Глава V. Терморецепция и гигрорецепция
1. Температурные рецепторы
Локализация и строение сенсилл. Восприятие температуры и способность ориентироваться по температурному градиенту -неотъемлемое свойство всех насекомых. Температура окружающей среды - важный экологический фактор, обеспечивающий нормальную жизнедеятельность организма насекомого. Для насекомых характерна реакция термопреферендума - способность собираться в зоне оптимальной температуры. Тепло становится одним из стимулов, по которому кровососущие насекомые - комары и клопы - осуществляют ориентацию на хозяина-прокормителя. Многие стволовые вредители, например жуки-усачи, отличают ослабленные деревья от здоровых по разнице температур их поверхности.
Чувствительными органами насекомых, реагирующими на изменение температуры, служат трихоидные, базиконические и цело-конические сенсиллы, расположенные главным образом на антеннах. Так, у таракана Periplaneta americana крупные волоски, чувствительные к изменению температуры, составляют 0,1% от всех сенсилл на антенне. Температурные рецепторы в целоконических сенсиллах найдены на антеннах саранчи, пчелы, различных видов комаров рода Aedes, личинок кожеедов Dermestes siblricus. Многие насекомые и их личинки имеют температурные рецепторы в базиконических сенсиллах на максиллярных и лабиальных пальпах. Это - гусеницы коконопрядов и сатурний, почвообитающие личинки жуков (табл. 10). Удаление антенн и пальп в эксперименте не снимает температурную чувствительность у сверчков и тараканов. Предполагают, что терморецепторы у этих насекомых могут быть также и на лапках ног, на брюшке, церках и т. д.

Таблица 10 Распределение обонятельных, вкусовых и температурных рецепторов на придатках головы личинок жуков (по Елизарову и др., 1968; Елизарову, Куэн, 1971)
Ультраструктура сенсилл, чувствительных к изменению температуры окружающего воздуха, до настоящего времени не изучена.
Известные температурные рецепторы насекомых реагируют на конвекционное тепло - энергию молекул нагретого воздуха. Предположение об ориентации насекомых по инфракрасному излучению нагретых тел пока не нашло экспериментального подтверждения. Вместе с тем высокая чувствительность к незначительным изменениям температуры позволяет говорить о способности насекомых чувствовать температурный градиент при удалении от источника тепла. Так, чувствительные к понижению температуры клетки в трихоидных антеннальных сенсиллах Periplaneta americana реагируют пятикратным увеличением частоты импульсов по сравнению с фоном при понижении температуры окружающего воздуха всего на 1о. Аналогичные рецепторы гусениц Dendrolimus pini и Philosamia cynthia увеличивают частоту импульсов на 150-300% при изменении температуры на 1о в течение 0,5 с. Сходная картина наблюдается в температурных рецепторах пчел и комаров. Температурный градиент над поверхностью кожи человека изменяется от 32 до 34° на расстоянии более 40 см. Конвекционные потоки теплого воздуха, идущие от человека, могут иметь локальные температурные различия на 1-2°, что используется комарами при ориентации на небольших расстояниях.
Свойства холодовых и тепловых рецепторов. Температурные рецепторы насекомых можно разделить на два типа: одни из них увеличивают частоту импульсов при снижении температуры, другие - при увеличении температуры. Холодовые рецепторы были впервые обнаружены на антенне пчелы Apis mellifera. Введение вольфрамового электрода в целоконическую сенсиллу позволило зарегистрировать фазово-тонический ответ чувствующей клетки на резкое понижение температуры. Чувствительность изменения частоты импульсов в тонической части ответа была равна 2 имп/с/град. Холодовой рецептор с фазово-тоническим ответом был обнаружен также в целоконической сенсилле антенны саранчи. Импульсный ответ холодового рецептора трихоидной сенсиллы антенны таракана Periplaneta americana имел хорошо выраженный фазовый характер. Уменьшение температуры на 0,5о вызывало увеличение фоновой активности с 25 до 50 имп/с. В любом случае высокая частота импульсов поддерживалась лишь в течение 1-2 с после начала раздражения, несмотря на то, что раздражитель продолжал действовать. В результате быстрой адаптации частота импульсов приближалась к величине фона и удерживалась на одном уровне в течение 100-200 с действия раздражителя. Прекращение стимуляции холодным воздухом вызывало кратковременное (1-2 с) торможение импульсной активности рецептора, после чего фоновая активность снова восстанавливалась.
Среди многих (около 18) чувствующих клеток, иннервирующих сенсиллы максиллярнои пальпы гусеницы соснового коконопряда Dendrolimus pini, можно выделить одну клетку, дающую залп импульсов при приближении к сенсиллам холодного предмета. Нагретый предмет вызывал торможение фоновой активности.
Тепловые рецепторы изучены в меньшей степени, чем холодовые. Установлено, что поднесение теплой стеклянной палочки к базиконическим сенсиллам антенн и максиллярных пальп личинки жука Anisoplia austriaca и к целоконическим сенсиллам антенн личинки кожееда Dermestes sibiricus вызывало залп импульсов при наличии слабой фоновой активности. Тепловые рецепторы дистальных члеников антенн клопа Oncopeltus fasciatus имеют хорошо выраженный фазовый ответ, длящийся после небольшого латентного периода около 2-3 с, причем фоновая активность отсутствует до и после залпа, несмотря на продолжающееся действие струи теплого воздуха. Максимальную чувствительность к повышению температуры рецептор имеет в диапазоне 28-30°. Сенсиллы самцов и самок клопов показывают одинаковую реакцию на температурный стимул.
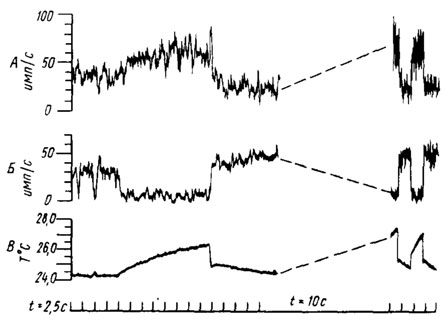
Рис. 39. Интегрированные импульсные ответы целоконической сенсиллы антенны комара Aedes aegypti, указывающие на реакцию двух клеток с разной амплитудой потенциала при повышении или понижении температуры струи воздуха (по Davis, Socolove, 1974). А, Б - тепловой и холодовый рецепторы соответственно; по оси абсцисс - время, с; по оси ординат - частота импульсов, имп/с. В - температура воздуха
Целоконическая сенсилла на кончике антенны комара Aedes aegypti имеет две чувствующие клетки, причем одна является Холодовым, а другая - тепловым рецептором. Их свойства идентичны описанным выше у других насекомых. Возбуждению теплового рецептора соответствует торможение холодового и наоборот (рис. 39). Ответ рецепторов фазово-тонический. Быстрое возрастание частоты импульсов над величиной фоновой активности в начале залпа проявляется при изменении температуры на 0,05° и составляет 4 имп/с для холодового рецептора и 3,2 имп/с для теплового. Частота импульсации в тонической части залпа теплового рецептора увеличивается при достижении температуры около 24° и достигает максимума при 28,5° (35 имп/с), а в случае холодового рецептора имеет пик при 26° (30 имп/с). Чувствительность тонической части залпа теплового рецептора равна 5-7 имп/с/град, а холодового - 5-6 имп/с/град. Действие на целоконическую сенсиллу инфракрасного излучения с длиной волны 10 мкм, двуокиси углерода, влажного воздуха, запахов аттрактантов и репеллентов не вызывало изменений фоновой активности.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'