4. Острота зрения. Восприятие формы и объема
Разрешающая способность
Острота зрения, или разрешающая способность, характеризует способность глаза различать степень расчлененности объектов, в том числе отличать две точки от одной более крупной. Измеряют ее тем минимальным расстоянием между двумя объектами, при котором они видны как два раздельных объекта. При прочих равных условиях острота зрения животных определяется в основном степенью расчлененности: 1) зрительного растра, или растра рецептивных единиц, имеющих самостоятельные поля зрения, и 2) оптического изображения на этом растре сетчатки. У позвоночных с их глазами камерного типа рецептивной единицей сетчатки может служить каждая зрительная клетка в отдельности, при этом условии качество ретинального изображения приобретает большое . значение для восприятия деталей внешнего мира. У насекомых, судя по особенностям проекции зрительных клеток омматидиев в первый оптический ганглий, такой рецептивной единицей, воспринимающей расчлененность, служит целый омматидий, а не его отдельные фоторецепторы. Поэтому качество ретинального изображения внутри омматидия несущественно для различения деталей внешнего мира - зрительный центр декодирует поступающую информацию с точностью до числа омматидиев, а не их зрительных клеток. Таким образом, разрешающую способность фасеточного глаза характеризует в первую очередь угловая плотность омматидиев, т. е. число омматидиев в пределах некоторого угла. Чем больше омматидиев приходится на один и тот же угол, т. е. чем ближе сведены их оптические оси, тем выше острота зрения.
Зависимость между угловой плотностью омматидиев и остротой зрения легко понять из следующего рассуждения. Допустим, что на темном фоне имеются две светлые точки. То, что мы видим их как два светлых пятна, разделенных темным промежутком, а не слившихся в одно сплошное большое пятно, - это результат прежде всего различного возбуждения соседних рецепторов сетчатки. Поэтому, чтобы судить, разрешает или не разрешает глаз какие-то объекты, достаточно сравнить интенсивность возбуждения смежных рецепторов этими объектами. Две светлые точки насекомое увидит как две - заметит между ними темный промежуток - только при условии, что ретинальное изображение этих точек, вместе с промежутками между ними, захватит по меньшей мере три омматидия. В таком случае промежуточный омматидий может оказаться возбужденным слабее, чем краевые омматидий, а это и есть условие для отличия двух светлых точек от одной более крупной. Но двух смежных омматидиев недостаточно, чтобы всегда отличать две точки от одной, равной им по суммарной площади.
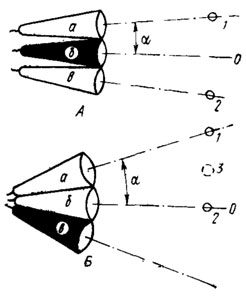
Рис. 52. Зависимость остроты зрения фасеточного глаза от угловой плотности омматидиев (по Мазохину-Поршиякову, 1965). Глаз А разрешает точки 1 и 2, так как они разделены углом, вдвое большим угла а между оптическими осями омматидиев. Глаз Б с меньшей угловой плотностью омматидиев те же точки 1 и 2 не разрешает. Здесь угловое расстояние между точками равно а, поэтому глаз их не отличит от одной более крупной точки. 3. а, б, в - отдельные омматидии; О - оптическая ось омматидиев; одинаково возбужденные омматидии выделены светлым
Соотношение между разрешающей способностью глаза и величиной межомматидийных углов или угловой плотностью омматидиев поясняет рис. 52. Глаз А позволяет разрешить светлые точки 1 и 2 потому, что они возбуждают краевые омматидии а и в сильнее, чем промежуточный омматидии б. Дело в том, что угловое расстояние между точками 1 и 2 вдвое больше угла а между осями смежных омматидиев. Две светлые точки, разобщенные углом 2 а и больше, очевидно, всегда будут возбуждать краевые омматидии не так, как промежуточный, т. е. в принципе всегда могут быть различимы насекомым. Комбинируя положение теста относительно осей омматидиев, легко себе представить, что в некоторых условиях возможно различение точек, разделенных углами менее 2 а, но более а. Глаз Б, у которого углы между осями омматидиев существенно больше, не позволит разрешить те же точки 1 и 2, т. е. не даст возможности насекомому отличить их от одной более крупной точки 3.
Глаза насекомых не обладают аккомодацией: радиус кривизны роговичной линзы у них постоянен. Поэтому для фасеточного глаза существует простая зависимость между расстоянием до объекта и числом различимых деталей: чем ближе объект, тем больше деталей видит насекомое.
Не следует думать, что острота зрения фасеточного глаза может неограниченно возрастать при увеличении угловой плотности омматидиев, т. е. за счет уменьшения диаметра каждого из них. Известный английский исследователь Г. Барлоу (1952) показал, что при малых диаметрах фасеток предел увеличению разрешающей силы глаза кладет дифракция на входном зрачке омматидия и что у перепончатокрылых, в частности у медоносной пчелы, угловая плотность омматидиев в средней части глаза оказалась оптимальной с этой точки зрения. Иначе говоря, в глазу пчелы выдержана оптимальная конструкция: оси омматидиев сближены ровно Настолько, чтобы обеспечить максимальную остроту зрения, допускаемую дифракцией лучей с длиной волны более 300 нм.
Разрешающую способность аппозиционного глаза насекомых, рассчитанную по угловой плотности омматидиев, промоделировал Мазохин-Поршняков (1958) фотографическим путем. В основу модели положены три предпосылки:
- каждый омматидий является рецептивной единицей, посылающей нервный сигнал, независимый от сигналов своих соседей;
- величина сигнала определяется общим количеством света, попадающего во входное отверстие омматидия;
- свет, исходящий от каждой точки поля зрения глаза, воздействует только на один омматидий.
Некоторая схематичность допущений сделана намеренно ради упрощения технического решения задачи. В действительности поля зрения смежных омматидиев частично перекрываются, и между омматидиями существуют взаимотормозные отношения, хотя эти два эффекта в значительной мере, как мы уже знаем, компенсируют друг друга.
Задача исследователя заключав лась в том, чтобы получить на фотобумаге изображение предмета, разбитое на столько отдельных элементов, сколько омматидиев охватывает ретинальное изображение этого предмета. В пределах каждого элемента участок изображения должен иметь равномерную плотность, имитирующую реакцию омматидия на весь попадающий в него свет.
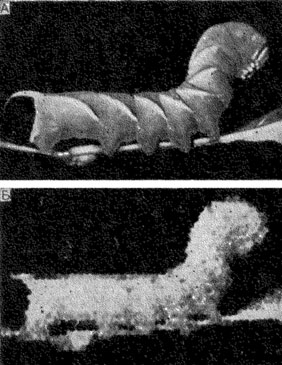
Рис. 53. Гусеница бражника Sphinx ligustri (А) и растровое изображение ее (Б) с теми деталями, которые разрешает глаз стрекозы Aeschna с расстояния 9,5 см (по Мазохину-Поршнякову, 1958)
Для этого негативное изображение предмета проецировали на покрытую молочным стеклом перфорированную пластинку - растр "омматидиев", разбивающую изображение на заданное число элементов. Под перфорированную пластинку помещали фотобумагу, на которой печаталась искомая картина, воспроизводящая степень подробности различения насекомым деталей этого или иного объекта. На рис. 53 изображена гусеница бражника Sphinx ligustri с теми подробностями, которые может заметить насекомое, имеющее глаза аппозиционного и неиросуперпозиционного типов. Это изображение в длину покрывает 45 отверстий растра. Для таких насекомых, как стрекозы Aeschna или мухи Eristalis, у которых углы между осями омматидиев равны примерно 1°, эта фотография воспроизводит гусеницу при рассматривании на расстоянии 9,5 см. Для бабочек Zygaena с вдвое меньшей угловой плотностью омматидиев эта же фотография будет соответствовать случаю, когда они смотрят на гусеницу с вдвое меньшего расстояния, т. е. когда угловой размер гусеницы достигнет 90°.
Ряд исследователей пытались измерить остроту зрения насекомых по их поведению. Например, судя по "критическому" периоду полос вертикального черно-белого узора, нанесенного внутри цилиндра, при котором вращение этого цилиндра перестает вызывать у сидящего внутри насекомого оптомоторную реакцию - компенсационное движение головы или всего тела при смещении поля зрения, - максимальная острота зрения действительно хорошо совпадает с величиной 2α. Именно для медоносной пчелы, мухи Calliphora erythrocephala, жука Chlorophanus критический период узора равен соответственно 1,96°; 4°; 14°, а удвоенный угол - между осями смежных омматидиев равен у этих насекомых соответственно 1,8°; 4,8°; 14°.
Личинки палочника Dixippus morosus стремятся "вползти" на нарисованные на бумаге вертикальные черные полосы, признавая их за стебли растений. Если полосы нарисованы слишком часто, то насекомые перестают на них реагировать. Оказалось, что минимальный угловой период полос, при котором они привлекают палочников, приблизительно вдвое больше межомматидийного угла в глазу насекомого, т. е. тоже равен около 2α.
Нередко даже опытные исследователи оценивают максимальную остроту зрения по минимальному размеру видимых предметов, незаконно подменяя одно понятие другим. Чтобы увидеть маленький предмет, т. е. отличить присутствие предмета в поле зрения от его отсутствия, совсем не требуется высокая разрешающая способность глаза, но важно, чтобы он имел достаточно высокую контрастную чувствительность. Поясним эту мысль на примере охотящейся стрекозы. Она замечает муху с расстояния в несколько метров, когда угловой размер добычи в несколько раз меньше межомматидийного угла α. Иными словами, стрекоза видит муху с такого расстояния, с которого она уже не может отличить двух близко расположенных мух от одного более крупного насекомого. Появление темной точки в поле зрения единственного омматидия уменьшает количество падающего на него света, и этого достаточно, чтобы отличить случай присутствия точки от случая ее отсутствия. При этом точка будет видна тем дальше, чем выше контрастная чувствительность фоторецепторов и чем больше контраст в яркости между точкой и фоном, на котором она находится. Таким образом, чтобы увидеть сколь угодно маленький предмет, достаточно одного рецептора, но чтобы отличить его от другого маленького предмета, требуется минимум три рецептора.
Разрешающая способность глаза в принципе зависит также от контрастной чувствительности, величины поля зрения и характера нервного взаимодействия его рецептивных единиц. Чем выше контрастная чувствительность зрительного растра, тем лучше острота зрения, так как тем больше перепадов освещенности заметит насекомое. Мы уже отмечали, что поля зрения соседних омматидиев частично перекрываются. Это может снизить потенциальную остроту зрения при данной угловой плотности омматидиев, поскольку наложение друг на друга полей зрения ведет к сглаживанию различий в состоянии возбуждения соседних омматидиев. Однако мы знаем, что в оптическом ганглии насекомого имеется механизм подчеркивания контуров, который функционально компенсирует отрицательный эффект наложения полей зрения у соседних омматидиев.
До сих пор мы подразумевали, что глаз замечает расчлененность окружающей картины в результате неодинакового возбуждения разных омматидиев одновременно, т. е. зрительный анализатор оценивает расчлененность по различию сигналов от разных клеток сетчатки. Возможен и другой способ восприятия расчлененности, когда последовательно, в разные моменты времени, сравнивается возбуждение одних и тех же зрительных единиц. Глаз, построенный на таком принципе, смог бы различать на движущейся картине даже больше деталей, чем это обеспечивает угловая плотность зрительных единиц. В зрительном анализаторе человека и, видимо, всех позвоночных животных этот принцип не реализован, но у насекомых, некоторых паукообразных и ракообразных скорее всего есть зрительный механизм, основанный на трансформации пространственной последовательности стимулов во временную последовательность возбуждения одних и тех же рецепторных единиц. Известный немецкий физиолог Г. Аутрум показал, что фасеточный глаз быстро летающих насекомых, для которых особенно важно различать предметы в движении, отвечает тем требованиям, которым должен удовлетворять обсуждаемый гипотетический зрительный механизм. Именно восприятие расчлененности на движущейся картине как последовательности мельканий возможно при условии малой инерционности сетчатки и ее высокой контрастной чувствительности. Как показали электрофизиологические измерения критической частоты мельканий и прямое наблюдение за поведением насекомых в оптомоторном барабане с периодическим узором полос, для хороших летунов - мух, пчелиных, стрекоз-мелькания не сливаются до тех пор, пока их частота станет равной 250-300 в секунду (см. рис. 48, в, г). Вспомним, что человек не видит смены кадров на киноэкране, следующих с частотой уже 16 в секунду. Инерционность нашего глаза намного ниже, чем у быстро летающих насекомых. О высокой контрастной чувствительности фасеточного глаза уже говорилось - она достигает 1 %, что достаточно для видения при движении.
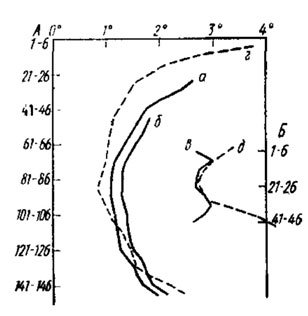
Рис. 54. Величина углов между оптическими осями омматидиев рабочей пчелы Apis mellifera в вертикальном (а, б, г) и горизонтальном (в, д) направлениях. а,б,в - результаты измерения del Portillo (1936); г, д - результаты измерения Baumgarten (1928). Измерения получены разными способами. По оси ординат - порядковые номера омматидиев сверху вниз (А) и спереди назад (Б); по оси абсцисс - величина углов
Еще одна особенность фасеточного глаза явно возникла как приспособление для видения в полете. У ряда подвижных насекомых, таких, как мухи и пчелиные, межомматидийные углы по горизонтальному диаметру глаза крупнее, чем по вертикальному. Дело в том, что для различения деталей во время движения глазу достаточно небольшого числа омматидиев, вплоть до одного, и явно невыгодна большая угловая плотность зрительных единиц, так как при этом освещенность соседних омматидиев будет изменяться (мелькать) слишком быстро для того, чтобы рецепторы успевали реагировать на такие мелькания. Поэтому в направлении смены объектов в поле зрения при движении (по горизонтали) межомматидийные углы у хороших летунов в два-три раза шире, чем по вертикали (рис. 54).
Прямым свидетельством реализации у животных принципа зрения в движении можно считать необычное устройство глаз у некоторых паукообразных и ракообразных. Например, в глазу морского рачка Сорiliа позади сравнительно крупной линзы лежит всего четыре зрительные клетки, но они подвижны за счет ритмических сокращений специальных мышц. Такая сетчатка из четырех элементов непрерывно колеблется, сканируя ретинальное изображение с частотой 5-6 раз в секунду. Малое число рецепторов здесь компенсируется их подвижностью, что, очевидно, позволяет рачку различать детали и форму предметов. Сканирующая сетчатка известна также в глазах некоторых пауков и клещей. Недавно описана особая мышца в глазах мух, периодически подергивающая проксимальную часть слоя рецепторов.
Человек не видит предметы, если их изображение неподвижно относительно сетчатки. Но стабилизация ретинального изображения осуществима только в эксперименте, а в реальных условиях глаз непрерывно совершает движения: произвольные скачки - смену точек фиксации, непроизвольные скачки, дрейф, тремор и другие, препятствующие возникновению, как говорят, "пустого" поля. Не исключено, что мышца в фасеточном глазу мух служит именно тому, чтобы обеспечить восприятие, периодически двигая рецепторы относительно сетчаточного изображения, поскольку сами глаза неподвижны относительно головы.
Существует любопытная и, возможно, глубокая по своему физиологическому смыслу аналогия между движениями глаз человека при рассматривании предметов и движениями головы у хищных мух-ктырей во время визуального выслеживания добычи. Человек не может плавно следовать взором от одной точки предмета к другой - глаза непроизвольно и очень быстро, за сотые доли секунды, скачком переходят из одного положения в другое. Киносъемка охотящегося ктыря показала, что и его голова очень быстрыми скачками переходит из одного положения в другое, когда муха обращает внимание на новые объекты. Но и человек и ктырь плавным движением (первый - глаз, второй - всей головы) прослеживают движущийся предмет; скачки в этом случае никогда не наблюдаются.
Различение формы и восприятие объема
До последнего времени исследователи связывали восприятие формы насекомыми исключительно с движением ретинального изображения относительно рецепторов, когда графические (пространственные) свойства предметов могут восприниматься как последовательность мельканий или последовательность возбуждения во времени одних и тех же омматидиев. Эта гипотеза возникла на основе следующих наблюдений. Пчелиных, бабочек, мух и других антофильных насекомых расчлененные фигуры спонтанно привлекают сильнее фигур с ровным сплошным контуром или однородным (нерасчлененным) внутренним полем. Колеблющиеся на ветру цветки насекомые посещают чаще, чем неподвижные цветки. Но самое главное доказательство состоит в том, что никому не удавалось обучить пчел различать фигуры с простым контуром, в частности отличать треугольник от квадрата или круга равной площади. Однако в тех же опытах пчелы различали, например, лучистые фигуры: 8- и 16-лучевые звезды, а также отличали крест и звезду от круга. "Решают" пчелы и более сложные задачи на опознавание, явно руководствуясь статистическими признаками рисунка фигур - повторяемостью одних и тех же сочетаний, например светлого и темного, - если это черный рисунок на белом фоне. После соответствующего научения пчелы в опытах Л. И. Францевича (1968) различали стандартного размера черно-белые случайные рисунки, предлагаемые по специальной программе компьютером, если они относились к разным статистическим классам в указанном выше смысле. Поэтому многие авторы утверждали, что насекомые могут различать фигуры только по степени расчлененности их контура или внутреннего поля, т. е. путем сравнения частоты мельканий, возникающих в глазу при движении насекомого относительно объекта или самого объекта относительно глаза. В действительности дело обстоит иначе. Пчелы при длительном обучении все же способны отличать, например, треугольник от квадрата, особенно если контур фигур многократно повторен, что позволяет насекомому оценивать форму как издалека, так и вблизи, когда в поле зрения помещается только часть ее. Опознать эти фигуры по частоте и последовательности мельканий практически невозможно. Следовательно, пчелы могут воспринимать пространственные отношения предметов или элементов рисунка (видеть форму), в том числе и по картине возбуждения разных рецепторов одновременно. Из этого следует, что насекомым свойственны оба рассмотренных выше механизма анализа расчлененности поля зрения.
Насекомые различают предметы по размеру. Например, пчелы различают два круга при условии различия их площадей в 2 раза и больше; но осы и мухи делают это более тонко: различие становится возможным уже при условии отношения площадей фигур не менее чем 1 : 1,5.
Насекомые, отличающиеся быстрым полетом или хорошо прыгающие, а также большинство хищников воспринимают также объемность предметов и хорошо оценивают глубину пространства. Стереоскопичность зрения достигается тем, что поля зрения обоих глаз впереди и внизу частично перекрываются: в правом и левом глазах есть омматидии, оптические оси которых сведены или хотя бы параллельны друг другу. Такие (корреспондирующие) пары омматидиев "видят" предмет в одной и той же точке пространства. Для разных точек пространства по мере удаления от глаза Корреспондирующими окажутся разные пары омматидиев, что в принципе достаточно для улавливания изменения глубины. Корреспондирующие омматидии, например, у богомолов лежат в специальной зоне глаза причудливой формы, одностороннее ослепление которой ведет к утрате способности насекомого безошибочно схватывать добычу. В норме же богомол, как и охотящаяся личинка стрекозы, никогда не выбрасывает ловчий аппарат, когда расстояние до жертвы больше последнего. Стереоскопичность зрения важна не только охотникам за добычей, но и всем быстро летающим и быстро бегающим формам для обеспечения посадки, своевременного маневра при облете препятствий или преследования полового партнера. В случае посадки сигнал для своевременного вытягивания ног посылают особые детекторы, описанные на примере зрительной системы мух.
В заключение этого раздела обратим внимание на функциональную неоднородность фасеточного глаза, четко проявляющуюся в ряде отношений в поведении насекомого, но не всегда морфологически ясно выраженную. Так, взрослая стрекоза Libellula quadrimaculata различает цвета только с помощью вентральной половины глаза, сложенной более мелкими зеленоватыми фасетками. Дорсальная половина глаза цветослепа, но обладает высокой чувствительностью к коротковолновым лучам: ультрафиолетовым, фиолетовым и синим, что облегчает стрекозе замечать мелкую добычу на фоне неба. В глазу богомола Stogmatoptera описана особая зона с повышенной остротой зрения. Эта так называемая фовеа, как уже говорилось выше, необходима и достаточна для точной оценки дистанции броска на добычу. У водомерок Gerris только дорсальная половина глаза с особым расположением микровилл в рабдомерах пригодна для анализа направления поляризации света и астрономической ориентации. У медоносной пчелы не замечено резких морфологических различий между верхней и нижней половинами глаза, но функционально они не равноценны. Пчела различает цвет обеими половинами глаза; но для опознавания объектов по форме и размеру пригодна в основном нижняя половина, а для анализа поляризации служит исключительно верхняя половина глаза.
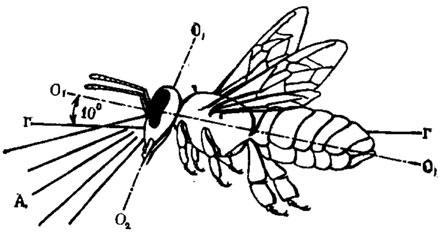
Рис. 55. Ориентация тела медоносной пчелы, летящей над горизонтальной поверхностью (по Мазохииу-Поршнякову, Таимовой, 1973): А - зона глаза, которой пчела смотрит на близлежащие горизонтальные предметы; O1-O1 - медиальная ось тела; О2-О2 - продольная ось головы; Г-Г - линия горизонта
Во время полета над горизонтальной поверхностью пчела удерживает медиальную ось тела примерно на 10° выше линии горизонта. Благодаря этому, а также постоянному наклону головы ротовыми частями несколько вперед она смотрит вниз вентральной третью поля зрения (рис. 55, А), как раз приспособленной для опознавания предметов по форме, размеру и цвету. Что же касается медиальной и дорсальной третей глаза, направленных в полете в основном вверх, то они предназначены для измерения и удерживания менотаксических углов ориентации по солнцу и поляризации света голубого неба и, конечно, для общего обзора.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'