1. Современное представление о функциональной организации фасеточного глаза
Фасеточные, или сложные, глаза имеют почти все взрослые насекомые подкласса Pterygota (кроме блох и некоторых других групп) и их личинки у форм с неполным превращением. В подклассе бескрылых насекомых только у Machilidae встречаются глаза, близкие по своей организации к фасеточным. У остальных представителей насекомых место фасеточных глаз занимают простые латеральные глазки.
Строение омматидиев
Омматидии-структурные единицы фасеточного глаза - имеют свой собственный светопреломляющий (диоптрический), светоизолирующий (экранирующий) и светочувствительный (фоторецепторный) аппарат. У разных насекомых число омматидиев различно. Чаще всего оно достигает нескольких тысяч, вплоть до 28 тыс. у некоторых стрекоз и бабочек.
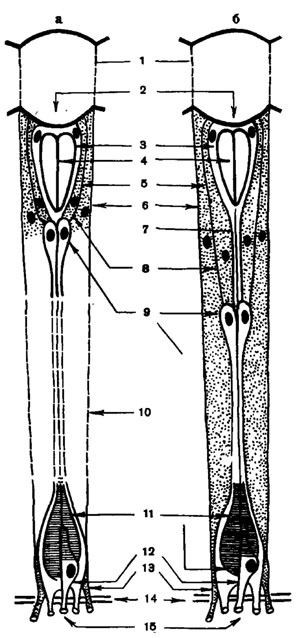
Рис. 42. Схема омматидия оптикосупер-позиционного глаза в состоянии световой (б) и темиовой (а) адаптации (по Ноrridge, 1972; Meyer-Rochow, 1972, видоизменено) : 1 - роговичная линза; 2 - роговичный конус; 3 - кристаллические клетки; 4 - кристаллический конус; 5 - гранулы экранирующего пигмента; 6 - придаточные пигментные клетки; 7 - кристаллический тракт; 8 - ирисовые пигментные клетки; 9 - тела ретинулярных клеток; 10 - прозрачная зона; 11 - рабдом; 12 - базальная зрительная клетка; 13 - трахейный тапетум; 14 - базальная мембрана; 15 - аксоны зрительных клеток
Светопреломляющий аппарат омматидиев образован прозрачными роговичнои линзой и кристаллическим конусом (рис. 42, 1, 2). С внешней стороны роговица омматидия имеет вид более или менее выпуклой шестигранной фасетки. При электронно-микроскопических исследованиях было обнаружено, что фасетки дневных и ночных бабочек, двукрылых, ручейников покрыты сеткой из прозрачных кутикулярных бугорков высотой около 200 нм и на столько же отстоящих друг от друга. На модели глаза, а также путем соответствующих расчетов показано, что наружная сетка из бугорков снижает отражение света и тем повышает (на 5%) прозрачность роговицы. Подобно просветляющему, или антирефлексному, покрытию у современной оптики, сетка облегчает проникновение лучей с λ=320-700 нм из воздуха в более плотную среду глаза. Ее биологическая роль дополняется также камуфлированием глаза за счет устранения блеска его поверхности.
Роговичная линза как собирательный светопреломляющий аппарат функционирует совместно с непосредственно примыкающим к ней кристаллическим конусом. Последний представляет собой прозрачное плотное или жидкое тело, образованное четырьмя Земперовыми (кристаллическими) клетками - их телами или экстраклеточным выделением (рис. 42, 3). Сравнительно недавно австралийский исследователь Г. А. Хорридж обнаружил, что кристаллические клетки продолжаются в виде тонких кристаллических нитей (тракта) (рис. 42, 7) в глубь омматидия. Это особенно заметно в оптикосуперпозиционных глазах, где им приписывают роль световода.
Светоизолирующий аппарат омматидия представлен набором пигментных клеток, содержащих гранулы экранирующего пигмента. Этот аппарат необходим для защиты рабдома от бокового засвета и повышения направленной чувствительности омматидия. В дистальной части омматидия всегда присутствуют две корне-агенные клетки и не менее четырех ирисовых пигментных клеток (рис. 42, 8), а в аппозиционных глазах имеется еще несколько ретинальных пигментных клеток, лежащих в проксимальной половине омматидия. Кроме того, гранулы защитного пигмента присутствуют в теле зрительных клеток и в межклеточной среде. Различают оммохромы - более темные (красные, коричневые) пигменты и птерины - более светлые. Первые поглощают лучи в широком диапазоне спектра, кроме красных лучей, вторые - главным образом ультрафиолетовые (УФ) лучи. Иногда защитные пигменты частично или полностью отсутствуют. Такие светлоглазые мутанты известны, например, среди мух, и их широко используют для исследования различных зрительных функций. В глазах опти-косуперпозиционного типа гранулы защитного пигмента периодически переходят из дистального положения в проксимальное и наоборот в зависимости от интенсивности освещения, времени суток и других причин (рис. 42, а, б).
Светочувствительный аппарат омматидия образуют чаще всего 8-9 зрительных, или ретинулярных, клеток с четко выраженными аксонами, связывающими их с оптическим центром. Зрительные клетки (рис. 42, 9) лежат обычно в 2-3 яруса, а одна или две из них, так называемые базальные клетки (12), асимметрично смещены к базальной мембране, подстилающей дно глаза. Аксон базальной клетки идет непосредственно во второй оптический ганглий, а не оканчивается в первом, как у остальных зрительных клеток. Разные зрительные клетки омматидия отличаются различной спектральной и поляризационной чувствительностью.
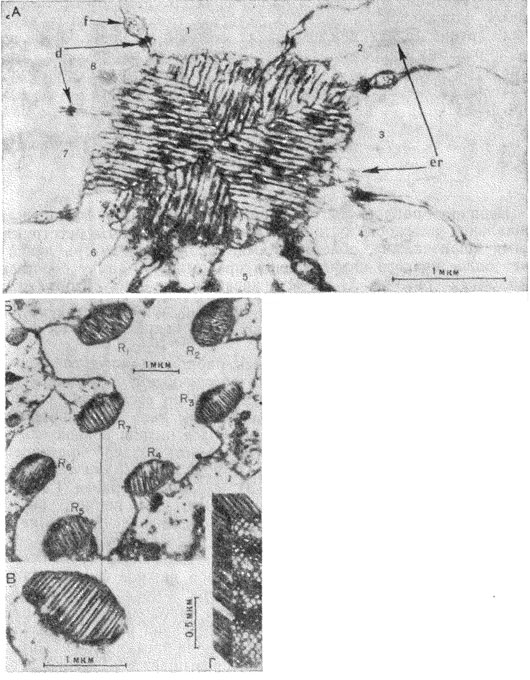
Рис. 43. Электронные микрофотографии поперечного среза раб-дома. А - медоносная пчела Apis mellifera (пo Goldsmith, 1962); 1-8 - зрительные клетки; d - скопление десмосом; еr - эндоплазматический ретикулум; f - нити кристаллических клеток; Б, В, Г - муха Drosophila melanogaster (пo Wolken, 1961): R1-R7 - семь раб-домеров; Г-реконструкция участка рабдомера, рассеченного вдоль
Вдоль оптической оси омматидия (иногда ярусами) расположены рабдомеры зрительных клеток, образующие в совокупности рабдом (рис.42, 11). Различают замкнутый (спаянный) рабдом (рис. 43, А) - когда отдельные рабдомеры тесно объединены друг с другом, и открытый (рис. 43, Б) - если отдельные рабдомеры (кроме двух) разобщены и оптически изолированы друг от друга. Каждый рабдомер сложен большим количеством микроскопических трубочек - микровилл - диаметром около 500 Å, лежащих перпендикулярно оптической оси омматидия (рис. 43, В, Г). Микровиллы, образованные двойной мембраной толщиной около 50 Å, содержат зрительный пигмент, концентрация которого достигает 107 молекул на рабдомер. Поэтому рабдомер является собственно фоторецепторным элементом зрительной клетки, функциональным аналогом наружного членика палочек и колбочек сетчатки позвоночных.
Замечательная особенность структуры рабдома заключается в упорядоченном расположении его микровилл. Например, у медоносной пчелы микровиллы восьми наиболее крупных клеток расположены в двух взаимно перпендикулярных направлениях, при этом ориентация трубочек в рабдомерах соседних омматидиев остается одинаковой на больших участках глаза. У других насекомых картина ориентации микровилл может быть иной, но они всегда по-разному направлены в несимметричных рабдомерах. Такая структура рабдома при дихроизме поглощения света молекулами зрительного пигмента обеспечивает анализ поляризации лучей (см. ниже).
Проекция внешнего мира на растр зрительных клеток
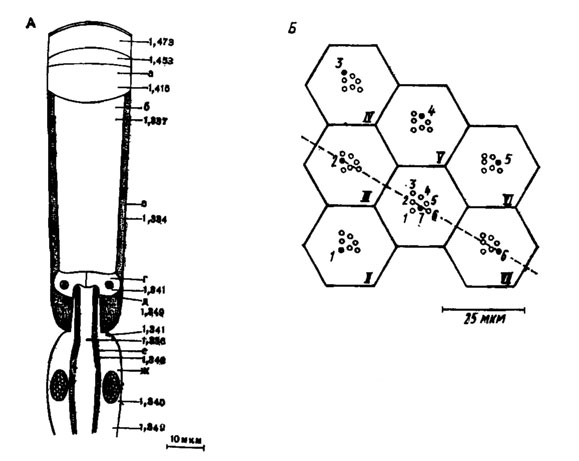
Рис. 44. Схема аппозиционного глаза мухи Calliphora erythrocephala (пo Seitz,1968): А - клеточные элементы омматидия и их показатели A - клеточные элементы преломления: а - роговица, б - кристаллический конус, в - пигментная клетка, г - Земперова клетка, д - заключенный в 'шапочку' вырост рабдомера, е - рабдомер, ж - зрительная клетка; Б - группа из семи (I-VII) омматидиев. Черным кружком изображен один из семи (1-7) рабдомеров каждого омматидня, смотрящий в одну и ту же точку пространства омматидия и их показатели преломления: а - роговица, б - кристаллический конус, в - пигментная клетка, г - Земперова клетка, д - заключенный в 'шапочку' вырост рабдомера, е - рабдомер, ж - зрительная клетка; Б - группа из семи (I-VII) омматидиев. Черным кружком изображен один из семи (1-7) рабдомеров каждого омматидия, смотрящий в одну и ту же точку пространства
Вершины рабдомеров, по крайней мере в глазу мух, лежат в фокальной плоскости оптики омматидия. Они одеты бесструктурными "шапочками", облегчающими вхождение внутрь лучей за счет более низкого коэффициента преломления, чем у собственно рабдомера (рис. 44, А, д). Что касается качества изображения, создаваемого оптикой омматидия, то оно малосущественно для зрения, поскольку его детали не могут быть разрешены ретинулярными клетками. Объяснение этому следует искать в особенностях проекции сетчатки в зрительном ганглии, при которой сигналы разных зрительных клеток теряют свой "адрес" уже на первом интернейроне.
Отдельные рабдомеры или весь рабдом замкнутого типа обладают свойствами волновода, что обеспечивает проведение лучей и их поглощение зрительным пигментом по всей длине рабдома. Дело в том, что к рабдомеру со стороны тела зрительной клетки примыкают структуры, имеющие более низкий показатель преломления. Поэтому часть лучей на границе раздела претерпевает полное внутреннее отражение. Так, у мухи Calliphora erythrocephala показатель преломления (п) рабдомеров равен 1,349, а у окружающих его структур - 1,344-1,340; угол полного внутреннего отражения для такого рабдомера-волновода составляет 83° (рис. 44, А).
Поле зрения отдельного рабдомера Calliphora erythrocephala в среднем около 7°. Но имеющая особо важное функциональное значение полуширина поля зрения - угол, в пределах которого интенсивность истечения света снижается наполовину от максимального, - отдельного рабдомера этой мухи равна всего 1,2°. Судя по электрофизиологическим измерениям, поле зрения замкнутого рабдома аппозиционных глаз тоже не превышает 8°. Однако в опти-косуперпозиционных глазах, например жука-плавунца, оно существенно шире - практически охватывает поле зрения целого глаза. Поэтому поля зрения смежных омматидиев (а их оси расходятся под углом обычно от 1 до 3-4о) даже в аппозиционных глазах заметно перекрываются. Впрочем, это обстоятельство, как полагают (см. ниже), благодаря механизму латерального торможения не ухудшает остроту зрения насекомого.
Со времени классических исследований фасеточного глаза членистоногих венским офтальмологом А. Экснером (1891) единодушно принималось, что диоптрический аппарат омматидия представляет собой преломляющий цилиндр - такую линзу, которая сложена слоями с показателем преломления, возрастающим к центру. Теперь всеобщность этого утверждения подвергли сомнению. Только у нескольких видов насекомых, в частности у светляка Phausis splendidula, показатель преломления действительно возрастает к центру кристаллического конуса, а у остальных насекомых диоптрический аппарат аналогичен обычной собирательной линзе (рис. 44, А). В аппозиционных и оптикосуперпозиционных глазах эта линза рисует в каждом омматидии обособленное прямое или перевернутое (у разных насекомых по-разному) изображение приблизительно в плоскости шапочек рабдомеров. Мы уже знаем, что качество этого изображения, иногда (у мух, например) довольно высокое, не определяет степени детализации воспринимаемой картины. Видеть отдельные детали изображения препятствует своеобразие проекции сетчатки в зрительном ганглии, а у насекомых с единым замкнутым рабдомом к тому же все зрительные клетки, независимо от геометрии изображения стимула на рабдоме, получают равное количество света. В результате внешний мир анализируется насекомыми с точностью до растра омматидиев, а не отдельных зрительных клеток, как у позвоночных. В этом смысле сохраняется прежнее - мюллеровское - представление о мозаичности зрения насекомых.
Неожиданной оказалась проекция внешнего мира на растр зрительных клеток у мух. Углы (2-4°) между оптическими осями семи рабдомеров каждого омматидия в глазу мухи находятся в таком соответствии с углами разобщения соседних омматидиев, что в семи смежных омматидиях имеется по одному рабдомеру, смотрящему в одну и ту же точку пространства (рис. 44, Б). Иначе говоря, семь рабдомеров одного омматидия смотрят в разные точки пространства, но те же точки видят по одному рабдомеру в шести соседних омматидиях. Аксоны шести зрительных клеток, рабдомеры которых направлены в одну и ту же точку, сходятся в одном и том же оптическом патроне ламины, где их сигналы суммируются. Поэтому между числом светящихся точек на сетчатке мухи и числом оптических патронов существует соотношение 1:1, т. е. число оптических патронов равно числу омматидиев. При такой нейрооптической конструкции за счет увеличения эффективности входного зрачка из многих фасеток светосила глаза повышается в 7 раз по сравнению с классическим аппозиционным глазом, как, например, у медоносной пчелы. Поэтому глаза мух теперь выделяют в особый тип нейросуперпозиционных глаз.
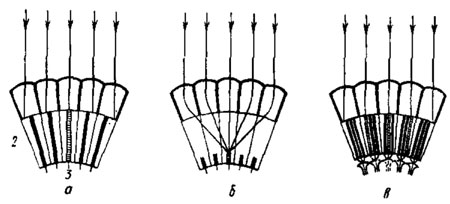
Рис. 45. Упрощенная схема возникновения сетчаточного изображения в аппозиционном (а), оптико- (б) и нейросуперпозиционном (в) глазу (по Kirschenfeld, 1970). 1 - диоптрический аппарат омматидия; 2 - зрительные клетки с замкнутым (а и б) или открытым (в) рабдомом; 3 - проекция аксонов зрительных клеток на оптические патроны ламины. Заштрихованы те рабдомеры, на которые попадают параллельно идущие лучи света (показаны стрелками)
Соответственно сказанному мы будем различать у насекомых три морфологических типа фасеточных глаза: аппозиционный, оптикосуперпозиционный и нейросуперпозиционный. Аппозиционные глаза свойственны большинству дневных: насекомых, кроме двукрылых и клопов. Глаза этого типа приспособлены для работы при избытке света. Анатомически для них характерны удлиненный рабдом, достигающий вершины кристаллического конуса, и постоянная пигментная изоляция смежных омматидиев (рис. 44, А; 45, а). Оптикосуперпозиционные глаза встречаются у подавляющего большинства "очных насекомых. Здесь рабдом укорочен, между ним и кристаллическим конусом имеется отчетливо выраженный кристаллический тракт (см. рис. 42, а). Защитный пигмент в удлиненных ирисовых пигментных клетках мигрирует в зависимости от состояния адаптации глаза. При пониженном освещении, в том числе вечером и мочью, гранулы пигмента сосредоточиваются в дистальном положении, в результате исчезает экранизация пространства между кристаллическими конусами и вершинами рабдомеров, и в один и тот же омматидий могут попадать лучи, прошедшие через группу смежных фасеток (рис. 45, б). Еще не ясно до конца, как работают оптикосуперпозиционные глаза. Но у некоторых ночных жуков и бабочек, а также у толстоголовок Hesperiidae при дистальном положении ирисового защитного пигмента безусловно имеет место суперпозиция лучей на рабдомах, чем достигается повышенная чувствительность к свету, хотя и ценой потери качества изображения. При дневном положении защитного пигмента эти глаза функционируют, по-видимому, аналогично аппозиционным; ретинальное изображение образуется за счет аппозиции лучей, прошедших через разные фасетки, т. е. без наложения друг на друга. Нейросуперпозиционные глаза известны только у двукрылых - насекомых с открытым рабдомом. В них нет суперпозиции лучей на сетчатке, но имеет место суперпозиция сигналов, исходящих от зрительных клеток разных омматидиев, в первом оптическом ганглии (рис. 45, в). Глаза этого типа, как и аппозиционные, обеспечивают высокое разрешение точек пространства, но отличаются от вторых повышенной светосилой. Недаром мухи активны при такой низкой освещенности, при которой не летают другие дневные насекомые.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'