7. Устройство и работа простых глазков
Дорсальные глазки и их назначение
Дорсальные, или теменные, глазки свойственны взрослым насекомым, а также личинкам - нимфам - видов с неполным превращением. В типичном случае имеются три глазка, которые расположены в виде треугольника на лобно-теменной части головы: ближе к ротовому аппарату лежит непарный медиальный глазок, а выше него, по бокам, находятся два латеральных глазка. Медиальный глазок, парный по своему происхождению, нередко крупнее остальных, но он может отсутствовать. Реже встречаются случаи, когда редуцированы только боковые глазки.
Морфологи утверждают, что у ряда представителей разных семейств и разных отрядов глазки вовсе отсутствуют. Однако совсем недавно у дневных бабочек (Rhopalocera), бражников, медведиц и представителей других семейств чешуекрылых, ранее считавшихся лишенными глазков, на темени вблизи антенн обнаружено по паре небольших (26 мкм) глазков. У бражников в глубине головы под каждым внешним глазком лежит связанный с ним внутренний глазок. Строение и электрическая реакция на свет этих внутренних глазков характерны для обычных глазков других насекомых.
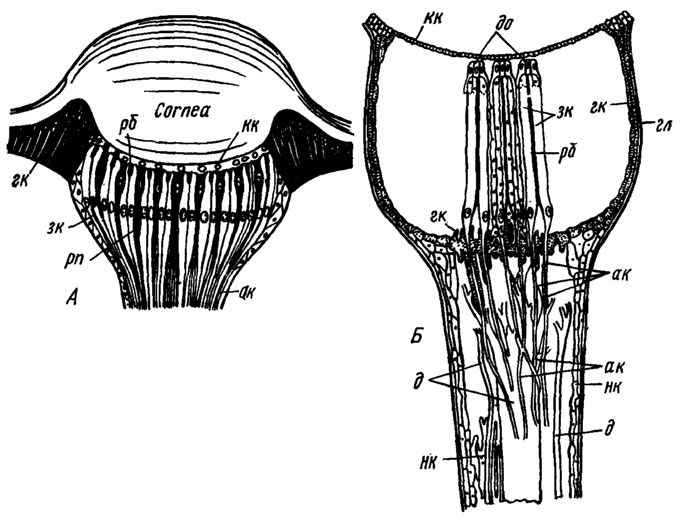
Рис. 63. Строение дорсальных глазков насекомых (продольный разрез, схематично): А - схема типичного глазка (по Weber, 1949); Б - устройство бокового дорсального глазка стрекоз (роговичная линза удалена) (по Ruck, Edwards, 1964). ак - аксоны зрительных клеток в области их синаптического контакта с интернейронами; гк - эпителиальные (гиподермальные) пигментные клетки; д - дендриты интернейронов; до - дистальные окончания клеток тапетума; зк - зрительные клетки, сгруппированные в ретинулы; кк - корнеагенные клетки; рб - рабдом; нк - непигментированные клетки оцеллярной ножки, часть которых футляром одевает нервные волокна; гл - расширенная часть глазка; рп - ретинальные пигментные клетки
Типичный внешний глазок имеет все три непременных для настоящего органа зрения морфофункциональных элемента: светопреломляющий, светочувствительный и светоизолирующий. Светопреломляющей линзой чаще всего служит прозрачное двояковыпуклое утолщение кутикулярного покрова (рис. 63, A, cornea). Под ним находится тонкий слой прозрачных корнеагенных клеток, образовавших линзу. К слою корнеагенных клеток обычно непосредственно примыкает вогнутая по форме линзы сетчатка, сложенная из отдельных ретинул, в которые сгруппированы по две-три удлиненные зрительные клетки (рис. 63, А, Б). Число ретинул варьирует от нескольких десятков до нескольких сотен. Внутри ретинулы, вдоль ее оси, лежит рабдом (рис. 63, Л, Б, рб), сложенный из рабдомеров отдельных зрительных клеток. Зрительная клетка имеет богатый эндоплазматический ретикулюм, многочисленные митохондрии и обычно гранулы темного экранирующего пигмента. Проксимальный конец клетки переходит в аксон (Б, ак), уходящий в оцеллярный нерв. Между ретинулами иногда вклиниваются узкие ретинальные пигментные клетки (А, рп) либо клетки трахеального тапетума (Б, до), хорошо отражающего свет. Светоизоляция сетчатки достигается в одних случаях тем, что ее кольцом охватывают столбчатые гиподермальные клетки, заполненные темным пигментом (Л, гк). В других случаях, когда отсутствуют ретинальные пигментные клетки, сетчатка целиком заключена в непрозрачный бокал, сложенный пигментными клетками (Б, гк).
Нередко встречаются отступления от описанного выше плана строения глазка. Например, у таракана Periplaneta ретинулы расположены хаотично, многослойной массой, так что их рабдомеры направлены в разные стороны. У первичнобескрылых насекомых, в частности у Machilis, роговичная линза едва намечается, а ретинулы и корнеагенные клетки лежат вперемешку друг с другом, не образуя обособленных слоев.
Оптический ганглий каждого глазка лежит вблизи его сетчатки в утолщенной части так называемой оцеллярной ножки, представляющей собой заключенную в футляр из непигментированных клеток (Б, нк) совокупность нервных волокон, уходящих в межцеребральную часть переднего мозга. В этом ганглии аксоны зрительных клеток вступают в многочисленные синаптические контакты с дендритами нейронов второго порядка (Б, д), тела которых лежат, видимо, в межцеребральной части мозга. Существенно, что число этих постсинаптических волокон значительно меньше числа пресинаптических волокон и что часть их отличается большой толщиной. В ножке медиального глазка саранчи Locusta migratoria содержится только шесть постсинаптических волокон: два более толстых, диаметром около 8 мкм, и четыре меньшей толщины - около 4 мкм. На обработанных хлоридом кобальта препаратах медиального глазка саранчи Schistocerca vaga обнаружено 17 больших билатерально симметричных глазковых интернейронов. В каждой ножке имеется по семь больших интернейронов, из которых четыре связаны сразу с двумя глазками. Один из интернейронов медиального глазка имеет в мозгу билатеральную связь, остальные ограничены каждый своей частью мозга. Обнаружена также связь ганглиозных интернейронов с оптическими ганглиями фасеточных глаз, в первую очередь с комиссурой между левым и правым оптическими ганглиями.
Судя по электрофизиологическим данным, сигналы с глазков влияют на активность нервных клеток не только переднего, но и среднего и заднего мозга головы. При освещении глазков спонтанная активность одних нервных клеток усиливается, других - тормозится. Замечена также импульсация в шейных коннективах нервной цепочки в ответ на возбуждение глазков: включение света или движение в их поле зрения предметов, контрастирующих с фоном.
Таким образом, дорсальные глазки связаны с мозгом чрезвычайно малым числом анатомически обособленных нервных каналов, и трудно ожидать, чтобы они передавали много информации. Но влияние сигналов с глазков прослеживается на многих мозговых центрах и даже на "исполнительной" части центральной нервной системы за пределами головы.
Электрическая активность глазка зависит от реакции его зрительных клеток и интернейронов оцеллярной ножки. Зрительные клетки реагируют на увеличение интенсивности света нарастающим деполяризационным потенциалом до 40 мВ. Крупные интернейроны ножки отвечают на свет гиперполяризационным потенциалом, максимальная амплитуда которого достигает 30 мВ. Для того чтобы вызвать ответ такого интернейрона, достаточно одной десятой интенсивности света, вызывающего реакцию отдельной зрительной клетки. Благодаря множеству контактов с разными рецепторами сетчатки динамический диапазон ответов интернейронов глазка, по сравнению с его зрительными клетками, сдвинут на более низкие интенсивности света.
Экстраклеточно легко отвести от глазка двухфазную ЭРГ с отрицательными и положительными волнами оn-эффекта, а от его ножки-спонтанную импульсацию, подавляемую светом. ЭРГ, например у саранчи-шистоцерки, состоит из трех компонентов, генерируемых соответственно деполяризационным потенциалом рецепторов, гиперполяризацией интернейронов и их off-активностью.
Из этих компонентов наиболее лабилен электрический ток, связанный с деполяризацией зрительных клеток. Судя по этому главному компоненту ЭРГ, мелькания сливаются для глазка стрекозы Pachydiplax только тогда, когда частота вспышек превышает 200 раз в секунду. Для глазка медоносной пчелы критическая частота слияния мельканий еще выше - 265 в секунду.
В темноте интернейроны оцеллярной ножки мух, пчелиных, саранчовых и других насекомых спонтанно генерируют 40-70 импульсов в секунду. Освещение сначала сильно подавляет частоту импульсации, а при достаточно ярком свете активность интернейрона совсем прекращается. Но по истечении двух-трех минут устанавливается новый уровень импульсации, который тем ниже, чем выше освещенность. При выключении света частота импульсов возрастает, но через две-три минуты возвращается к стационарному темновому уровню. Оба переходных процесса, видимо. отражают адаптационные изменения, протекающие в глазке достаточно быстро.
Следовательно, по данным электрофизиологии, глазок служит не только для регистрации изменения освещения, но и для того, чтобы отличать неизменное освещение от темноты. Поскольку частота постоянной импульсации глазков градуально зависит от силы света, они могли бы давать информацию и об абсолютной освещенности. Наконец, благодаря глазкам центральная нервная система насекомого получает в темноте высокое тоническое возбуждение.
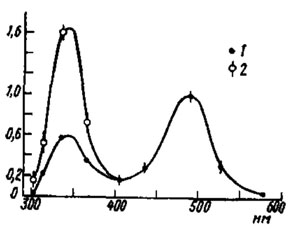
Рис. 64. Кривая равных ответов медиального дорсального глазка пчелы Apis mellifera (no Goldsmith, Ruck, 1958): 1 - значения, полученные при уравнивании высоты отрицательной волны оn-эффекта ЭРГ; 2 - значения, полученные при уравнивании высоты положительной волны оn-эффекта ЭРГ. По оси ординат - величины, обратные числу квантов света, вызывающего ЭРГ равной высоты; за 1,0 принято значение для λ=490 нм; по оси абсцисс - длина волны света, нм
О спектральной чувствительности глазков дает представление кривая равных ответов (ЭРГ) медиального глазка медоносной пчелы (рис. 64). Она двухвершинная, причем пик ультрафиолета (λ=340 нм), если кривую рассчитать по положительной волне оn-ответа ЭРГ, превышает подъем в сине-зеленой (λ=490 нм) части спектра. Зависимость формы ЭРГ от спектрального состава стимулирующего света можно расценивать как одно из указаний на цветоразличительную способность глазка.
Понять назначение дорсальных глазков до конца можно было бы при наличии их у всех насекомых, ведущих открытый активный образ жизни и лишенных признаков редукции других органов или упрощения общего уровня организации. Но морфологи утверждают, что нет глазков, например, у cлейней. Зрительное поведение слепней принципиально не отличается от поведения других мух с глазками, и в таком случае нужно думать, что функции глазков все же могут компенсироваться фасеточными глазами. С другой стороны, глазки в качестве дополнительного органа зрения появились у членистоногих очень рано. Они были уже у трилобитов и есть у мечехвостов в дополнение к их латеральным глазам, напоминающим фасеточные глаза насекомых.
Если судить ,по поведению насекомых и результатам электрофизиологических экспериментов, глазки физиологически не равноценны фасеточным глазам и не заменяют последние, а только дополняют их. Мухи, пчелы, муравьи, саранчовые и другие насекомые с закрашенными фасеточными глазами и интактными дорсальными глазками ведут себя как слепые, не ориентируясь даже по обычному источнику света. Вместе с тем закрашивание глазков, хотя бы одного из трех, более или менее существенно нарушает фотокинетические реакции: они замедляются, теряют четкость и требуют для своего осуществления более интенсивного освещения. Поэтому уже давно сложилось мнение, согласно которому глазки регулируют фотокинетические реакции организма, обычно усиливая их при оптимальном или пониженном освещении и тормозя их при чрезмерном освещении. Предполагается, что сигналы с глазков влияют в первую очередь на чувствительность интернейронов фасеточного глаза, хотя стимулирующий эффект глазков замечен даже на сердечной активности насекомого: их освещение вызывает тахикардию у неподвижно сидящей мухи.
В последствии выяснились другие функции глазков. И прежде предполагали, что глазки имеют отношение к осуществлению полета. По крайней мере, была замечена корреляция между бескрылостью и отсутствием глазков в пределах одного и того же таксона. Так, глазков нет у бескрылых форм трипсов, тлей, муравьев, но они имеются у крылатых. У хороших летунов они, как правило, крупнее, чем у слабых летунов. Например, у неравнокрылых стрекоз Anisoptera, в сравнении с медлительными Zygoptera, глазки крупнее, с более сложной сетчаткой, причем медиальный глазок ориентирован вперед, а не вверх, как у равнокрылых стрекоз. Но только теперь электрофизиологическим методом-по импульсной активности в нервной цепочке - обнаружили, что глазки могут служить анализаторами движения крупных объектов, учитывающими направление их движения относительно стрекозы, в частности приближение или удаление. Так, медиальный глазок Sytnpetrutn flaveolum сигнализирует о движении крупного предмета в основном сверху вниз, а латеральные глазки - о движении вверх-вниз.
Причастность глазков к ориентации непосредственно замечена по поведению мух Sarcophaga и нескольких видов шмелей. Эти насекомые с закрашенными фасеточными глазами, но не тронутыми глазками реагируют на поворот плоскости поляризации света. Глазки отличаются от фасеточных глаз намного большей светосилой и могут функционировать даже в сумерках, когда света недостаточно для работы фасеточных глаз. В результате суточный период активности насекомого удлиняется благодаря глазкам. По крайней мере, на перепончатокрылых замечено, что при увеличении размера глазков расширяется диапазон освещенностей, в которых насекомое активно. Различные виды пчел и ос прекращают фуражировку при определенной пороговой интенсивности освещения, которая оказывается тем ниже, чем крупнее глазки, причем виды с глазками более 0,37 мм в диаметре летают даже ночью. Если у медоносной пчелы закрасить дорсальные глазки, то она утром вылетает позднее, а вечером возвращается в улей раньше, чем контрольные интактные пчелы. Следовательно, глазки действительно позволяют насекомому оценивать абсолютную освещенность, что предполагалось исходя из результатов описанных выше электрофизиологических опытов.
Латеральные глазки
Латеральные глазки встречаются у личинок насекомых с полным превращением и отдельных представителей имаго, например у ногохвосток, чешуйниц, блох, самцов червецов и некоторых других. У первых - это временные органы зрения, именуемые стеммами. В процессе метаморфоза стеммы, как правило, дегенерируют, не превращаясь в сложные глаза, и редко переходят имаго в неизменном виде.
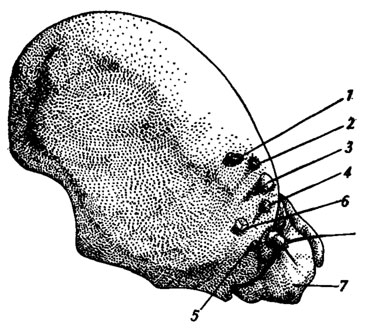
Рис. 65. Расположение стемм на голове гусеницы Isia Isabella (no Dethier, 1942) (вид сбоку): 1,2,6 - тройные глазки; 3,4,5 - одинарные глазки; 7 - ротовой аппарат
Латеральные глазки располагаются по бокам головы в количестве от одного до шести (обычный случай), редко их больше. Если глазков несколько, то не все они развиты в равной мере: обычно два бывают крупнее остальных (рис. 65). Диаметр глазков варьирует у разных насекомых от 50мкм до 0,5 мм, неодинаково и их устройство. Более примитивные из них близки к дорсальным глазкам, а более сложные напоминают своей организацией отдельный омматидий фасеточного глаза. Примером первого типа может служить глазок личинки пилильщика Allantus togatus (рис. 66, А). Под массивным двояковыпуклым линзой-хрусталиком (cornea) лежат корнеагенные клетки (кк), а за ними - чашевидная сетчатка. Дистальные концы зрительных клеток (зк), сгруппированных по четыре в ретинулы, образуют слой рабдомеров. Число зрительных клеток может быть относительно велико - более 6300 в крупных стеммах личинок жуков-скакунов Cicindela.
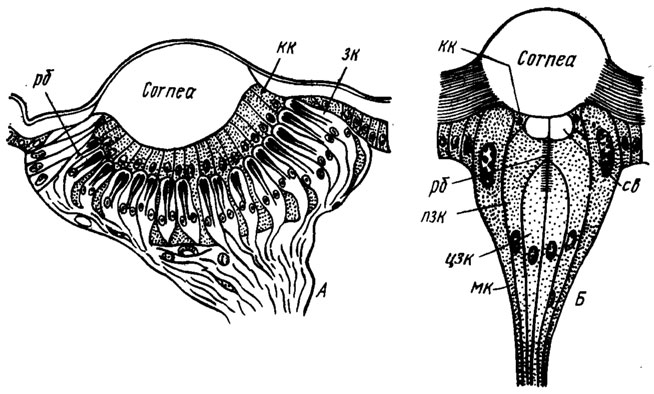
Рис. 66. Латеральные глазки личинок (продольный разрез схематично): А - глазок личинки пилильщика Allantus togatus (по Cornell, 1924); Б - схема глазка гусениц (по Snodgrass, 1924). Св - кристаллическое тело; cornea - роговичиая линза; зк - зрительная клетка; кк - корнеагенные клетки; мк - мантийная клетка; пзк - периферическая зрительная клетка; рб - рабдом; цзк - центральная зрительная клетка
Глазки гусениц бабочек, личинок ручейников, жуков-плавунцов Dytiscus и других построены по типу омматидия: снабжены в дополнение к роговичной линзе кристаллическим телом (конусом), а ретинула сложена небольшим числом зрительных клеток. В глазке гусеницы (рис. 66, Б) позади линзы находится кристаллическое тело (св), окруженное корнеагенными клетками (кк). По бокам от них лежат вытянутые клетки, именуемые мантийными (мк). Последние частично заполнены гранулами темного пигмента и одевают, как футляром или мантией, ретинулу вмеcтe с кристаллическим телом. Ретинула состоит из семи зрительных клеток: трех периферических (пзк) и четырех центральных (цзк), которые все вместе образуют единый осевой рабдом (рб). В аналогична устроенных латеральных глазках личинки муравьиного льва насчитывается до 40-50 удлиненных зрительных клеток.
Оптические свойства и предполагаемая функция глазков гусеницы своеобразны. Роговичная линза и кристаллическое тело функционируют как единая оптическая система. Эта система чрезвычайно короткофокусна (f≈20 мкм), поэтому ее относительное отверстие превышает 1:1,0 и, следовательно, светосила глазка очень велика. Благодаря малой величине фокусного расстояния создаваемое оптической системой изображение незначительно перемещается по глубине, когда объект приближается к глазку или удаляется от него. Поэтому резкое изображение объекта всегда остается в плоскости рабдома, вытянутого вдоль оптической оси. Поле зрения отдельного глазка такого типа, измеренное у личинки муравьиного льва электрофизиологическим способом, равно примерно 47°, а его чувствительность даже под углом 8° к оптической оси не превышает 50% от максимальной.
В глазках с трехраздельной роговицей (рис. 65, 1, 2, 6) образуются соответственно три изображения одной и той же точки. Последнее обстоятельство, а также узкое поле зрения и вытянутый "двухэтажный" рабдом свидетельствуют в пользу того, что отдельный глазок гусеницы не предназначен для восприятия деталей предметов и функционирует подобно одиночному омматидию сложного глаза. Шесть глазков расположены так, что каждый имеет обособленное, практически независимое поле зрения. Поэтому гусеница одновременно видит с каждой стороны головы, очевидно, только шесть независимых точек, каждая из которых есть совокупность всех лучей, попадающих в один глазок. Мозаика этих точек слишком груба, чтобы различать детали и форму предмета. Но малочисленность зрительных единиц, возможно, компенсируется их движением: "привычкой" большинства гусениц раскачивать переднюю часть тела из стороны в сторону и таким способом сканировать окружающие предметы. В разделе о восприятии формы мы уже отмечали, что сканирования поля зрения даже единичным чувствующим элементом в принципе достаточно для оценки визуальной расчлененности окружающей картины.
Стеммы личинок, построенные по типу дорсальных глазков, обладают, вероятно, теми же оптическими свойствами, что и последние. К сожалению, мы вынуждены это только предполагать, так как латеральные глазки личинок и с анатомической и с функциональной стороны изучены незаслуженно мало.
Электрическую реакцию глазка-ЭРГ - отводили только у гусениц бабочек, личинок комара Aedes aegypti и муравьиного льва Euroleon nostras. У первых двух ЭРГ монофазная, роговично-отрицательная, но у муравьиного льва двухфазная: с on- и off-пиками, направленными в противоположные стороны. В ходе темновой адаптации ЭРГ глазка гусеницы тутового шелкопряда увеличивается примерно в течение одного часа. На свету ее амплитуда стабилизируется быстрее: за 8-10 мин. Слияние мельканий наступает уже при 25-30 вспышках в секунду света умеренной яркости, но для личинки муравьиного льва критическая частота мельканий выше - более 40 мельканий в секунду.
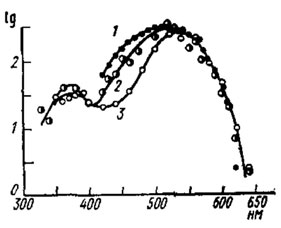
Рис. 67. Кривые дифференциального спектра родопсина (1) и спектральной чувствительности белоглазой (2) и нормальной (3) личинок комара Aedes aegypti (no Seldin, 1972). По оси ординат - логарифмы спектрального поглощения (для 1) или относительной чувствительности (для 2 и 3); по оси абсцисс - длина волны света, нм
В глазках личинки комара Aedes aegypti обнаружен микро-спектрофотометрическим способом зрительный пигмент родопсин с λmах=515 нм и для этих же глазков была измерена по ЭРГ кривая равных спектральных ответов (рис. 67). У белоглазых личинок-мутантов, лишенных экранирующего пигмента, спектральная чувствительность зрительного пигмента хорошо совпадает с кривой равных ответов глазка, из чего можно заключить, что личинка комара цветослепа. Противоположный вывод следует из электрофизиологических измерений спектральной чувствительности глазков гусеницы тутового шелкопряда. Ее кривая равных ответов с пиками в ультрафиолетовом и зеленом участках спектра изменяет свою форму при цветовой адаптации. Глазки гусеницы, по-видимому, имеют два светочувствительных пигмента, что обеспечивает ей возможность цветоразличения.
Этот вывод хорошо согласуется с наблюдением за поведением гусениц дневных бабочек Vanessa и Pieris. В период питания зеленая, желтая и оранжевая окраски привлекают гусениц, а синяя отталкивает, причем обе группы окрасок гусеницы отличают от серых поверхностей разной светлоты. Ко времени окукливания привлекающими становятся темные тона: коричневый и черный. Скорее всего, цветовое зрение имеют и личинки колорадского жука Leptinotarsa decemlineata, глазки которых отличаются высокой контрастной чувствительностью. Зеленую бумагу личинки жука предпочитают серым бумагам, а эти последние с коэффициентами отражения 87 и 93% или 54 и 41% отличают друг от друга.
Показано, что гусеницы бабочек могут различать размер и форму предметов. Из двух равных по ширине вытянутых прямоугольных фигур гусеницы монашенки Lymantria monacha предпочитают более высокую, причем с расстояния 30 см они различают фигуры высотой 9 и 10 см. Сужающаяся кверху фигура привлекает их сильнее фигуры той же высоты и площади, но сужающейся книзу или имеющей параллельные края.
Гусеницы бабочек Malacosoma, Choristoneura, личинка пилильщика Neodiprion реагируют на поворот плоскости поляризации света и могут пользоваться как ориентиром поляризацией света голубого неба.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'