6. Цветовое зрение
Способность различать цвета - одна из важнейших функций зрения, кажущаяся самоочевидной, но далеко не всеми правильно понимаемая, каждый светочувствительный аппарат, будь то фотографическая эмульсия, фотоэлемент или глаз животного, может неодинаково реагировать на свет, отличающийся по спектральному составу. Но о цветовом зрении говорят только тогда, когда разная реакция на излучения различного спектрального состава не зависит от их интенсивности. Отсутствие взаимозаменяемости видимых излучений произвольного спектрального состава - необходимый и формально достаточный критерий цветового зрения.
Для человека и высокоорганизованных животных, таких, как позвоночные и насекомые, в определение цветового зрения следует включить способность опознавать окраску предметов независимо от цвета освещения. Так, пчелы, как и человек, могут отличать, например, белые предметы от желтых даже тогда, когда первые освещены желтым светом, а вторые - белым, т. е. и те и другие предметы отражают свет одного и того же спектрального состава. Это называется константным восприятием окраски, без которого операция различения отраженного света по спектральному составу была бы недостаточной для надежной ориентации в многообразии окружающих предметов. В природе все живые организмы встречаются в основном с такими объектами, которые не светятся сами, а только отражают свет, падающий извне. Поэтому константность - чрезвычайно важное свойство зрительного анализатора, делающее его совершенным прибором для распознавания несамосветящихся предметов, спектральная характеристика которых зависит от цвета освещения. Механизм константности еще не выяснен до конца, но ясно, что он основан на сопоставлении спектральной информации от разных частей глаза и к нему причастны интернейроны с рецептивным полем, "оппонентным" по реакции на коротко- и длинноволновые спектральные лучи (см. ниже).
Изучение цветового зрения животных доступно путем регистрации их разнообразных реакций, зависящих от спектрального состава света. Это могут быть реакции целого организма (его поведение) или реакции отдельных частей зрительной системы (вызванные светом электрические потенциалы сетчатки и зрительных центров, фотохимические превращения и морфологические изменения в рецепторах под действием света и т. п.). Следует, однако, отметить, что если для понимания механизма цветоразличения может оказаться достаточным знание принципа кодирования в рецепторах информации о цвете и картины передачи соответствующих сигналов на разных этажах зрительного анализатора, то для окончательного суждения о цветоразличительных возможностях животного необходим только поведенческий эксперимент. Известны примеры (кошка), когда потенциальные цветоразличительные возможности сетчатки глаза почти не реализуются в поведении, и животное ведет себя в эксперименте и в обычной обстановке практически цветослепым. Это важно помнить биологу, когда он, опираясь на выводы, например электрофизиологических опытов, объясняет физиологический механизм той или иной реакции животного на цветные стимулы, особенно при этологических исследованиях.
Периферический аппарат цветоразличения
Понятие о светоприемниках. В колориметрии - науке об измерении цвета - вопрос о многообразии различимых глазом цветов решается путем нахождения числа светоприемников в сетчатке и спектральной чувствительности каждого из них в отдельности. Под светоприемниками понимают рецепторы с различной спектральной чувствительностью, передающие в оптический центр независимые и невзаимозаменяемые сигналы. Морфологически им соответствуют рецепторы сетчатки, как правило, с различным пигментом и обособленным каналом связи - аксоном. Светоприемники находятся на входе зрительной системы, поэтому излучения, неразличимые - тождественные - для светоприемников, неразличимы и для зрительного центра. Светоприемники обладают свойством аддитивности, которое выражается в том, что излучения А и В, приводящие светоприемник в одно и то же состояние, обычно остаются для него неразличимыми и после прибавления к каждому любого третьего излучения С, т. е. если А = В, то и А + С=В + С. Аддитивность светоприемников позволяет характеризовать их спектральной чувствительностью в строго физическом смысле.
Необходимым условием для спектрального анализа света сетчаткой считается присутствие в ней минимум двух типов светоприемников с различной спектральной характеристикой. При одном работающем светоприемнике любые излучения могут привести его в тождественное состояние, в результате чего декодирующее центральное устройство по величине сигналов светоприемника сможет судить одновременно только об одном параметре света - о его интенсивности, а не о цвете. При двух и большем числе одновременно работающих светоприемников с несовпадающей спектральной чувствительностью по совокупности их сигналов возможен анализ не только интенсивности, но и спектрального состава света. Например, большинство пар монохроматических излучений, равновозбуждающих светоприемник А, окажется нетождественным для светоприемника В. Поэтому в величине и соотношении ее у сигналов двух светоприемников А и В будет закодирована информация как об интенсивности, так и спектральном составе света. Три типа светоприемников обеспечат различение большего числа излучений. В центральной ямке сетчатки человека имеется, как известно, три типа светоприемников. Минимум три типа светоприемников существует и в глазу пчелиных и дневных бабочек, образ жизни которых связан с необходимостью опознавать по окраске цветущие растения.
Спектральная чувствительность светоприемника определяется в первую очередь тем зрительным пигментом, который заключен в фоторецепторе. В ряде случаев на пути лучей к фоторецептору стоят цветные светофильтры в виде, например, экранирующего пигмента, селективно пропускающего свет. Поэтому спектральная чувствительность светоприемника может зависеть также и от оптических свойств тех сред, которые фильтруют свет на пути к рецепторам.
Измерение спектральной чувствительности светоприемников. Из предыдущего изложения ясно, что изучение механизма цветового зрения необходимо начинать с измерения числа типов свето-приемников глаза и относительной спектральной чувствительности каждого в отдельности. Существует несколько методов изучения светоприемников.
Чаще всего прибегают к наиболее простому методу равных ответов. Он заключается в подборе и измерении энергии различных монохроматических лучей, вызывающих равную по амплитуде ЭРГ глаза или равную поведенческую (обычно фототропическую) реакцию насекомого. Результат эксперимента представляют обычно в виде кривой величин, обратных числу квантов (1/Q) или энергии (1/Е) спектральных стимулов, вызывающих равный ответ.
Получаемая "кривая равных спектральных ответов" темноадаптированного глаза сама по себе почти не несет информации ни о числе светоприемников, ни о спектральной чувствительности каждого из них. Но если дополнительно измерить кривую равных ответов при фоновом цветном подсвете, то по ее изменению можно с большой вероятностью судить о числе зрительных пигментов в сетчатке и на основе этого косвенно предполагать число светоприемников и их спектральную характеристику. Идея заключается в том, что фоновое цветное излучение, например синее, сильнее разлагает ("утомляет") прежде всего тот зрительный пигмент, который более чувствителен к нему. Поэтому кривая равных ответов, измеренная при адаптации к синему, будет отличаться от кривой темноадаптированного глаза только в том случае, если сетчатка содержит различные зрительные пигменты. Остается неясным, однако находятся ли разные пигменты в разных фоторецепторах или они содержатся во всех рецепторах в виде смеси. Между тем о разных типах светоприемников можно было бы говорить только в первом случае.

Рис. 57. Кривая спектральной эффективности зрительной клетки с 540 нм кузнечика Tetligonia cantans (по Вишневской, Мазохину-Поршиякову, 1972). А - снимок с экрана осциллографа ответов клетки на. 19 равноэнергетических спектральных стимулов с λ=390-669 нм. Б - кривая спектральной эффективности (1) и рассчитанная по ней кривая спектральной чувствительности (2). Вертикальные черточки - среднее квадратичное отклонение. По оси ординат в одном и том же масштабе в относительных единицах отложены амплитуды ответов клетки (для 1) и величин, обратных энергии света, вызывающего равные по амплитуде спектральные ответы (2); по абсциссе - длина волны света, нм
Надежнее в этом отношении метод измерения спектральной эффективности отдельных зрительных клеток. Глаз возбуждают с помощью специального стимулятора обычно равно-квантовыми и равными по продолжительности вспышками различных монохроматических лучей, регистрируя при этом внутриклеточным микроэлектродом амплитуду деполяризационного потенциала фоторецептора (рис. 57, А). Непосредственно в эксперименте получают так называемую "кривую спектральной эффективности" - кривую, огибающую амплитуды ответов клетки на равные спектральные стимулы (рис. 57, Б). Светоприемники целесообразнее характеризовать относительной спектральной чувствительностью приведенной к максимуму кривой, обратной отношению энергии света различных длин волн, одинаково возбуждающего фоторецептор. Ее легко рассчитать из кривой спектральной эффективности, если известна зависимость амплитуды потенциала фото-рецепторной клетки (в мВ) от интенсивности света.
Для расчета кривой спектральной чувствительности фоторецептора достаточно иметь амплитудные характеристики данной клетки для различных монохроматических лучей, но такой эксперимент более длителен и поэтому трудновыполним, так как клетка с введенным в нее микроэлектродом "живет" недолго - несколько минут.
Метод микроспектрофотометрии - изящный прием определения спектрального поглощения пигментов in situ. Именно этим методом недавно впервые были определены кривые поглощения колбочковых зрительных пигментов человека. Он заключается в том, что с помощью микроспектрофотометра измеряют спектральное поглощение отдельного фоторецептора непосредственно в живой сетчатке. Диаметр светочувствительного элемента фоторецептора составляет всего несколько микрометров, у насекомых диаметр рабдомера равен 0,5-2,0 мкм - отсюда трудность использования метода, так как требуется сфокусировать свет строго на одном рабдомере, а не на нескольких.
Наиболее надежен в отношении изучения спектральной чувствительности сетчатки метод мелькающей колориметрии, или метод нуль-установки, посредством которого судят о числе светоприемников по числу базисных (основных) излучений, необходимых и достаточных для воспроизведения в смеси друг с другом всех спектральных цветов. Он основан на подборе визуальных цветовых равенств (тождеств) и измерении энергии отдельных их компонентов: базисных и монохроматических излучений. В разработке его большую роль сыграл выдающийся отечественный специалист в области физиологической оптики проф. Н. Д. Нюберг. Эксперимент ведется на колориметре замещения, позволяющем быстро заменить монохроматическое излучение на смесь базисных цветов и наоборот. Критерием тождественности для насекомого каждого монохроматического излучения подобранной смеси базисных цветов принимают отсутствие ЭРГ в момент их замены друг на друга. Сама возможность такой подмены, остающейся незамеченной глазом, служит доказательством объективности и надежности метода.
Величину отношения энергии (в процентах от максимума) каждого базисного цвета (Еλ1-n ) в их смесях друг с другом к энергии каждого визуально тождественного этим смесям монохроматического цвета (Eλ) изображают в виде так называемых кривых сложения базисных цветов. Кривые сложения, т. е. функции 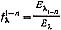 , характеризуют с точностью до их линейной комбинации спектральную чувствительность каждого светоприемника. Если видимый глазом спектр имеет одномерные - монохроматические - области, где работает по одному светоприемнику, то кривые сложения базисных цветов легко пересчитать непосредственно в кривые спектральной чувствительности светоприемников.
, характеризуют с точностью до их линейной комбинации спектральную чувствительность каждого светоприемника. Если видимый глазом спектр имеет одномерные - монохроматические - области, где работает по одному светоприемнику, то кривые сложения базисных цветов легко пересчитать непосредственно в кривые спектральной чувствительности светоприемников.
Колориметрический метод рассчитан не на определение зрительных пигментов сетчатки, а только на то, чтобы непосредственно найти число фактически работающих в глазу светоприемников и определить их спектральную чувствительность. Иными словами, он более других методов адекватен задачам описания светоприемников сетчатки и измерения многообразия цветов, различимых глазом. Именно колориметрические измерения послужили в свое время математику Грассману экспериментальной основой для выведения трех известных законов оптического смешения цветов для человеческого глаза.
Зрительные пигменты. Зрительный акт начинается с момента поглощения фотонов зрительным пигментом фоторецептора. Молекула фотопигмента состоит из двух непременных частей: прозрачной белковой - опсина и красящего хромофора - ретиналя, представляющего собой альдегид витамина A1 или А2. Под действием поглощенного кванта света молекула пигмента фотоизомеризуется: переход из цис- в трансконфигурацию. В этом и состоит единственная роль поглощаемого фотона, так как все дальнейшие реакции в фоторецепторе уже не зависят от наличия света. Чтобы вызвать посредством фотоизомеризации пигмента реакцию зрительной клетки, достаточно хотя бы одного кванта света.
Зрительных пигментов насекомых известно пока немного, что связано с трудностями их выделения и идентификации. Например, чтобы извлечь зрительный пигмент у медоносной пчелы, американскому исследователю Т. Голдсмису (1958) пришлось измельчить 20 тыс. голов этого насекомого. Экстрагируют фотопигменты насекомых, обычно дигитаниновым раствором в нейтральном фосфатном буфере. Чаще всего у насекомых встречаются фотопигменты типа родопсина (палочкового пигмента позвоночных) с λmах между 440-510 нм и вторым пиком поглощения в ультрафиолетовых лучах. Например, для мух характерен родопсин с λmах = 495 им. Фотохимический цикл его превращения может быть описан как P495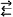 М580: нативный пигмент Р495 под действием света переходит в термостабильный фотопродукт - метародопсин М580, который регенерирует в исходный пигмент не только в темноте, но и под действием света, в особенности желтого. В глазу, адаптированном к обычному "белому" свету, между Р495 и количеством М580 устанавливается фотохимическое равновесие. Видимо, особый зрительный пигмент, чувствительный в основном к ультрафиолету, свойствен только насекомым. Такой фотопигмент с λmах =345 нм извлечен из головы булавоуски Ascataphus macaronius и некоторых других насекомых.
М580: нативный пигмент Р495 под действием света переходит в термостабильный фотопродукт - метародопсин М580, который регенерирует в исходный пигмент не только в темноте, но и под действием света, в особенности желтого. В глазу, адаптированном к обычному "белому" свету, между Р495 и количеством М580 устанавливается фотохимическое равновесие. Видимо, особый зрительный пигмент, чувствительный в основном к ультрафиолету, свойствен только насекомым. Такой фотопигмент с λmах =345 нм извлечен из головы булавоуски Ascataphus macaronius и некоторых других насекомых.
Рис. 58. Кривые спектральной чувствительности светоприемников комнатной мухи Musca domestiса (А) и стрекозы Libellula quadrimaculata (Б) (по Мазохину-Поршнякову, 1965). а, б - коротко- и длинноволновые светоприемники мухи; в - единственный светоприемник верхней половины глаза стрекозы; г, д - коротко и длинноволновые светоприемники нижней половины глаза стрекозы. По оси ординат - относительная чувствительность к равиоэнергетическому спектру в процентах от максимума; по оси абсцисс - длина волны света, нм
Влияние фильтрующего аппарата на спектральную чувствительность глаза. Аппарат светоизоляции фоторецепторов подробнее изучен у мух, у которых он особенно сильно влияет на спектральную чувствительность и другие параметры глаза. В норме в глазах Calliphcfra и других мух есть два типа экранирующих пигментов; красные оммохромы и желтоватые птерины. Первые задерживают все лучи, кроме красных, вторые задерживают в основном ультрафиолетовые лучи. Поэтому все спектральные лучи могут проникать к рабдому отдельного омматидия только через его собственную фасетку, а красные - и через соседние фасетки, так как экранирующий пигмент для них прозрачен. В результате чувствительность глаза мух к красным лучам повышена: кривая равных ответов имеет характерный пик у λ=620 нм, а сетчатка оказалась наделенной своеобразным дополнительным светоприемником. У мух нет специального красночувствительного зрительного пигмента, но есть красночувствительный (λmах =620 нм) светоприемник "фильтровой" природы. Он представляет собой, по-видимому, совокупность из шести рабдомеров каждого омматидия, которые, по сравнению со сдвоенным центральным (седьмым и восьмым) рабдомером, находятся в преимущественных условиях освещения красными лучами за счет красных рефлексов от экранирующего пигмента и проникновения красных лучей со стороны смежных омматидиев. Вместе с основным зеленочувствительным (λmах≈500 нм) светоприемником этот дополнительный "фильтровой" светоприемник, в основе которого лежит тот же самый или близкий родопсиноподобный зрительный пигмент, позволяет мухам различать цвета (рис. 58, А).
Системы цветового зрения. Видимый насекомыми спектр шире, чем у человека: он включает ультрафиолетовые лучи с длиной волны вплоть до 240 нм, хотя в естестенной обстановке лучи с λ<300 нм отсутствуют (см. ниже, рис. 60, А). О наборе светоприемников у разных насекомых дает представление табл. 11.
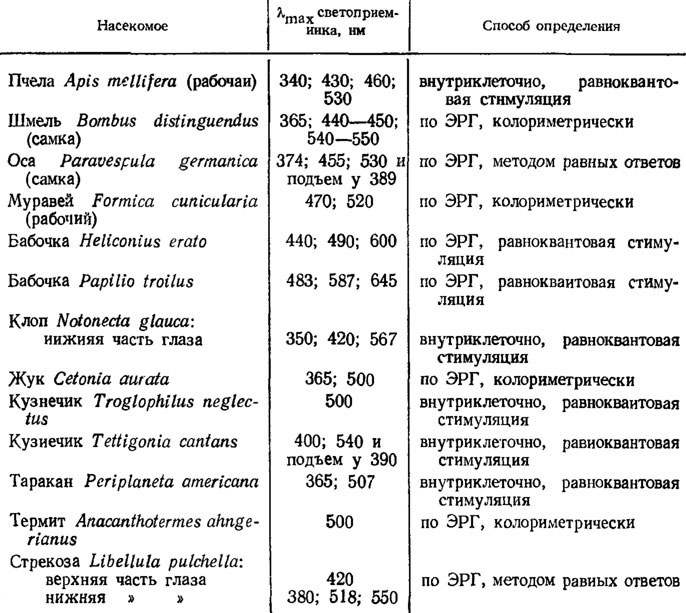
Таблица 11 Примеры светоприемников у взрослых насекомых
Цветослепые формы, по крайней мере среди взрослых насекомых, встречаются крайне редко. К ним относятся скрытоживущие виды: пещерный кузнечик Troglophilus neglectus, термит Anacantho termes ahngerianus, мучной хрущак Tenebrio molitor, которые имеют по одному-единственному светоприемнику.
В основном приходится иметь дело с ди- и трихроматами, периферический цветоразличающий аппарат которых образован двумя или тремя типами светоприемников. Иногда, как у медоносной пчелы, в глазу бывает четыре спектральных типа рецепторов, но поведенческие опыты показывают, что цветовое зрение пчелы трихроматическое и, значит, сигналы двух близких рецепторов с λmах, равной 430 и 460 нм, взаимозависимы. Трихроматами среди насекомых бесспорно являются пчелиные и, возможно, дневные бабочки.
Для всех насекомых характерна обычно высокая чувствительность к ультрафиолету, определяющаяся специальным "ультрафиолетовым" светоприемником, как у булавоуски Ascalaphus macaronius или жука-бронзовки, а также частично (иногда целиком, как у мух) вторым пиком поглощения (в ультрафиолетовых лучах) у родопсиноподобного зрительного пигмента. Поэтому видимый насекомыми спектр простирается от ультрафиолетовых лучей (λ=300 нм) до красных включительно (λ=650 нм). Судя по колориметрическим измерениям в электрофизиологическом эксперименте и результатам поведенческих опытов, близкие монохроматические цвета насекомые дифференцируют наиболее четко в средней части спектра (зеленые и сине-фиолетовые цвета), где резко изменяется спектральная чувствительность у разных светоприемников. Цвета, лежащие на краях спектра, а именно: оранжево-красные (с одной стороны) и далекие ультрафиолетовые (с другой стороны), - насекомые различают слабо.
У трихроматов (пчелиных) для цветоразличения, видимо, особенно велика роль рецепторов ультрафиолета. Например, медоносные пчелы легко отличают смесь из 99% желтых лучей с c λ=588 нм + 1% УФ с λ=360 нм от 100% желтых лучей с λ=588 нм.
У некоторых насекомых, в особенности у хищников, охотящихся в воздухе, замечено различие в спектральной чувствительности верхней и нижней частей глаза. Так, у стрекоз Libellula (рис. 58, Б), булавоуски Ascalaphus macaronius верхняя половина глаза цветослепа, но обладает высокой чувствительностью к лучам, рассеиваемым голубым небом; нижняя половина глаза способна к цветоразличению.
Кодирование и передача спектральной информации в мозг
Судя по физиологическим и морфологическим признакам, отдельный омматидий представляет собой единицу, достаточную для спектрального, а также поляризационного анализа света. Различными способами показано, что слагающие его зрительные клетки неодинаковы по спектральной чувствительности. Так, у медоносной пчелы в каждом омматидии имеется три типа светоприемников; у американского таракана Periplaneta americana из восьми зрительных клеток омматидия пять являются рецепторами с λmах =507 нм и три клетки - рецепторами с λmах = 365 нм.
О способе передачи с сетчатки сигналов о цвете и их последующей обработке пока известно немного. Считается, что электрическая связь между разными рецепторами внутри омматидия практически отсутствует. В таком случае можно думать, что постретинальные сигналы отдельного омматидия поступают в оптические патроны ламины по многим, обычно по шести независимым, каналам, а в медуллю поступает информация, по крайней мере, по двум разным каналам: аксонам базальных клеток и интернейронам оптического патрона. К этому нужно добавить, что в действительности в каждом омматидии есть обычно две базальные клетки, а в оптическом патроне - два-три перпендикулярных интернейрона плюс несколько центробежных волокон.
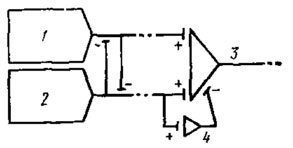
Рис. 59. Модель передачи информации о цвете в зрительной системе бабочки Epargireus clarus (пo Swihart, 1969): 1 - 'синий' рецептор; 2 - 'зеленый' рецептор; 3 - цветное ганглиозиое волокно; 4 - вставочный интернейрон с 'задержкой'.
В оптических ганглиях и в особенности в грибовидных телах переднего мозга удается электрофизиологически зарегистрировать волокна, отличающиеся специфической реакцией на свет различных длин волн, что говорит об их причастности к обработке спектральной информации (см. рис. 5). У бабочки Epargyreus clarus замечен особенно интересный тип цветного "оппонентного" волокна. Это волокно интенсивно разряжается импульсами при освещении глаза синим светом, очень коротко отвечает на зеленые лучи, но не возбуждается "белыми" лучами. Здесь синий свет тормозит ответ волокна на зеленые лучи и наоборот; при этом ответ на синий свет возникает без задержки, а на зеленый - с задержкой. В сетчатке этой бабочки только два спектральных типа рецепторов: с λтax =440 нм (синий светоприемник) и λmах =540 нм (зеленый светоприемник), - поэтому легко представить себе модель связанного с ними цветного оппонентного волокна (рис. 59).
У другой бабочки - Papilio troilus - среди разнообразных цветных интернейронов мозга наиболее интересны волокна с двумя пиками чувствительности - в синем и оранжевом участках спектра. Эта бабочка спонтанно предпочитает синие и оранжевые цветки растений, поэтому вполне вероятно, что именно названные интер-нейроны управляют ее пищевым поведением.
Поведенческие реакции на цветные стимулы
В начале раздела о цветовом зрении мы уже говорили о том, что знания периферических и даже центральных механизмов зрения еще недостаточно для правильного предвидения визуальных реакций целого организма. Дело в том, что с помощью зрительного и любого другого анализатора животное не только различает стимулы или объекты, но и опознает их как таковые - выбирает определенные объекты среди многих. Целесообразные поведенческие реакции всегда развиваются с участием врожденной или приобретенной памяти. Именно память позволяет насекомому опознать требуемый объект как таковой, например, как съедобный, но о содержании памяти мы можем пока судить исключительно по поведению животного. В этом заключается незаменимая роль поведенческого эксперимента.
Опыты с регистрацией поведения в ответ на те или иные стимулы - технически более простой способ изучения цветового зрения, и они были поставлены на насекомых раньше электрофизиологических измерений чувствительности фасеточного глаза. Мы остановимся на двух видах поведенческих реакций на цвет: спонтанном (врожденном) предпочтении и выборе на основе индивидуального научения.
Замечено, что многие антофильные насекомые: бабочки, мухи, различные пчелы - избирательно посещают цветущие растения, предпочитая те из них, которые имеют определенную окраску. Эксперименты с цветными макетами подтвердили этот вывод. Так, если поместить в обширный садок с недавно вышедшими из куколок голодными парусниками Рарiliо troilus макеты цветков из пурпурной, синей, зеленой, желтой, оранжевой, красной бумаги, а также из серой бумаги разной светлоты, то бабочки будут садиться преимущественно на синие и пурпурные, отчасти на оранжевые макеты. Тот факт, что преимущественный выбор синих, пурпурных и оранжевых макетов был сделан в присутствии серых разной светлоты, позволяет заключить об опознавании предлагаемых макетов именно по цвету, а не по светлоте. В этом опыте бабочки демонстрируют разную реакцию на объекты, окрашенные по-разному, независимо от яркости (светлоты) последних, а это отвечает сделанному нами выше определению цветового зрения.
Мухи Eristalis, жуки Meligaethes, а также тли в период питания предпочитают желтые окраски. Стремление к желтому положено в основу привлекающего действия липких или наполненных водой ловушек для тлей и других вредных насекомых. В разные периоды жизни насекомого его цветовое представление может измениться, в частности тлей во время расселительных полетов привлекают преимущественно синие поверхности - цвет безоблачного неба как признак открытого пространства.
Врожденное стремление насекомого к излучениям определенного спектрального состава непосредственно свидетельствует о его способности к различению цвета. Этой особенностью поведения можно пользоваться как способом быстрой проверки способности насекомого к цветоразличению.
Но более гибок и информативен другой способ доказательства и подробного изучения цветового зрения, основанный на выработке условного рефлекса на определенный цвет. Чаще всего вырабатывают пищевой рефлекс на различение цветных образов. К. Фриш еще в 1914 г. первым поставил такой эксперимент на пчелах. На специальном столике он разложил в виде шахматного поля 16 бумажных квадратиков, среди которых 15 были изготовлены из "ахроматической" бумаги разной светлоты: от белой до черной, а 16-й квадратик был вырезан из голубой бумаги. На голубом образце была сладкая приманка, а на остальных в аналогичных сосудах - вода. После нескольких посещений приманки пчелы находили голубой образец в любом месте шахматного поля и садились на него даже тогда, когда приманку снимали с него и все образцы покрывали новым чистым стеклом. В условиях такого эксперимента пчелы могли опознать голубой образец только по цвету, а не по его светлоте. Если бы они ориентировались по светлоте голубого квадратика, то среди ахроматических образцов обязательно нашелся бы равносветлый голубому, и пчелы садились бы на него так же часто, как на голубой. В действительности же обученные пчелы практически не ошибались в выборе голубого, что исчерпывающим образом доказывает их способность к различению цвета.
Впоследствии также путем обучения Г. А. Мазохиным-Поршняковым была показана способность пчел дифференцировать близкие окраски, например желтую и оранжевую. В этом опыте шахматное поле было сложено из оранжевых квадратиков разной светлоты, среди которых присутствовал один желтый квадратик с приманкой. После обучения в контрольном эксперименте желтый образец был заменен другим, желтым, но более темным, и, несмотря на это, пчелы садились только на желтый квадратик.
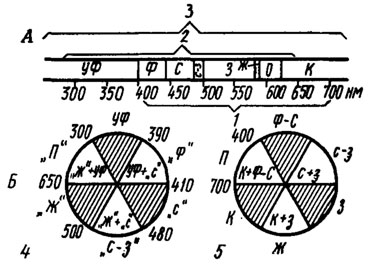
Рис. 60. Границы видимого спектра (А) для человека (1), медоносной пчелы Apis mellifera (2), мухи Drosophilla me-lanogaster (3) и цветовой круг (Б) для медоносной пчелы (4) и человека (5) (рис. 4,5 - по Daumer, 1956). Заштрихованы (Б) области основных цветов, которые при оптическом смещении друг с другом дают все остальные цвета. Между ними лежат промежуточные цвета, возникающие при оптическом смешении двух соседних основных цветов. Цвета противоположных секторов являются дополнительными. Числа - длина волны света, нм. Буквы в кавычках - условные названия пчелиных цветов. ж - желтый, з - зеленый, к - красный, о - оранжевый, п - пурпурный, с - синий, с-з - сине-зеленый, уф - ультрафиолетовый, ф - фиолетовый, ф-с-фиолетово-синий
В широкоизвестных опытах с обучением пчел К. Даумер доказал, что для этих насекомых солнечный белый свет неотличим от смеси трех спектральных цветов: 55% желтого с λ=588 нм плюс 30% сине-фиолетового с λ=440 нм и плюс 15% ультрафиолетового с λ=360 нм. Тем самым оказалось, что ультрафиолетовые лучи оказываются для глаза пчел дополнительным цветом к смеси желтых и сине-фиолетовых лучей, а смесь желтых и ультрафиолетовых лучей дает "пчелиный пурпурный" цвет, отсутствующий в спектре*. Как известно, пурпурный цвет, воспринимаемый человеком и тоже отсутствующий в спектре, представляет собой смесь синих лучей с красными и считается дополнительным к зеленому цвету (рис. 60, Б). Результаты этих выдающихся опытов наряду с последующими электрофизиологическими экспериментами послужили основанием для утверждения о трихромазии зрения пчел.
* (Дополнительными цветами называют такую пару цветов, которые при смешении друг с другом дают белый (серый) цвет, т. е. дополняют друг друга до белого. Здесь имеется в виду оптическое смешение излучений, а не механическое смешение красок на палитре.)
Пчелы очень быстро и прочно запоминают окраску пищевых объектов. Достаточно двух-трех подкреплений выбора, чтобы пчела на всю жизнь (летом - две-три недели) запомнила цвет кормушки, в которой она нашла сладкую приманку. Показано, что пчелы способны запоминать комбинацию из двух-трех окрасок в качестве отличительного признака пестро раскрашенных предметов. Например, они легко научаются безошибочно выбирать тестовую карту с приманкой, составленную из оранжевых и голубых квадратиков, среди четырех других карт без приманки: оранжевой с желтым, голубой с желтым, желтой с зеленым и зеленой с оранжевым. Среди такого набора карт подкрепляемую комбинацию красок можно запомнить только по двум признакам: "есть и оранжевый и голубой" или "нет ни желтого, ни зеленого". Несомненно, что пестрая раскраска венчиков - двухцветная у нивяника (Leucanthemum vulgare) или трехцветная у пикульника (Galeopsis speciosa) - помогает пчелам опознать эти растения среди красочного разнообразия других. При скученном расположении ульев пчеловоды иногда красят их также в два цвета, чтобы облегчить пчелам поиск своей семьи.
Другие насекомые, например осы Vespidae, не имеющие такой тесной связи с цветущими растениями, как пчелиные, хотя и легко научаются отличать одну окраску от другой, но совершенно не способны запоминать парные и тем более тройственные сочетания цветов.
Удивительную сложность визуального поведения пчел, связанного с распознаванием окрасок, может иллюстрировать еще один пример. В ходе тренировки по специальной схеме у пчелы удается выработать условный рефлекс на обобщение типа окраски по признаку "двухцветность", а именно: пчела научается выбирать всегда двухцветную тестовую карту независимо от конкретных цветов, составляющих ее. Например, обученная пчела выбирает только двухцветную карту среди ранее незнакомого ей набора из трех карт: 1) двухцветная- голубой-(-желтый цвета, 2) одноцветная голубая и 3) одноцветная желтая. В другом наборе: 1) оранжевый+синий, 2) сплошь синяя и 3) сплошь оранжевая карты - она избирает также двухцветную. Такой сложный выбор основан на обобщенном представлении о двухцветности, на противопоставлении всех двухцветных образцов всем одноцветным образцам.
В заключение этого раздела, в котором особенно часто употреблялись названия воспринимаемых человеком цветов, следует предостеречь читателя о незаконности переноса наших ощущений на восприятие животных. Ощущение цвета, запаха и прочего стимула всегда субъективно, т. е. принципиально невозможно ощутить ощущения другого субъекта, чтобы убедиться в их тождественности. Но можно (и именно так поступают исследователи) проверить сходство реакций у разных индивидов на физически тождественные стимулы, и в этом смысле можно говорить о равной способности различать или отождествлять разные раздражители. Поэтому, когда говорилось, что пчела отличает "желтый цвет от зеленого", имелось в виду, что те излучения, которые называем желтыми мы, и пчелы могут отличать от излучений, вызывающих у нас ощущение зеленого цвета. Но как выглядят для пчел, как ощущаются ими эти излучения - мы не знаем.
Окраски растений и покрова тела насекомых, различимые фасеточным глазом
Разнообразие окрасок несамосветящихся объектов - растений, большинства животных, минералов и т. п., - которое мы привыкли видеть в естественной обстановке, порождено тем, что, во-первых, разные предметы по-разному отражают спектральные лучи, и, во-вторых, солнечный свет есть смесь монохроматических лучей широкого непрерывного диапазона спектра. Так, оранжевые окраски отличаются от желтых тем, что первые сильнее поглощают сине-фиолетовые и зеленые лучи. Если же смотреть на эти окраски при искусственном освещении натриевой лампой, то большинство образцов оранжевой окраски мы не отличим от желтых. Свет натриевой лампы монохроматический, желтый, а желтые и оранжевые окраски отражают эти лучи практически одинаково сильно, поэтому под натриевой лампой названные образцы визуально неразличимы. Из этого примера ясно, как зависит возможность различения окрасок от их отражательной характеристики и спектрального состава освещения.
Экспериментально доказано, что зрительный анализатор человека и, видимо, всех цветозрячих животных приспособлен для опознавания предметов главным образом по их спектральной отражательной характеристике, а не спектральному составу тех лучей, которые отражают данные предметы в данный момент времени. Один и тот же предмет в зависимости от цвета освещения может отражать свет разного спектрального состава, и задача зрительного анализатора заключается в том, чтобы выделить ту инварианту, которая характеризует предмет. Последнее достигается благодаря автоматическому (у человека - подсознательному) введению "поправки на освещение" (Гельмгольц) к цвету рассматриваемого предмета за счет сравнения отражаемого им спектра со спектром других предметов в поле зрения. Мы уже говорили, что это свойство зрения называют константностью восприятия окраски. Константность цветовосприятия позволяет опознать предмет в разных условиях освещения, когда он отражает свет разного спектрального состава. Например, пчела зрительно замечает цветок одуванчика не только на солнце, но и в тени, где он объективно зеленый, поскольку в спектре голубого неба, создающего освещение в тени, очень мало желтых я тем более оранжевых лучей. Точно так же белый цветок в тени - объективно зеленовато-голубой за счет освещения голубым небом и зелеными рефлексами (отраженными лучами) от листьев растений, но это обстоятельство не затрудняет поиск насекомым, привыкшим кормиться на белых цветах.
Для человека многообразие окрасок природных объектов порождается избирательным отражением ими света в диапазоне спектра от фиолетовых лучей до красных включительно. Но насекомые чувствительны к ультрафиолетовым лучам, поэтому характер отражения этих лучей позволяет им распознавать окраску предметов в той же мере, как на наши цветовые оценки влияют сине-фиолетовые лучи. Прежде всего для глаза насекомых не идентичны по цвету все те окраски, которые мы называем белыми. Например, давно замечено, что пчелы легко отличают кажущиеся нам одинаково белыми две поверхности, одна из которых выкрашена цинковыми белилами, а другая-свинцовыми. Свинцовые белила гораздо сильнее отражают ультрафиолетовые лучи; они "действительно белые" для насекомых в отличие от цинковых белил, которые сильно поглощают ультрафиолетовые лучи и поэтому имеют для насекомых цвет, дополнительный к ультрафиолетовому. В частности, для пчел дополнительным к ультрафиолетовому является сине-зеленый цвет.

Рис. 61. Цветки и листья лапчатки Potentilla anserina (а) и лютика Ranunculus aces (б) (по Мазохину-Поршнякову, 1965). Снимок А сделан в ультрафиолетовых лучах (внизу - шкала стандартов с коэффициентами отражения: 22, 11, 6 и 3%). Снимок Б сделан в видимом человеком свете
Измерение коэффициента отражения спектральных лучей цветками растений показало, что белые венчики, как правило, слабо отражают ультрафиолетовые лучи и, следовательно, выглядят для насекомых окрашенными в дополнительный к ультрафиолетовому цвет. Желтые, оранжевые и красные цветки отражают ультрафиолетовые лучи иногда очень сильно. Из них цветок красного мака Papaver rhoeas выглядит для насекомых, не чувствительных к красным лучам, например для пчел, чисто ультрафиолетовым, поскольку венчик мака практически не отражает иных лучей, кроме красных и ультрафиолетовых. Неожиданной оказалась раскраска для глаза насекомых некоторых цветков, кажущихся человеку однотонножелтыми. Всем знакомы очень похожие друг на друга желтые цветки гусиной лапки Potentilla anserina и лютика едкого Ranunculus acer, но на снимке в ультрафиолетовых лучах они выглядят резко различными (рис. 61). Цветок гусиной лапки неравномерно отражает ультрафиолет - периферия венчика отражает сильнее (около 20%), чем его внутренняя часть вместе с генеративными элементами (около 2%)*. Поэтому цветок гусиной лапки в отличие от цветка лютика выглядит насекомым двухцветным за счет "скрытого" (от взора человека) рисунка. Скрытые рисунки помогают насекомым не только различать разные цветки, но и быстрее находить нектар. Узор, за счет более сильного поглощения ультрафиолетовых лучей внутренней частью цветка, служит пчелам визуальным указателем нектара и пыльцы. Как только пчела подползает к указателю нектара, даже на искусственно раскрашенной модели цветка, она опускает голову и выбрасывает хоботок, готовясь к приему пищи. Указатели нектара могут иметь различную окраску, не обязательно ультрафиолетовую.
* (Для оценки интенсивности отражения обычно фотографируют вместе с растением шкалу с известными коэффициентами отражения. Тогда, сравнивая на негативе или позитиве плотность почернения элементов растения с элементами шкалы, можно судить о степени отражения света растением.)

Рис. 62. Фотоснимки самца (1) и самки (2) бабочки Gonepteryx rhamni, сделанные в ультрафиолетовых лучах с 365 нм (по Мазохину-Поршнякову, 1965)
Скрытые рисунки несут на крыльях некоторые дневные бабочки, у которых особенности отражения телом ультрафиолетовых лучей следует считать проявлением скрытого для глаза человека полового диморфизма. Так, на снимке в ультрафиолетовых лучах самец бабочки-лимонницы Gonepteryx rhatmni имеет скрытый рисунок за счет более сильного отражения этих лучей основной частью поля переднего крыла (рис. 62). Самка лимонницы слабо и равномерно по всему полю переднего и заднего крыла отражает ультрафиолетовые лучи. Таким образом, самец лимонницы выглядит для самки двухцветным, а не однотонножелтым, каким мы воспринимаем его. Еще более интересен факт существования резкого полового диморфизма в окраске у таких видов, у которых человеческий глаз его практически не замечает. Например, белые и издалека (в полете) неразличимые нами самец и самка капустной белянки Pieris brassicae или боярышницы Aporia crataegi выглядят для насекомых по-разному за счет неодинакового отражения ультрафиолета. Самцы почти не отражают эти лучи, поэтому имеют для глаза насекомых цвет, дополнительный к ультрафиолетовому, тогда как самки за счет более сильного отражения лучей по всему спектру выглядят для насекомых иначе. Различие в отражении ультрафиолетовых лучей разными участками крыла, на наш глаз одинаково окрашенными, зависит в одних случаях от содержания особых пигментов, например птерина, в других - от оптических свойств и ориентации самих чешуек. Аналогичные различия у разных участков лепестка или венчика в целом, как полагают, только пигментной природы.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'