3. Контрастная чувствительность, адаптация и ретиномоторные явления
Для нормального видения, позволяющего различать многообразие окружающих предметов, глаз должен иметь высокую контрастную чувствительность. Контрастную чувствительность сетчатки измеряют как ΔI/I o 100 %, где ΔI - минимальное визуально контролируемое изменение интенсивности света, а I - исходная интенсивность света. У хорошо видящих животных, в том числе у насекомых, она достигает 5% и даже 1%. Это значит, что, например, пчелы или мухи замечают разницу в интенсивности света на 5% и меньше. Общая же освещенность в естественных условиях изменяется в громадном диапазоне: от 100 тыс. л к в летний полдень до 0,2 лк при полной луне и 0,0005 лк безлунной ночью. Как же обеспечивается в зрительной системе высокая контрастная чувствительность при работе в условиях непостоянной освещенности, изменяющейся в миллионы раз?
Прежде всего нужно учесть, что в природе животных окружают почти всегда несамосветящиеся объекты, которые только отражают свет, падающий извне. Но такие объекты отличаются по интенсивности отражения не более, чем в 20-30 раз, и именно такой сравнительно узкий перепад яркостей должен замечать их глаз. Последнее достигается тем, что амплитудные характеристики реакции фоторецепторов приходятся наиболее крутой частью как раз на такой узкий перепад интенсивностей света. Для этого необходимо, чтобы при изменении общего уровня освещения количество поглощаемых рецепторами фотонов изменялось бы по возможности мало. В таком случае амплитудные характеристики будут как бы сдвигаться, подстраиваясь своей крутой частью к данному уровню освещения. Разнообразные опыты показали, что именно так обстоит дело и в глазу камерного типа позвоночных, и в совершенно иначе устроенном фасеточном глазу насекомых.
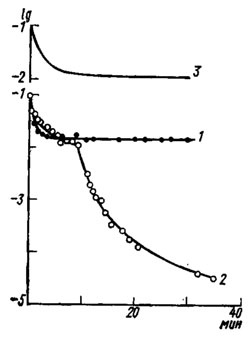
Рис. 51. Кривые изменения чувствительности глаза ночной бабочки Cerapterix graminis (1, 2) и дневной бабочки Argynnis adippe (3) в ходе темновой адаптации (по Bernhard, Ottoson, 1960): 1 - случай, когда ирисовый пигмент не перемещался; 2 - при перемещении пигмента. По оси ординат - логарифмы пороговой интенсивности света; по оси абсцисс - время, мин
Регуляция количества света, поглощаемого рецепторами, достигается благодаря согласованной работе нескольких механизмов зрительной адаптации, или приспособления глаза для функционирования при широко меняющейся освещенности. О состоянии адаптации обычно судят по пороговой энергии света, достаточной для появления электрического ответа глаза (ЭРГ) или моторной реакции целого организма. Образец кривых изменения пороговой чувствительности глаза в процессе темновои адаптации представлен на рис. 51. У дневных насекомых с аппозиционными (Argynnis) и нейросуперпозиционными (Calliphora) глазами чувствительность глаза в темноте возрастает только в десятки раз, а у ночных, например у бабочек совок или пядениц, она увеличивается на 4-5 десятичных логарифмических порядков. У первых - это однофазный и короткий процесс, у вторых - двухфазный и продолжительный. Начальные 8-10 мин темновая адаптация идет и у тех и у других практически одинаково, но затем чувствительность ночных видов с оптикосуперпозиционными глазами круто повышается: наступает вторая фаза адаптации, а у дневных видов светочувствительность сохраняется на прежнем уровне. Гистологический контроль положения ирисового защитного пигмента в омматидиях показал, что перегиб адаптационной кривой ночных бабочек хорошо совпадает по времени с началом перехода этого пигмента в положение, характерное для ночи. То и другое наступает в среднем на 6-8 мин темновои адаптации, и оба эти процесса заканчиваются примерно за 30-50 мин.
Световая адаптация глаза, подстройка его чувствительности для работы при ярком освещении, идет гораздо быстрее.
Таким образом, к адаптации глаза причастны ретиномоторные явления, одним из примеров которых служит перемещение гранул защитного пигмента в ирисовых пигментных клетках оптикосуперпозиционного глаза. Регуляция доступа света к фоторецепторам в данном случае целиком механическая. Под действием света или утром, в силу внутреннего физиологического ритма, или "часов", насекомого, гранулы пигмента мигрируют проксимально, что по результату аналогично сужению входного зрачка. Темнота или наступление вечера стимулируют обратный процесс: гранулы пигмента собираются у дистального конца клеток, благодаря чему больше лучей света достигает рабдома каждого омматидия.
Миграции экранирующего пигмента в ирисовых и других пигментных клетках нередко сопутствует изменение длины рабдома. Под действием избытка света он укорачивается, что ухудшает условия его освещения. В темноте и при низкой освещенности рабдом медленно (часами) удлиняется, как бы тянется к свету. Адаптационные изменения длины рабдома замечены в секомых.
Но как объяснить быстрые изменения чувствительности глаза дневных насекомых? Известны, по крайней мере, два механизма быстрой адаптации глаза. Первый из них связан с фотохимическими превращениями зрительного пигмента в рецепторах. Под действием света концентрация исходного зрительного пигмента падает, и, следовательно, уменьшается вероятность встречи фотонов с неразложенными молекулами исходного фотопигмента. В темноте идет ресинтез фотопигмента, что и обеспечивает повыше ние светочувствительности рецептора. Первая (быстрая и короткая) фаза темновой адаптации оптикосуперпозиционного глаза, возможно, отражает именно этот фотохимический процесс в сетчатке.
Второй механизм адаптационной подстройки чувствительности глаза основан на изменении волноводных свойств рабдома, регулирующих в конечном счете угловое поле зрения рецептора. Так, у саранчи Locusta migratoria при световой адаптации палисад из вакуолей эндоплазматического ретикулюма вокруг рабдома распадается, и вместо него здесь скапливаются митохондрии, коэффициент преломления которых ближе к таковому рабдома, чем у палисада. В результате ухудшаются условия светопроведения, меньше света удерживается рабдомом при многократном отражении лучей на границе с митохондриями, а поле зрения рабдома при этом сужается. В итоге падает уровень светочувствительности глаза. И наоборот, образование палисада в темноте повышает светочувствительность глаза, поскольку уменьшается угол полного внутреннего отражения рабдома, что приводит к расширению его поля зрения.
Сходное явление описано у мухи Musca domestica. Оно наблюдалось за счет миграции гранул экранирующего пигмента в цитоплазме зрительных клеток. При освещении глаза гранулы экранирующего пигмента быстро, в течение нескольких секунд, мигрируют к рабдомерам, вследствие чего в последних нарушаются условия для полного внутреннего отражения и свет частично рассеивается, частично поглощается экранирующим пигментом. Это влечет за собой сужение поля зрения рецепторов, что показано по уменьшению угла, в пределах которого свет вызывает электрический ответ зрительной клетки. Таким способом фоторецепторы мухи защищаются от избытка света. Регуляция осуществляется, по-видимому, на основе петли обратной связи между интенсивностью возбуждения зрительной клетки и количеством мигрирующего в ней защитного пигмента, т. е. она похожа по своей сущности на так называемый "зрачковый рефлекс" камерного глаза позвоночных.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'