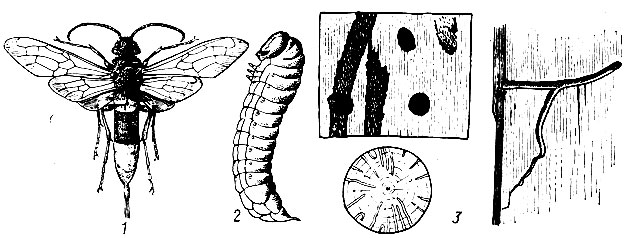
Рис. 117. Рогохвост гигант: 1 - имаго, 2 - личинка, 3 - повреждение
Ольховая ксифидрия (Xiphydria camelus L.). Повреждает ряд лиственных пород, но особенно сильно вредит средневозрастным насаждениям ольхи, образуя часто большие очаги. Лёт в июне - июле, генерация одногодовая.
Дубовая ксифидрия (X. longicoltis Geoffr.). Повреждает дуб; образ жизни, как у предыдущего вида.
Чешуекрылые (Lepidoptera)
Стволы деревьев повреждают бабочки, относящиеся к семействам древоточцев и стеклянниц.
Древоточцы (Cossidae)
Крупные густоволосистые бабочки, ведущие ночной образ жизни.
Древесница въедливая (Zeuzera pyrina L.). Бабочка в размахе крыльев 40 - 70 мм, атласно-белая с многочисленными угловатыми синевато-черными пятнами (рис. 118). Лёт бабочек начинается во второй половине июня и продолжается до середины августа. Бабочки мало подвижны (в особенности самки), почти не летают и не питаются. Самок в природе обычно бывает больше, чем самцов. Самки откладывают яйца эллиптической формы, сначала желтого, затем оранжевого цвета по одному на верхушки молодых побегов, в пазухи листьев, на листовые рубцы и почки. Плодовитость одной самки в среднем 1000 яиц, иногда 1140 и даже 2280 яиц. Развитие гусеницы в яйце длится 12 - 15 дней (Анфинников, 1961).
После выхода из яйца молодая гусеница вбуравливается в черешок листа, отчего поврежденные листья засыхают и преждевременно опадают. Через 7 - 10 дней молодые гусеницы покидают листья, добираются до побегов последнего года, проникают внутрь них и протачивают ходы, питаясь сердцевиной. До наступления низких температур гусеницы успевают перелинять и переселиться в побеги предыдущих лет, где закупориваются червоточиной и зимуют. На второй год кроме вертикальных ходов гусеницы прокладывают ходы горизонтального направления, которые перерезывают сосуды и ослабляют дерево. По мере роста гусеницы продолжают менять свои ходы и опускаются все ниже и ниже по дереву. Осенью они закупориваются в ходах, проложенных в средней и нижней частях дерева, и вторично зимуют. Весной третьего календарного года гусеницы уже не меняют хода, а только расширяют в нем входное отверстие и заканчивают развитие. Ход состоит из довольно широкой неправильной формы полости между древесиной и корой, с отверстием в последней, и отходящего вверх канала, сначала изгибающегося, а затем прямого, длиной до 15 - 20 см.
Рис. 118. Древесница въедливая: 1 - яйца, 2 - гусеница, 3 - куколка, 4 - бабочка, 5 - повреждение
Взрослая гусеница 16-ногая желтовато-белая, на каждом сегменте тела по ряду черных точек, несущих по одному волоску. Голова большая темно-бурая, длина тела 50 - 60 мм.
В конце мая - июне гусеница окукливается без кокона в верхней части хода, где зимовала. Куколка желтовато-бурая. На голове между глазами короткий рог, обращенный вершиной вперед. Ее развитие длится 6 - 10 дней. Перед выходом бабочки куколка спускается вниз и высовывается наполовину из входного отверстия.
Генерация древесницы двухгодовая, с четко выраженными лётными годами. Они чаще всего приходятся на нечетные годы. В городских насаждениях четко выраженных лётных годов часто не бывает.
Древесница въедливая многоядна, она повреждает более 70 древесных пород, половина из которых широко распространена в насаждениях европейской части СССР. Наиболее повреждаются ясени, затем ильмовые, реже дуб; в ряде городов сильно заражены рябина, ясенелистный клен, липа, белая акация, из плодовых - яблоня и груша. Из ясеней сильнее всех повреждается ясень обыкновенный, а меньше других - ясень зеленый.
Интенсивность заражения древесных пород древесницей находится в обратной зависимости от энергии их роста. Поэтому особенно сильно заражаются посадки и отдельные деревья, характеризующиеся плохим ростом, отсутствием ухода, имеющие механические повреждения. В лесных условиях интенсивность заражения резко возрастает после кульминации текущего прироста (к 15 - 25 годам).
Древесница предпочитает освещенные и прогреваемые участки, поэтому зараженность возрастает по направлению к южным и западным опушкам, на широких улицах, при редком стоянии деревьев. Меньше всех повреждаются посадки с хорошим боковым отенением, густые, с наличием второго яруса или созданные по древесно-кустарниковому типу. Насаждения и деревья порослевого происхождения всегда повреждаются больше, чем возникшие из семян. Древесница распространяется с посадочным материалом, иногда с древесиной.
Гусеницы древесницы истребляются дятлами, а яйца - большой синицей. Энтомофаги большого значения в колебаниях численности древесницы не имеют. Очаги ее массового размножения в степных лесах юго-востока европейской части СССР распространены на больших площадях и довольно устойчивы.
Меры борьбы проводятся в комплексе. Для ликвидации очагов массового распространения древесницы и выращивания здоровых насаждений необходимо выполнять комплекс мероприятий. Он состоит из санитарных рубок, мероприятий по созданию новых устойчивых насаждений, карантина и химических мер борьбы.
Выборка отдельных деревьев, заселенных древесницей въедливой, должна производиться в слабо зараженных насаждениях, чтобы достигнуть локализации возникающих очагов. Выборочные санитарные рубки проводятся в насаждениях со средней степенью заселенности при условии их нормальной сомкнутости. Под сплошные санитарные рубки отводятся насаждения с полнотой не выше 0,6 и со степенью зараженности не менее 50%. Эти рубки необходимо увязывать с лётными годами и сроками развития древесницы.
Все рубки должны сопровождаться тщательным уничтожением мелких ветвей, которые могут быть в это время заселены гусеницами древесницы. Рубки ухода в зараженных древесницей насаждениях рекомендуется производить один раз в четыре года, в лётные годы, соблюдая при этом те же условия, что и при санитарных рубках.
При создании новых устойчивых против древесницы насаждений рекомендуется:
насаждения с участием ясеня создавать по древесно-кустарниковому типу с введением ясеня не более 10% состава, снижая это количество в худших условиях роста, вплоть до полного исключения из состава; ясень обыкновенный и пушистый лучше заменять ясенем зеленым, а берест и вяз - вязом мелколистным;
новые посадки в непосредственной близости от зараженных насаждений производить без ясеня, ввода в состав насаждений дуб, полевой клен и другие устойчивые породы.
Посадочный материал в питомниках перед вывозом необходимо проверять и в случае обнаружения зараженных гусеницами древесницы саженцев немедленно уничтожать их. Питомники закладывать не ближе 500 м от зараженных насаждений, особенно ясеневых.
Во всех насаждениях, где имеется возможность производить индивидуальный уход за деревьями, целесообразно применять дихлорэтан и гексахлоран для введения в окончательные ходы древесницы, обычно располагающиеся в нижней части стволов. Химикаты вводятся в ходы с помощью резиновой груши с изогнутым наконечником или с помощью влажных тампонов. Отверстия ходов для более эффективного действия химикатов необходимо замазывать глиной или еще лучше цементировать. Нормы расхода - 0,5 г на один ход. Вводить химикаты в отверстия ходов следует в августе - сентябре межлётного года или в мае лётного года, когда зараженные деревья хорошо заметны по скоплению кала возле основания стволов и легко различимы свежие действующие ходы.
На больших площадях можно применить авиационно-химическое опрыскивание в период лёта бабочек и против молодых гусениц с помощью водных растворов концентрированных эмульсий гексахлорана.
Древоточец пахучий (Cossus cossus L.). Бабочка в размахе крыльев 80 - 85 мм. Обе пары крыльев коричневато-серые, испещренные многочисленными поперечными черными полосами. Усики гребенчатые (рис. 119).
Лёт бабочек в лесостепи начинается со второй декады июня и продолжается около двух недель. В лесной зоне он более растянут. Бабочки летают в вечерние часы. Погодные условия не оказывают существенного влияния на их лёт. Самка откладывает яйца в трещины коры кучками по 20 - 70 шт. (колебания 4 - 228 шт.). Плодовитость самки около 1000 яиц (колебания 237-1350 шт.). Яйца откладываются самкой в основном в первые три-четыре дня. Отложенные в последние дни яйца мелкие, весят на 30% меньше, чем отложенные в первый день (Насонова, 1960). Фаза яйца длится 10 - 12 дней. Первые два-три дня гусеницы сидят под оболочками яиц, потом вгрызаются под кору и все вместе грызут общий поверхностный ход неправильной формы. Гусеницы выбрасывают кал красно-бурого цвета, по которому легко обнаружить заражение.
Гусеницы 16-ногие, длиной 100 - 120 мм, с черными бляшками на теле, несущими волоски; голова темно-бурая, блестящая. Цвет тела гусениц меняется на протяжении жизни. Только что отродившиеся гусеницы розовые, затем они становятся темно-бордового цвета, а перед окукливанием вновь меняют окраску на розовую и наконец приобретают кремовый цвет.
В первый год развития гусеницы успевают слинять четыре-пять раз. Они зимуют в семейных ходах, а на следующий год расходятся и порознь втачиваются в древесину, где проделывают широкие, преимущественно продольные ходы. Всего гусеницы имеют восемь возрастов, и развитие их продолжается 22 месяца. Осенью второго года многие гусеницы бросают ходы и ползают в поисках места для окукливания.
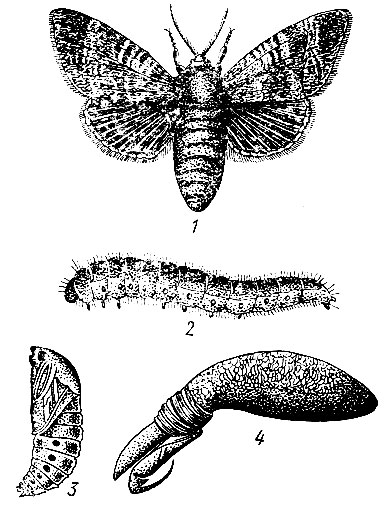
Рис. 119. Древоточец пахучий: 1 - бабочка, 2 - гусеница, 3 - куколка, 4 - кокон
Окукливание происходит на третий год во второй декаде мая - начале июня. Гусеницы окукливаются в плотном шелковистом коконе в почке, старых пнях и у основания стволов деревьев, в которых жили. Фаза куколки длится около месяца. Генерация двухгодовая.
Древоточец заселяет преимущественно нижнюю часть стволов деревьев различных лиственных и плодовых пород: ив, тополей, ольхи, вяза и дуба. Часто образуются небольшие, но устойчивые многолетние очаги. Заселение легко узнать по опилкам, вытекающему из отверстий соку и сильному запаху древесного уксуса.
Меры борьбы. Санитарные рубки и вырубка заселенных деревьев осенью лётного года, когда гусеницы зимуют под корой в общих ходах. В садах и городских посадках можно вводить в ходы гусениц эмульсию гексахлорана и замазывать ходы глиной.
Древоточец осиновый (Cossus terebra F.). Бабочки похожи на пахучего древоточца, но общий тон окраски более серый, а не светло-коричневый, как у предыдущего вида. Усики гребенчатые. Образ жизни этого вида очень напоминает таковой у предыдущего вида, но самки откладывают яйца разбросанно, по два-три яйца в одно место, и не прикрывают их бурой затвердевающей жидкостью. Гусеницы никогда не выползают из своих ходов, окукливаются в дереве, где проходило их развитие, не делают кокона (Золотаренко, 1959). Генерация точно не установлена. Этот вид повреждает только осину и тополя и, видимо, имеет широкое распространение, но его гусеницы часто принимаются за гусениц ивового древоточца.
В Средней Азии широкое распространение имеет тамариксовый древоточец (Holcocerus arenicola Stqr.). Гусеницы живут в нижней части стволов и корнях тамарикса, саксаула и других Древесных растений, произрастающих в пустыне и тугайных лесах. На тамариксе биология этого вида сходна с таковой у других древоточцев. Встречаются очаги спорадически, но в отдельных случаях может наносить тамариксу большой вред (Синадский, 1960).
Стеклянницы (Aegeriidae)
Небольшие бабочки с узкими прозрачными крыльями, напоминающие перепончатокрылых насекомых. Задние крылья короче передних, чешуйки сосредоточены на жилках. Тело довольно стройное, брюшко длинное, далеко выдается за крылья, усики веретеновидные. Летают днем. Гусеницы беловатые, 16-ногие, с бурой головой и редкими, правильно расположенными по кольцам тела волосками. Большинство видов живет в древесине деревьев, часто нанося им большой вред. Наибольшее распространение имеют темно-крылая и большая тополевая стеклянницы.
Темнокрылая стеклянница (Paranthrene tabaniformis Rtt.). Caмый распространенный и опасный вредитель тополей в насаждениях большинства городов. Бабочка в размахе крыльев 24 - 28 мм, синевато-черная, блестящая, на сегменте брюшка узкие желтые кольца. Передние крылья кофейно-бурые, а у основания прозрачные с несколько более темной бахромой. Задние крылья прозрачные, стекловидные (рис. 120).
Лёт начинается в средней полосе с конца июня и в июле, а на юге в конце мая. Самки откладывают овально-вытянутые, смоляно-черного цвета яйца, по одному или реже сразу по несколько штук на ветви и стволы в местах различных повреждений. Плодовитость одной самки 200 - 600 яиц. Развитие гусеницы в яйце 12 - 13 дней, а при высоких температурах (до +30° С) сокращается в два-три раза. Вышедшие из яиц гусеницы вгрызаются под кору, где делают отдельные площадки (полости), а затем углубляются в древесину до 4 см и прокладывают в ней продольные ходы длиной до 15 - 24 мм. Характерным признаком заселения деревьев стеклянницей являются кучки бурых экскрементов и буровой муки на стволах в местах отверстий и у основания деревьев.
Молодые гусеницы беловато-розовые, а взрослые белые или желтоватые. Голова и затылочный щиток коричнево-бурые, на последнем брюшном сегменте имеются два коричневых шипика. Длина тела 22 - 24 мм. Гусеницы пять раз линяют и имеют шесть возрастов. Они живут два календарных года - первый год зимуют в третьем возрасте в полостях под корой, а второй раз - в шестом возрасте в ходах в древесине. Перед окукливанием на третий календарный год весной они делают ниже верхнего конца хода в древесине боковой лётный ход до поверхности коры. Затем гусеница окукливается в верхнем конце хода в древесине в желтоватом коконе. Место окукливания отгораживается гусеницей от остального хода пробкой из опилок и паутины. Фаза куколки длится 12 - 14 дней. Перед выходом бабочки куколка при помощи шипинок брюшка продвигается по ходу, раздвигает тонкий слой коры и высовывается наружу примерно на 2/3 своей длины.
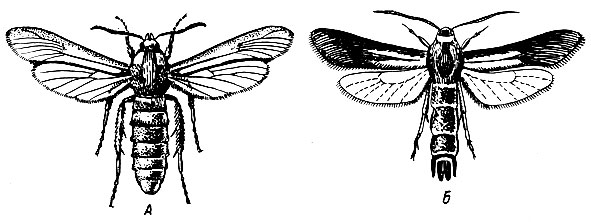
Рис. 120. Стеклянницы. А - тополевая; Б - пятнистая
Куколка темно-желтая или красно-бурая, становящаяся почти черной перед выходом бабочки. Длина 15 - 20 мм. Генерация двухгодовая. Стеклянница заселяет деревья всех возрастов, в том числе порослевые побеги уже со второго года их роста толщиной от 0,7 см и выше. На молодых побегах, стволиках и ветвях в местах поселения вредителя образуются галлообразные или односторонние вздутия. На деревьях от 10 лет и старше стеклянница может заселить не только нижнюю часть, а весь ствол, образуя на всем его протяжении наросты с вытекающим бурым соком. Заселяя пни, она препятствует развитию поросли. Через ходы гусениц деревья могут заражаться грибными и бактериальными заболеваниями, а внутри древесины возникает краснина.
Ходы стеклянниц часто раздалбливают дятлы, а на гусеницах паразитируют наездники. Однако, несмотря на значительную смертность во всех фазах развития, численность стеклянницы в тополевых насаждениях большинства городов РСФСР очень высокая и массовые повреждения и гибель тополей требуют систематических и усиленных мер борьбы с ней.
Большая тополевая стеклянница (Aegeria apiformis Cl.) (рис. 120). Бабочка в размахе крыльев 35 - 45 мм, черно-бурая с лимонно-желтыми пятнами и полосами, прозрачными крыльями. По внешнему виду напоминает осу. Лёт начинается в средней полосе в июле, на юге - в июне и длится около месяца. Самки откладывают овально-уплощенные, бурого цвета яйца по одному или небольшими кучками на нижнюю часть стволов деревьев, на корни и почву. Плодовитость одной самки 1000-1300 яиц; иногда достигает 2500 яиц. Развитие гусеницы в яйце длится две-три недели.
Вышедшие из яиц гусеницы втачиваются под кору корней нижней части стволов деревьев, где выгрызают сначала небольшие площадки, а затем углубляются в заболонь и проделывают желобчатые ходы, забитые опилками. В комлевой части стволов и в толстых корнях ходы бывают неправильные, часто сливающиеся в площадки, а на корнях - продольные, иногда с углублением в почву на 20 - 30 см. Молодые гусеницы бледно-розовые, взрослые - белые или слегка желтоватые. Голова красно-бурая. На тергите последнего сегмента брюшка имеется небольшая бородавка с малозаметным хитинизированным щитком, наклоненным в сторону головы. Длина тела до 55 мм.
Гусеницы семь раз линяют и проходят восемь возрастов. Они обычно живут два календарных года, зимуют в ходах и на третий год весной окукливаются в колыбельке под корой у комлевой шейки в плотном коконе из опилок и экскрементов; нередко - в почве близ корней. Фаза куколки длится 20 - 25 дней. Куколка коричневая или красно-бурая с рядом шипиков на спинной стороне брюшка.. Голова и переднеспинка с общим продольным килеобразным углублением на 2/3 высовывается из лётного отверстия. После вылета бабочки шкурка куколки остается торчать в круглом лётном отверстии. По этому признаку и выступающей из-под коры крупной буровой муке легко определить заселенные деревья.
Меры борьбы. Со стеклянницами меры борьбы носят главным образом профилактический характер. При создании культур тополя черенками или саженцами необходимо тщательно отбраковывать посадочный материал, не допуская к использованию заселенные темнокрылой стеклянницей экземпляры, для чего необходимо отбраковывать экземпляры со вздутиями и с опилками. При создании тополевых культур желательно затенение из кустарников, которые препятствуют заселению. Рекомендуется введение бузины. На плантациях и городских насаждениях необходим ремонт посадок, замена тополей другими породами, подбор устойчивых видов тополей и т. д. Для уничтожения выходящих бабочек в конце мая - июне обмазывают участки ствола деревьев пастой следующего состава: 40 частей навоза, 50 частей глины и 10 частей воды. В питомниках, при ценных посадках - вырезка и уничтожение заселенных частей растений, раздавливание проволокой гусениц в ходах, замазывание поврежденных мест цементом и т. п. Химическая обработка ценных культур системными инсектицидами (рогором, фосфамидом, Би-58), а также хлорофосом в 0,5 - 1%-ной концентрации в период отрождения личинок и их питания в поверхностных слоях коры до заглубления в древесину. Возможна также защитная обработка деревьев и пней перед лётом вредителей 3-5%-ной эмульсией гамма-изомера ГХЦГ (по 16%-ному препарату). При заселении до 30 деревьев - выборочная химобработка, при более высокой заселенности - сплошная обработка насаждений. Расход рабочей жидкости при подеревной обработке 0,2 - 0,5 л на одно дерево, механизированной сплошной- 100 - 300 л/га. Выборка свежезаселенных усыхающих деревьев.
Литература
Анфинников М. А. Древесница въедливая и борьба с ней. Киев, 1961.
Бородин А. Л. Подход к изучению популяционной экологии стволовых вредителей.- Зоолог, журн. т 55, вып. 2. М., 1976.
Бородин А. Л., Кирста Л. В. Количественный учет малого соснового лубоеда.- Науч. тр. Моск. лесотехн. ин-та, вып. 65. М., 1974.
Гайченя И. А., Сериков О. Я., Фасулати К. К. Стовбурни шкидники лису (атлас-справочник). Киев, 1970.
Галасьева Т. В. Таблицы выживаемости большого соснового лубоеда на гарях в Московской области.- Тр. Моск. лесотехн. ин-та, вып. 90. М., 1976.
Головянко 3. С. К методике учета зараженности сосен короедами.- Тр. по лесн. опытн. делу Украины, т. 4. Киев, 1926.
Гурьянова Т. М. Биология пихтовой смолевки.- Сб. работ Моск. лесотехн. ин-та, вып. 26. М., 1969.
Гурьянова Т. М. Роль паразитов в снижении численности пихтовой смолевки Pissodes piceae (Curculionidae).- Зоолог, журн., вып. 4. М., 1969.
Земкова Р. И. Стволовые вредители темнохвойных лесов Западного Саяна. Красноярск, 1965.
Ильинский А. И. Закономерности в размножении малого соснового лубоеда и теоретическое обоснование мер борьбы с ним в лесу.- Защита растений, т. 5, 1928, № 5-6.
Ильинский А. И. Вторичные вредители сосны и ели и меры борьбы с ними.- Сб. работ по лесн. хоз-ву Всес. ин-та лес-ва и механиз., вып. 36. М., 1958.
Исаев А. С. Стволовые вредители лиственницы даурской. М, 1966.
Исаев А. С, Гирс Г. И. Взаимодействие дерева и насекомых-ксилофагов. Новосибирск, 1975.
Исаев А. С, Петренко Е. С. Биогеоценотические особенности динамики численности стволовых вредителей.- Лесоведение, 1968, № 3.
Катаев О. А., Голутвин Г. И. Некоторые особенности сосновых и еловых насаждений как среды размножения стволовых насекомых.- Лесоведение, 1979, №6.
Киселев В. В. Моделирование динамики численности ксилофагов на примере большого лиственничного короеда.- В кн.: Закономерности распространения и динамика численности лесных насекомых. Красноярск, 1978.
Коломиец Н. Г., Богданова Д. А. Фенология короеда-дендроктона на юге Западной Сибири.-Изв. СО АН СССР, сер. биол., вып. 2. Новосибирск, 1978.
Костин И. А. Стволовые вредители хвойных пород Казахстана. Алма-Ата, 1964.
Костин И. А. Жуки-дендрофаги Казахстана. Алма-Ата, 1973.
Коротнев Н. И. Короеды. М., 1926.
Криволуцкая Г. О. Короеды острова Сахалин. М., 1958.
Криволуцкая Г. О. Скрытностволовые вредители в темнохвойных лесах Западной Сибири. М.- Л., 1965.
Куренцов А. И. Короеды Дальнего Востока СССР. М.- Л., 1941.
Куренцов А. И. Вредные насекомые хвойных пород Приморского края.- Тр. Дальневост. Фил. АН СССР, сер. зоол., т. 1 (4). Владивосток, 1950.
Кутеев Ф. С. Узкотелые златки - вредители дуба.- Лесное хозяйство, 1972, №4,
Линдеман Г. В. Заселение стволовыми вредителями лиственных пород в дубравах лесостепи в связи с их ослаблением и отмиранием.- В кн.: Защита леса от вредных насекомых. М., 1964.
Линдеман Г. В. Пути приспособления короедов-заболонников (Coleoptera, Scolytidae) к обитанию на мало ослабленных деревьях.- Экология, 1978, № 6.
Лозовой Д. А. Вредные насекомые парковых и лесопарковых насаждений Грузии. Тбилиси, 1965.
Лурье М. А. Группировки стволовых вредителей ели в южной подзоне тайги европейской части СССР.- Зоолог, журн., т. 44, вып. 10. М., 1965.
Мамаев Б. М. Биология насекомых-разрушителей древесины.- Всес. ин-т научн. и техн. информации, сер. Итоги науки и техники, т. 3,- Энтомология, М., 1977.
Маслов А. Д. Вредители ильмовых пород и меры борьбы с ними. М., 1970.
Маслов А. Д., Кутеев Ф. С, Прибылова М. В. Стволовые вредители леса. М., 1973.
Махновский И. К. Вредители горных лесов и борьба с ними. М., 1966.
Мозолевская Е. Г. Особенности освоения кормовых ресурсов насекомыми-ксилофагами.- Лесоведение, 1979, № 6.
Озолс Г. Э., Бичевкис М. Я. Исследование аттрактантов короеда-типографа Ips typographic (Col. Ipidae) в Латвийской ССР.- В кн.: Защита хвойных в Латвийской ССР. Рига, 1976.
Огибин Б. Н. О регуляции плотности популяции Ips typographies (Coleoptera, Ipidae) на предимагинальных стадиях развития. - Зоолог, журн., т. 53, вып. 1. М., 1974.
Павлинов Н. П. Большая тополевая стеклянница - вредитель культур Подмосковья.- В кн.: Защита леса от вредителей. Пушкино, 1963.
Павлинов Н. П. Большой осиновый усач и меры борьбы с ним.- В кн.: Защита лесных насаждений от вредителей и болезней. М., 1965.
Павлинов Н. П. Малый осиновый усач и меры борьбы с ним.- В кн.: Защита леса от вредителей и болезней. М., 1968.
Петренко Е. С. Насекомые - вредители лесов Якутии. М., 1965.
Плавильщиков Н. Н. Жуки-дровосеки - вредители древесины. М.- Л. Насекомые жесткокрылые, т. XXI, 1936; т. XXII, 1940; т. XXIII, вып. 1, 1958.
Погориляк И. М. Короеды и биологические основы борьбы с ними. Ужгород, 1973 (на укр. яз.).
Положенцев П. А. Энтомоинвазия ветровальной сосны.- Научн. зап. Воронежск. лесотехн. ин-та, т. XII. Воронеж, 1953.
Прозоров С. С. Большой черный усач на пихте сибирской.- Тр. Сибирск. лесотехн. ин-та, т. XXI, вып. 2. Красноярск, 1958.
Рихтер А. А. Обзор златок европейской части СССР.- Сб. Зоологич. ин-та Арм. ССР. Ереван, 1944.
Рихтер А. А. Златки.- Фауна СССР, Насекомые жесткокрылые, т. XIII, вып. 2. М.-Л., 1949, вып. 4, 1952.
Рожков А. С. Дерево и насекомые. Новосибирск, 1981.
Руднев Д. Ф. Большой дубовый усач в лесах Советского Союза. Киев, 1957.
Руднев Д. Ф., Смелянец В. П. О природе устойчивости древесных насаждений к вредителям.- Зоолог, журн., т. 48, вып. 12. М., 1969.
Старк В. Н. Короеды.- Фауна СССР, Насекомые жесткокрылые, т. 31. М.- Л., 1952.
Строганова В. К. Рогохвосты Сибири. Новосибирск, 1968.
Трофимов В. Н. Биология ольхового рогохвоста в Хоперском заповеднике.- Научн. тр. Моск. лесотехн. ин-та, вып. 90. М., 1976.
Трофимов В. Н. Биология мраморного скрипуна Saperda scalaris L. (Coleoptera, Cerambycidae) по наблюдениям в Хоперском заповеднике. - В кн.: Экология и защита леса, вып. 5. Л., 1980.
Трофимов В. Н. Объем выборки при учете насекомых-ксилофагов - Лесоведение, 1979, № 6.
Турундаевская Т. М. Стеклянницы - вредители древесных пород Западного Казахстана. М., 1981.
Шевырев И. Я. Загадка короедов, 4-е изд./Под ред. П. Г. Трошанина. М., 1969.
Яновский В. М. Роль энтомофагов в динамике численности большого лиственничного короеда.- В кн.: Экология популяций лесных животных Сибири. Новосибирск, 1974.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'