2. Оптические ганглии и механизмы предварительной обработки зрительной информации
Оптические ганглии и типы их интернейронов
Зрительный центр фасеточных глаз находится в парных оптических (зрительных) долях, представляющих собой боковые части переднего мозга. В каждой оптической доле различают четыре отдела:
- ламину (I оптический ганглий), обычно непосредственно подстилающую дно глаза;
- медуллю (II оптический ганглий), связанную с ламиной внешней хиазмой;
- лобулю и
- лобулярную пластинку, или лобулюс (рис. 46).
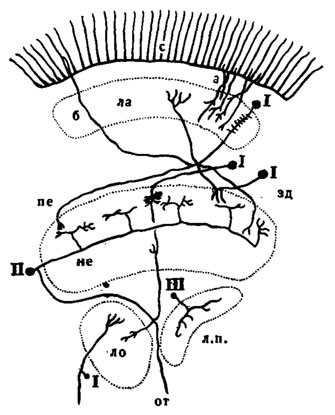
Рис. 46. Морфологические типы нейронов в оптической доле переднего мозга (по Collett, 1970): I - перпендикулярный нейрон; II - тангенциальный нейрон; III - амакриновая клетка. а - короткий рецепторный аксон; б - длинный рецепторный аксон; с - сетчатка; зд - задний край; ла - ламина; ло - лобуля; л.п. - лобулярная пластинка; ме - медулля; от - оптический тракт, ведущий к медиальной части протоцеребрума; пе - передний край
Два последних отдела, связанных с медуллей внутренней хиазмой, ранее объединяли под названием "III оптический ганглий". Все отделы представляют собой чрезвычайно сложные нервные массы, образованные большим числом нейронов различного вида ,и назначения. Нейроны подразделяют на три основных класса: перпендикулярные (рис. 46,1), тангенциальные (II) и амакриновые клетки (III). Перпендикулярные нейроны, соединяющие друг с другом разные нервные массы (отделы), отличаются тем, что их аксоны располагаются более или менее перпендикулярно к поверхности ганглия, в котором они лежат. Тангенциальные нейроны связывают по горизонтали параллельные ряды перпендикулярных клеток, а также образуют прямую связь между ипси- и контралатеральны-ми оптическими долями. Амакриновые клетки - это нейроны, отростки которых не выходят за пределы одного отдела оптической доли. Они встречаются в разных ганглиях и имеют окончания на тех же уровнях, что и нейроны остальных классов.
В дистальной части ламины плотно лежат друг возле Друга так называемые "оптические патроны". В каждом оптическом патроне сходятся в виде пучка обычно шесть коротких волокон зрительных клеток, синаптически контактирующих здесь с двумя-тремя перпендикулярными клетками - нейронами второго порядка, а также с несколькими центробежными волокнами. Число оптических патронов в ламине равно числу омматидиев.
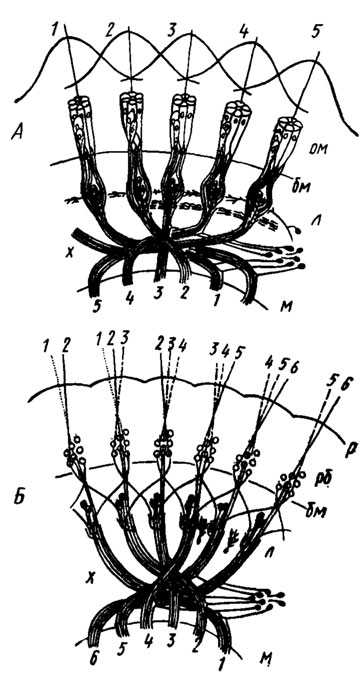
Рис. 47. Проекция сетчатки с замкнутым (А) и открытым (Б) рабдомом на оптическую долю (по Horridge Meinertzhagen, 1970). На А: 1, 2, 3, 4, 5 - поля зрения отдельных омматидиев и их проекция в медуллю; на Б; 1, 2, 3, 4, 5 - оптические оси отдельных рабдомеров и проекция последних в медуллю. бм - базальная мембрана; л - ламина; м - медулля; ом - омматидии; р - роговица; рб - рабдомеры; х - хиазма
Соответственно двум типам рабдома описаны два типа проекции растра сетчатки на ламину. У насекомых с замкнутым рабдомом в каждом оптическом патроне сходятся аксоны одного и того же омматидия, т. е. наблюдается полная и прямая проекция короткоаксонных ретинальных клеток каждого омматидия на I оптический ганглий (рис. 47, А). У мух и, вероятно, у других насекомых с открытым рабдомом в каждом омматидии сходятся аксоны группы смежных омматидиев, именно тех ретинальных клеток, рабдомеры которых смотрят в одну и ту же точку пространства (рис. 47, Б).
Длинные волокна, принадлежащие базальным клеткам омматидия, проходят ламину насквозь, затем вместе с перпендикулярными волокнами вступают в наружную хиазму и оканчиваются в виде пучков в медулле. За счет этих пучков возникает столбчатость медулли. Проекция оптических патронов ламины на медуллю полная, но зеркально повернутая в вертикальной плоскости.
Для наглядности можно записать, что некоторой последовательности омматидиев - 1, 2, 3, 4, 5 соответствует в ламине тот же ряд оптических патронов, а в медулле он переходит в преобразованный ряд - 5, 4, 3, 2, 1 (рис. 47, А, Б).
Связь первых двух оптических ганглиев с третьим еще сложнее и пока недостаточно изучена. Полагают, что имеется не менее шести раздельных путей проекции сетчатки на третий оптический ганглий. Что касается более высоких связей, то оптические волокна, судя по данным электрофизиологических опытов, прослеживаются в средней части протоцеребрума и в его грибовидных телах.
В настоящее время точная картина нервных связей в зрительном центре усиленно изучается многими исследователями-морфологами совместно с электрофизиологами. Раскрытие ее даст ключ к пониманию механизмов обработки зрительной информации, к чему так стремятся конструкторы "думающих" машин.
Электрическая реакция глаза и передача сигналов зрительной клеткой
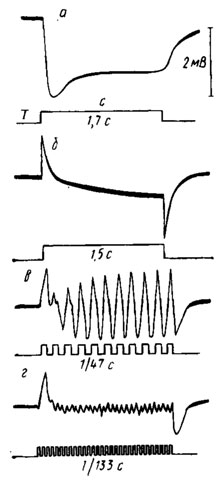
Рис. 48. Электроретипограммы: а - комара Anopheles maculipennis (по Мазохину-Поршнякову, 1965); б, в, г - мухи Calliphora erythrocephala (по Autrum, 1958, видоизменено). ЭРГ на в, г получены при мелькающем освещении. Нижние кривые - отметки раздражения; частота стимуляции обозначена на рисунке; с - свет; т - темнота
Под действием света в зрительной клетке возникает характерный рецепторныи потенциал, имеющий положительный знак при внутриклеточном отведении (см. рис. 4,А,Б). Совокупность этих рецепторных потенциалов возбужденной сетчатки образует основную часть ЭРГ, регистрируемой на роговице в виде медленного столообразного отрицательного потенциала, сохраняющегося в течение всего периода засвета глаза (рис. 48, а). Другой (быстрый) компонент двухфазной ЭРГ, наблюдаемый у мух, бабочек, стрекоз и некоторых других насекомых, имеет ганглиозное происхождение (рис. 48, б). После удаления оптических ганглиев или выключения их стрихнином и другими агентами двухфазная ЭРГ с быстрыми on- и оff-пиками превращается в медленную монофазную отрицательную ЭРГ обычного вида.
Возбуждение зрительной клетки передается по ее аксону, по-видимому, электротонически, а не в форме дискретных спайков, как у хеморецепторных и большинства других чувствующих клеток насекомых. Разные авторы при тщательном специальном поиске не нашли внутриклеточных импульсов в фоторецепторах стрекоз, прямокрылых, мух, рабочих медоносных пчел. Только у трутня медоносной пчелы видели одиночный спайк, накладывающийся на фазу нарастания медленного деполяризационного потенциала зрительной клетки.
У мечехвоста Limulus эксцентрическая клетка разряжается залпом импульсов, легко отводимым внутри- и внеклеточно. Зрительный центр у него удален от глаза, и связь омматидиев с ним осуществляет пучок длинных рецепторных аксонов. У насекомых оптические ганглии сближены с рецепторами глаза, аксоны ретинулярных клеток короткие (например у саранчи перелетной - 200 мкм), поэтому неудивительно, что им свойственна безымпульсная передача сигналов сетчатки. Пока остается неизвестным, по аксонам каких клеток омматидия и каким образом передается информация одновременно о яркости, цвете и поляризации стимула. Ясно одно: визуальная информация о внешнем мире поступает в мозг с точностью до растра омматидиев. Она может передаваться по двум разным каналам: коротким аксонам зрительных клеток и более длинному волокну базальной клетки. Если сигналы всех зрительных клеток одного омматидия суммируются на первом же интернейроне в ламине, то базальная клетка может передавать свой сигнал непосредственно в медуллю.
Детектирование признаков и переработка информации в зрительном центре
В оптических ганглиях происходит сложная обработка зрительной информации, поступающей от сетчатки глаза. Некоторые механизмы этой обработки уже выявлены, другие - интенсивно изучаются.

Рис. 49. Импульсная активность нейронов-детекторов направления движения в зрительной системе стрекозы Aeschna viridis (по Зенкину, Пигареву, 1969): а, б - реакция нейрона на движение соответственно черной или белой полосы снизу вверх; в - реакция того же нейрона на выключение света; г - реакция нейрона 'с привыканием' на движение темного пятна диаметром 3° (вторая стимуляция следовала через 20 с после первой); д - реакция off-элемента (детектора затемнения) на выключение света; е, ж - реакции предыдущего нейрона на движение темной полосы к центру и от центра рецептивного поля глаза; з - реакция off-элемента на изменение освещения. Стрелками показано направление движения тестовых объектов; калибровка-100 мкв; отметка времени - 0,5 с
Сначала у позвоночных, затем у насекомых были открыты тангенциальные ганглиозные нейроны, избирательно реагирующие на определенные параметры зрительного стимула: размер, характер движения, цвет и т. п. (рис. 49). Такие, иногда чрезвычайно узкоспециализированные интернейроны получили название детекторов. Детекторы возникли у животных как способ экономного описания внешнего мира и передачи в высшие ассоциативные центры только существенной для опознания объектов информации, отфильтрованной от ее избытка. Нет сомнения в том, что у низших животных, у насекомых в особенности, сигналов некоторых видов детекторов достаточно для запуска врожденных стереотипных реакций.
У мух, бабочек, жуков во II и III оптических ганглиях, а также в средней части переднего мозга при электрофизиологических исследованиях обнаружено много видов зрительных детекторов с разнообразными свойствами. Их рецептивные поля варьируют от 1-6° до размера поля зрения целого глаза и могут быть как монокулярными, так и бинокулярными. В последнем случае одновременная стимуляция левого и правого глаз нередко вызывает в нейронах реципрокно-тормозной ответ. Одни детекторы длительно реагируют на продолжительную стимуляцию, другие обладают "привыканием", но наиболее замечательное свойство большинства их - дирекциональная (направленная) чувствительность к движению: реакция в основном или исключительно на те стимулы, которые движутся в строго определенном направлении (рис. 49, а, б, в). Движение в "предпочитаемом" направлении возбуждает нейрон, а в прямо противоположном подавляет его фоновую активность.
Простейшими среди зрительных детекторов являются нейроны "тревоги", реагирующие только на затемнение части глаза или на начало движения объекта в любом направлении. Импульсации клеток такого рода, сигнализирующих о всяких изменениях в поле зрения, в принципе достаточно для своевременного запуска реакции убегания (рис. 49, г, д, е).
Более сложны и интересны детекторы направления движения. Они избирательно разряжаются импульсами при движении объекта сверху вниз или снизу вверх, слева направо или справа налево, в ростральном или каудальном направлениях и т. д. Понятно, что на основе набора подобных нейронов с их бинокулярными перекрестно-тормозными связями легко организовать детектирование и автоматический контроль продольного и поперечного крена в полете и при плавании. Благодаря им же тело насекомого легко удерживается в определенной точке пространства, например при сносе течением, а также создается механизм отличения собственного движения от смещения фонового окружения. Бинокулярные нейроны, чувствительные либо к ростральному, либо к каудальному движениию, лежат в основе управления оптомоторной реакцией, служащей для компенсации отклонений от прямолинейного курса движения. Свойства этих нейронов у пластинчатоусых жуков хорошо совпадают с характеристиками их оптомоторной реакции, как например, у навозных жуков Geotrupes.
Из визуальных детекторов пищи примечательны детекторы небольшого движущегося пятна, первоначально описанные у лягушек, а затем у стрекоз. У имаго Aeschna - это клетки с "привыканием", реагирующие на движение только маленьких (диаметром 1-6°) темных или светлых пятен. Детекторы движущегося пятна имеют прямое отношение к организации пищевого поведения: с их помощью насекомые опознают мелкую добычу, и, кроме того, видимо, осуществляется запуск реакции преследования у охотящейся стрекозы.
Зрительные детекторы позволяют опознавать по окраске особей своего вида. Одним из примеров могут служить мозговые интернейроны тропической Morpho amathonte, возбуждаемые движущимися стимулами голубого цвета и тормозимые стимулами с большей длиной волны. Вся зрительная система этой атласно-голубой бабочки хорошо приспособлена для обслуживания брачного поведения, причем названные детекторы специально настроены на опознавание полового партнера. В какой мере зрение управляет по-ведением бабочек-морфид, можно судить по распространенному в тропиках способу их привлечения и лова: берут голубую блестящую пластинку или зеркало и этим предметом размахивают в воздухе.
Таким образом, ганглиозные нейроны-детекторы обеспечивают прежде всего механизм выделения признаков, необходимых, а иногда и достаточных для опознавания определенных объектов. Кроме того, они являются датчиками информации, достаточной для формирования целесообразной моторной команды, т. е. для запуска поведенческого ответа и наведения организма на цель на основе алгоритма одного из таксисов (см. ниже).
Электрофизиологические исследования позволили выявить в глазах животных другой очень интересный механизм предварительной обработки зрительных сигналов. Речь идет о так называемом "латеральном торможении" нейронов, первоначально описанном и подробно изученном в фасеточном глазу мечехвоста Limulus Нобелевским лауреатом X. Хартляйном, а затем обнаруженном в сетчатке позвоночных и оптическом ганглии насекомых.
Регистрируя импульсную реакцию пучка аксонов от одиночного омматидия мечехвоста, идентичную электрическому ответу аксона отдельной зрительной клетки, X. Хартляйн обратил внимание на то, что ответ омматидия зависит не только от возбуждающего стимула, но и от активности соседних омматидиев. Рецептивные единицы мечехвоста оказались взаимозависимыми, при этом взаимодействие между ними было строго тормозным: каждый возбужденный омматидий тормозил соседние омматидии и в то же время сам тормозился ими. Торможение было тем больше, чем сильнее возбуждался омматидий. Оно передавалось через сплетение тончайших боковых веточек аксонов рецепторных клеток - нейропиль - проксимальнее базальной мембраны. У насекомых, как полагают, взаимотормозные отношения существуют между оптическими патронами ламины.
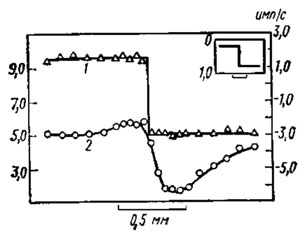
Рис. 50. Частота импульсов в пучке волокон отдельного омматидия при освещении ступенчатым яркостным узором глаза мечехвоста (по Ratliff, Hartline, 1959). На разные участки глаза последовательно проецировали с помощью полупрозрачной маски прямоугольную яркостную ступеньку с соотношением интенсивности левого и правого полей, равным 4: 1 (вставка в верхнем правом углу; по оси ординат - оптическая плотность маски). 1 - изменение возбуждения исследуемого омматидия при различных положениях яркостной ступеньки, когда освещен только этот омматидий, а все остальные затемнены непрозрачным экраном. Кривая возбуждения полностью воспроизводит (в логарифмическом масштабе) ступенчатый перепад в интенсивности освещения, т. е. в отсутствие торможения. Отдельные точки - частота импульсов (цифры справа) тестового рецептора при освещении яркостной ступенькой. 2 - изменение возбуждения (частота импульсов - цифры слева) того же омматидия и для тех же положений яркостной ступеньки после снятия экрана. В этих условиях был освещен весь глаз, и в результате торможения рецепторов кривая возбуждения тестового омматидия резко отличается от той, которая получена в предыдущем опыте. Масштабная черта по абсциссе равняется 0,5 мм на поверхности глаза. По оси ординат справа и слева - частота импульсов в секунду
Благодаря механизму латерального торможения, как видно на рис. 50, усиливается различие в возбуждении по-разному освещенных соседних омматидиев. Единицы сильнее освещенной левой половины глаза, примыкающие к зоне слабого освещения (левая половина кривой 2), испытывают со стороны этой зоны меньшее торможение, поэтому реагируют более частыми импульсами, чем те омматидии, которые окружены зоной одинакового возбуждения. И наоборот, омматидии правой половины глаза, примыкающие к зоне более сильного засвета (правая половина кривой 2), тормозятся ею сильнее, чем друг другом, и поэтому они реагируют более редкими импульсами. В результате подчеркивается краевой контраст на границе перепада освещенности на сетчатке, что в конечном счете способствует выявлению контура или границы по-разному освещенных участков. Расчеты показали также, что механизм латерального торможения, устраняя последствия перекрывания полей зрения смежных омматидиев, улучшает разрешающую способность глаза. Идея латерального взаимодействия между соседними рецепторами положена в основу некоторых технических декодирующих устройств, предназначенных, например, для расшифровки нечетких фотоснимков.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'