3. Морфологическая характеристика обонятельных сенсилл
Локализация. У большинства насекомых обонятельные сенсиллы расположены на антеннах. Экспериментально это впервые было показано в 1921 г. Е. Фишером, который наблюдал постепенное уменьшение интенсивности ольфакторной реакции вплоть до ее полного исчезновения в результате последовательной ампутации члеников антенны пчелы. Однако у некоторых мух и комаров обонятельные сенсиллы могут встречаться также и на околоротовых придатках. Регистрация биопотенциалов от антенналыного нерва и нервной цепочки сверчка показала, что обонятельные сенсиллы у этого насекомого находятся на околоротовых придатках вместе с контактными хеморецепторами.
Рассматривая распределение ольфакторных сенсилл на антеннах насекомых, можно увидеть различную степень их концентрации на поверхности кутикулы. Диффузное распределение сенсилл наблюдается у тараканов, некоторых прямокрылых и перепончатокрылых насекомых, в частности у ос, пчел и наездников.
Для комаров характерна группировка сенсилл на различных участках флагеллума антенны. Обонятельную функцию у этих насекомых выполняют тонкостенные волоски, разделяемые на три типа по своей величине. Один тип сенсилл доминирует на дистальных члениках флагеллума антенны комаров. Сенсиллы другого типа сконцентрированы на проксимальных члениках, а третьего типа равномерно распределены по флагеллуму антенны.
Следующую ступень концентрации обонятельных сенсилл можно встретить у дневных бабочек. На большей части члеников антенны, начиная с дистального конца флагеллума, базиковические тонкостенные сенсиллы собраны в тесные группы, локализованные на медиальной стороне члеников и ограниченные от других участков кутикулы. Скопление обонятельных сенсилл, образующих рецепторное поле, окружено гребневидными утолщениями кутикулы. Такой тип обонятельного органа свойствен антеннам бабочек из семейств нймфалид и бархатниц. У бабочек из семейства белянок чувствительные поля имеют меньшие размеры и углублены по отношению к поверхности кутикулы членика антенны.
Дальнейшее развитие обонятельных органов наблюдается у двукрылых насекомых из подотряда короткоусых мух. Здесь на третьем членике антенны встречаются так называемые обонятельные ямки, представляющие собой глубокие впячивания кутикулы членика, открывающиеся наружу небольшими отверстиями. Дно этих впячиваний покрыто базиконическими сенсиллами, воспринимающими запахи. В отдельных случаях, связанных с олигомеризацией антенн, например, у личинок комаров и некоторых жуков. Обонятельные сенсиллы могут сливаться, образуя массивные органы с единым кутикулярным отделом. Наиболее ярко такие органы выражены у цикад, где второй членик антенны плотно покрыт розетками уплощенных пальцеобразных выростов, выходящих из углубления кутикулы и представляющих собой результат слияния многих обонятельных сенсилл.
Таким образом, на примере наездников, комаров, дневных чешуекрылых, мух и цикад мы видим процесс постепенного усложнения обонятельного органа насекомых - от равномерно распределенных по антенне сенсилл до специализированных многоклеточных образований.
Самым характерным структурным признаком обонятельных сенсилл насекомых следует считать многочисленные поры их кутикулярного отдела. Впервые поры были обнаружены в базиконических сенсиллах кузнечика Romalea microptera, когда было показано, что кутикула базиконических сенсилл проницаема для водных растворов кристаллического фиолетового и что краситель проникает через поры, имеющиеся в кутикулярном отделе. В последние 10 лет поры широко исследовались с помощью электронного микроскопа.
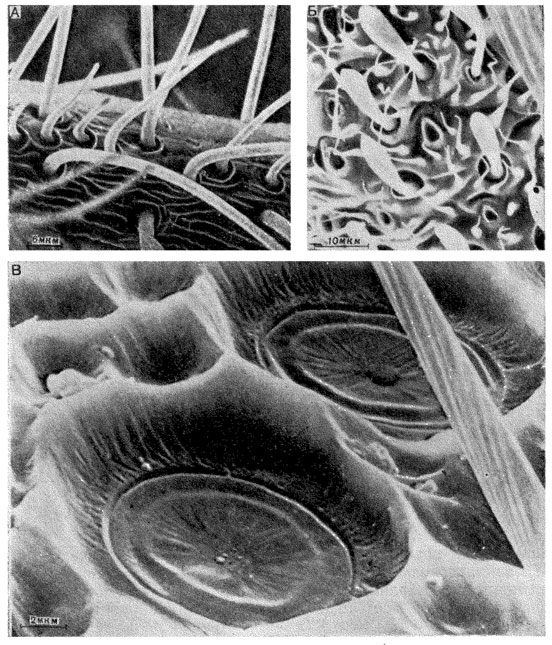
Puc. 26. Кутикулярные отделы обонятельных сенсилл. А - трихоидные обонятельные сенсиллы на поверхности флагеллума антенны бабочки волнянки Orgyia prisca; Б - булавовидные сенсиллы максиллярных пальп комара Culex pipiens; В - плакоидные сенсиллы на поверхности флагеллума антенны самца пчелы Еnсеrа стеrеа
Ультраструктура обонятельных сенсилл. Кутикулярный отдел плакоидной сенсиллы имеет вид тонкой кутикулярной пластинки овальной или продолговатой формы (рис. 26,(В). В настоящее время плакоидныё сенсиллы обнаружены на антеннах жуков, равнокрылых хоботных и перепончатокрылых. Плакоидныё сенсиллы пчелы были первыми хеморецепторными сенсиллами насекомых, исследованными с помощью электронного микроскопа. Очень тонкая сочленовная мембрана сенсиллы пересечена 120-150 радиально идущими утолщениями, между которыми расположены поры. Дальнейшие электронно-микроскопические исследования показали, что поверхность пластинки и сочленовной мембраны представляет собой слой эпикутикулы, под которым расположены зубцы, состоящие из менее электронноплотной кутикулы.
В промежутках между зубцами обнаруживаются поры, диаметр которых составляет около 100 Å. В состав плакоидной сенсиллы пчелы входят 12-18 чувствующих клеток, периферические отростки которых заканчиваются под сочленовной мембраной, т. е. в области локализации пор.
Кроме сочленовной мембраны поры могут располагаться также по всей поверхности кутикулярной пластинки плакоидной сенсиллы. У осы Nasonia vitripennis вся пластинка перфорирована небольшими неправильными отверстиями, через которые проходит раствор кристаллического фиолетового при нанесении его на наружную поверхность сенсиллы.
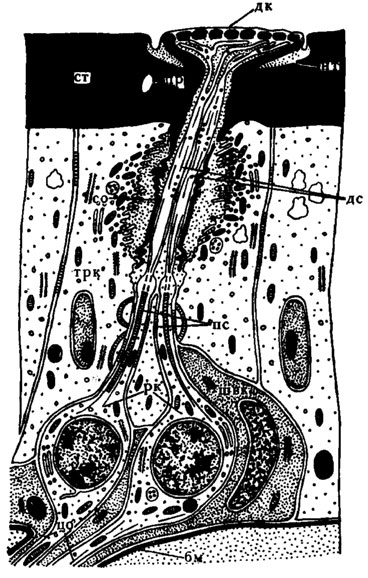
Рис. 27. Схема строения плакоидной сенсиллы жука Acilius sulcatus (по Иванову, 1969): бм - базальная мембрана; дк - диск; дс - дистальиый сегмент; ит - наружное тельце; пс - проксимальный сегмент периферического отростка; пр - проток железы; ст - стенка наружного скелета антенны; трк - трихогенная клетка; швк - Шванновская клетка. Остальные обозначения, как на рис. 22
Иное строение кутикулярного отдела плакоидных сенсилл у жука Acilias silvaticus (рис. 27). Здесь полость кутикулярного диска, погруженного в чашеобразное углубление кутикулы, связана с каналом, пронизывающим кутикулу антенны. Через этот канал проходят два периферических отростка чувствующих клеток, разветвляющихся в полости диска. Верхняя пластинка снабжена лорами, диаметр которых увеличивается в направлении наружной поверхности.
Уникальная плакоидная сенсилла описана у наездников Coleoides brunneri. В ней находятся две продолговатые куполообразные пластинки, подвешенные изнутри к кутикулярному отделу сенсиллы. Этими пластинками внутренняя полость сенсиллы разделяется на три канала. Ветви периферических отростков заполняют две трети срединного канала, а два боковых канала и треть срединного заняты тормогенной клеткой. Авторы приписывают этим сенсиллам восприятие инфракрасного излучения. К разновидности плакоидных сенсилл могут быть отнесены также сложные чувствительные органы - ринарии, локализованные на антеннах тлей.
Тринхоидные обонятельные сенсиллы с тонкими стенками представляют собой один из наиболее распространенных типов обонятельных сенсилл насекомых (см. рис. 26,А). Они характеризуются небольшой толщиной кутикулы и наличием пор. Число пор на одном волоске может достигать 1500-2500. Показано, что поры трихоидных сенсилл шелкопряда связаны с периферическими отростками чувствующих клеток поровыми трубками.
Трихоидные обонятельные сенсиллы имеют различное число чувствующих клеток. Так, в сенсиллах короеда Scolytis multistriatus обнаружены 2 клетки, в сенсиллах муравья Lasius fuliginosus - 20-30, а в сенсиллах клопа Lygaeus kalmii - 50-60 клеток.
Периферические отростки чувствующих клеток обонятельных сенсилл обычно начинают ветвиться в основании волоска. Однако в некоторых трихоидных сенсиллах, например на антеннах шелкопряда Bombyx mori, периферические отростки не ветвятся.
Своеобразно устроены трихоидные сенсиллы ногохвостки Onychiurus sp. Поры и поровые трубки кутикулярного отдела подстилаются аморфным электронноплотным материалом, секретируемым оберточными клетками, служащим, по всей вероятности, стимулопроводящим элементом сенсиллы.
Разновидностью трихоидных сенсилл считаются сенсиллы антенн эмбий Ptilocerembia sp. и тараканов Arenivaga sp. Это толстостенные волоски, кутикула которых имеет поры. Такого рода сенсиллы сочетают в себе признаки толстостенных и тонкостенных хеморецепторов.
Тонкостенные базиконические сенсиллы широко распространены на антеннах насекомых и, согласно электрофизиологическим данным, могут функционировать как типичные обонятельные рецепторы.
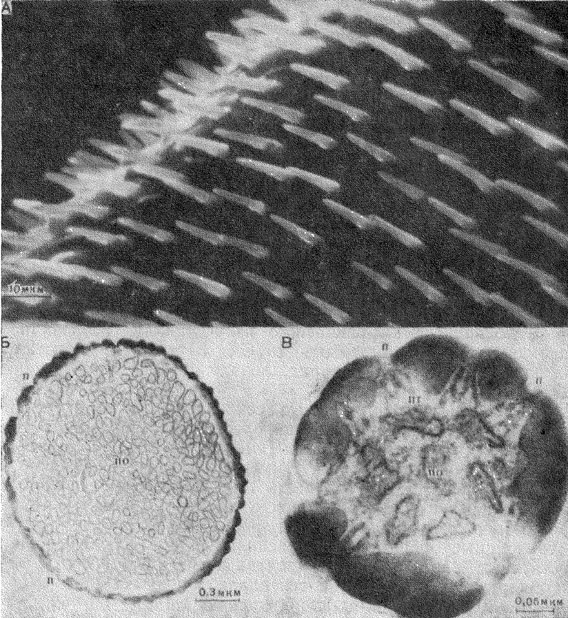
Рис. 28. Строение обонятельных сенсилл. А - базиконические сенсиллы на антенне мухи Protophormia terraenovae; Б - поперечный срез кутикулярного отдела базиконической сенсиллы москита Phlebotomus papatasi; В - поперечный срез антеннальной трихоидной сенсиллы бабочки павлиноглазки Actias artemis. п - пора; пт - поровые трубочки; по - периферические отростки
Наружный кутикулярный отдел базиконической сенсиллы имеет вид небольшого шипа, стенки которого перфорированы многочисленными порами (рис. 28,А,Б). Базиконические сенсиллы, сохраняя общий план строения, различаются у разных видов насекомых по числу чувствующих клеток, величине кутикулярного отдела, а также по числу и размеру пор.
Кутикулярный отдел базиконических сенсилл обычно имеет толщину стенок 0,05-0,3 мкм с гексагонально размещенными порами. Такое размещение пор наиболее экономично с точки зрения эффективного использования площади. Кутикулярный отдел базиконических сенсилл мухи Phormia regina имеет 180-360 пор, а в базиконических сенсиллах жука Necrophorus sp. число пор достигает 13000-17000. Наружное отверстие поры не превышает 50 Å.
В полости кутикулярного отдела базиконических сенсилл периферические отростки рецепторных клеток интенсивно ветвятся. В 1967 г. в первых электронномикроокопических работах Е. Слайфер установила, что к порам подходят тонкие ветви, диаметр которых равен 200 Å. Позже было показано, что в порах находятся не кончики периферических отростков, а пучки тонких нитей, которые образуются путем выпячивания стенки периферического отростка. Эти тончайшие нити диаметром 200 Å были названы "поровыми филаментами", или поровыми нитями. Им приписывается функция первичного взаимодействия с молекулами пахучих веществ. Однако в 1969 г. немецким морфологом К.нстом было показано, что в базиконических сенсиллах жука Necrophorus трубочки, сходные с поровыми нитями, не являются выростами мембран периферических отростков, а имеют экстраклеточное происхождение. В период онтогенеза сенсилл трубочки, связанные с порами, встречаются в просвете шипа еще до вхождения периферических отростков в полость последнего. В пользу экстраклеточной природы трубок говорит также их небольшой диаметр, равный 150-200 Å, в то время как диаметр тончайших ветвей периферических отростков - около 500 Å. Кроме того, толщина стенки трубочек составляет около 30 Å, что не соответствует толщине клеточной мембраны. Таким образом, поры и поровые трубки образуют единую по происхождению поро-трубчатую систему, которая формируется в процессе онтогенеза оберточными клетками и, видимо, служит для проведения молекул пахучих веществ к периферическим отросткам.
На максиллярных пальпах комаров и москитов встречаются булавовидные органы, имеющие тонкую кутикулярную стенку (см. рис. 26,Б). Изучение поведения комаров и электрофизиологические исследования показали, что булавовидные органы имеют чувствующие клетки, воспринимающие углекислоту, и клетки с ольфакторной функцией.
Длина булавовидных органов составляет 16-17 мкм, а их диаметр - 5-6 мкм. Они иннервируются тремя биполярными нейронами. Толщина кутикулы булавовидного органа равна в среднем 0,1 мкм. Через весь слой кутикулы булавовидных органов проходят поровые каналы. С каждым поровым каналом связаны 2-3 поровые трубки диаметром 100-140 Å и толщиной стенок 30 Å. Поровые трубки представляют собой кутикулярные образования, и их сердцевина электроннопрозрачна. Начинаясь от проксимальной части порового канала, они простираются в полость волоска на расстояние до 0,1 мкм. Иногда периферические отростки образуют спиралеобразную пластинчатую структуру, состоящую из складок их мембран.
Целоконическая сенсилла представляет собой конусообразный или волосковидный орган, погруженный в кутикулу. Целоконические сенсиллы широко распространены среди насекомых и встречаются у представителей многих отрядов.
В связи с тем, что число целоконичеоких сенсилл на единицу площади невелико, исследование их ультраструктуры затруднено, и в настоящее время нет полных сведений о строении этих сенсилл.
По данным световой и электронной микроскопии, в целоконических сенсиллах прямокрылых имеется несколько чувствующих клеток, периферические отростки которых подходят к вершине конуса. Периферические отростки не ветвятся и заключены в сколопоидную оболочку. Вершина конуса имеет отверстие, которое обнаруживается при нанесении растворов красителей. Сходное строение имеют целоконические сенсиллы антенн уховерток.
В отличие от вышеописанного типа стенки целоконических сенсилл бабочки Danaus gillipus перфорированы многими порами, однако поровые трубки отсутствуют.
У некоторых видов стрекоз Lygoptera и Anisoptera имеются как простые, так и сложные целоконические сенсиллы. Кутикулярный конус сенсилл пронизан порами. Сложные сенсиллы представляют собой несколько кутикулярных отделов, отличающихся по внешнему виду, которые находятся в углублении кутикулы.
Целоконические сенсиллы присутствуют на антеннах слепней в разном количестве в зависимости от вида насекомого и обнаруживают поразительную общность ультраморфологии. Наибольшее число целоконических сенсилл (300-400) насчитывается у слепней родов Tabanus и Hybomitra, среднее (150-180) - у видов рода Haematopota и наименьшее (40-120) - у видов рода Chrysops.
Сложные органы обоняния. Кроме сенсилл с четко выраженной ольфакторной функцией в последнее время в литературе появились сообщения о новых типах чувствительных органов, ультраструктурная организация которых свидетельствует об их обонятельной функции.
Опоясывающие нити. Ультраструктура тонкостенных кутикулярных нитей, окружающих многими петлями антенны самцов и самок видов семейства Cecidomyidae, исследована у галлицы Contarinis sorghicola. Опоясывающие нити по своему строению подобны тонкостенным обонятельным сенсиллам; их диаметр свыше 1 мкм, а стенки пронизаны порами. На поперечных срезах через нити виден просвет, заполненный ветвями периферических отростков чувствующих клеток.
Чувствительные придатки, или конусы. Антеннальный придаток личинки жука Ctenicera destructor - сложный чувствительный орган, образованный соединением 12 базиконинеских сенсилл. В придатке имеются 36 биполярных чувствующих клеток, распределенных в 12 групп по три в каждой. Кроме них в состав чувствительного придатка входят по 12 трихогенных, тормогенных, трофических и нейриллемных клеток. Все клетки, принадлежащие одной элементарной сенсилле, отделены от соседних мембрановидной перегородкой. В конусе периферические отростки чувствующих клеток ветвятся и заполняют все его пространство. Кутикула конуса пронизана небольшими трубочками, диаметр которых равен 300 Å. Непосредственной связи периферических отростков с трубочками не обнаружено.
Кутикулярные выросты, напоминающие базиконические сенсиллы и связанные с 30 чувствующими клетками, описаны на антенне муравья Camponotus vagus. Поры и поровые трубки имеются только в апикальной части выростов. Кольцевидное утолщение стенок выростов делит внутреннюю полость сенсиллы на две камеры. В нижней камере находится специфическая жидкость, через которую должны пройти молекулы пахучих веществ, прежде чем достигнут периферических отростков чувствующих клеток.
Постантеннальный орган ногохвосток по своему строению отличается от обычной схемы строения сенсиллы. В постантеннальном органе Onychiurus sp., имеющем предположительно хемо- или гигрорецепторную функцию, перикарионы чувствующих клеток локализованы в головном мозгу. Имеются и другие ультраструктурные особенности, выделяющие этот орган среди остальных сенсилл: наличие двух зон ресничек в периферических отростках и отсутствие корешковых структур. Сходный орган описан также и у бессяжковых из семейства Acerentomidae.
Целосферические сенсиллы обнаружены только на антеннах жуков Necrophorus sp. Кутикулярный отдел целосферических сенсилл, имеющий вид сферы, полностью погружен в углубление кутикулы. Углубление открывается наружным отверстием диаметром 1-2 мкм. Стенка кутикулярного отдела пронизана порами, с которыми связаны мешковидные образования из волокнистого материала. В сенсиллах первого подтипа эти образования оканчиваются во впячиваниях третьей обкладочной клетки, которая выстилает внутреннюю сторону кутикулярного отдела сенсиллы. В сенсиллах второго подтипа мешковидные структуры окаймляют наружную полость сенсиллы. В сенсилле найдены две чувствующие клетки, периферические отростки которых окружены со всех сторон сколопоидной оболочкой. По видимому, проникновение молекул пахучих веществ к ветвям периферических отростков осуществляется путем диффузии через сколопоидную оболочку.
Сложные чувствительные органы сконцентрированы на флагеллуме антенн цикады Pyrops candelaria. Они имеют вид округлых участков сильно складчатой перфорированной кутикулы. Общее число пор на пластинке достигает 40000-160000. В состав каждой сенсиллы, считаемой сложным обонятельным органом, возникшим из нескольких базиконических сенсилл, входит большое число чувствующих клеток, собранных в 25-35 групп по 6-10 клеток в каждой. Периферические отростки каждой группы окружены чехлом, который образован базальной клеткой. В терминальной части периферические отростки ветвятся, причем каждая ветвь связана с несколькими порами.
Стимулопроводящая система обонятельных сенсилл. За исключением отдельных примеров, все перечисленные типы обонятельных сенсилл характеризуются пористостью стенок кутикулярного отдела. Для того чтобы вступить во взаимодействие с рецепторными зонами мембран чувствительных сенсилл, молекулы раздражителя должны проникнуть во внутреннюю полость сенсиллы, чему способствует особая стимулопроводящая система, включающая в себя поры и соединенные с ними поровые трубочки или филаменты в стенке сенсиллы.
В обонятельных волосках обнаружено несколько типов организаций стимулопроводящей системы. В толстостенных сенсиллах на антенне таракана Arenivaga sp. описаны поровые каналы диаметром 100-200 Å и длиной до 1 мкм, соединяющие с внешней средой полость волоска, где находятся периферические отростки. Проксимальные части каналов часто бывают расширенными, образуя капсулы, заполненные электроннопрозрачным содержимым. Внутренняя поверхность стенок каналов выстлана электронноплотной пленкой. Использованием различных режимов фиксации препаратов, действием органических растворителей и гидрофильных красителей была показана неоднородность содержимого каналов. Оказалось, что капсулы заполнены липидами, а электронноплотная пленка представляет собой липопротеиновую мембрану, которая выстилает и внутреннюю полость волоска.
Большинство обонятельных сенсилл насекомых имеет тонкие стенки кутикулярного отдела. В обонятельных сенсиллах кровососущих двукрылых и самцов бабочек сатурний наиболее типичное строение поры - коническая полость, пронизывающая стеику сенсиллы и открывающаяся наружу узким отверстием (около 100 Å). Полость содержит 4-8 поровых трубочек, обычно выходящих из нее в просвет сенсиллы и во многих случаях вступающих в контакт с мембранами разветвлений периферических отростков. Отверстие поры может находиться в углублении, в отдельных случаях принимающем вид капсулы. При этом дно капсулы образуется периферическими отростками поровых трубочек.
Проводящая роль поровых трубочек в антеннальных сенсиллах жуков Necrophorus vespillo была показана Эрнстом в 1969 г. Он наблюдал проникновение частиц коллоидного серебра (протаргола) в просвет трубочки, хотя выхода частиц в полость сенсиллы не происходило. Аналогичная картина была описана для поро-трубчатой системы дорсального органа личинки мухи Musca domestica, а также для поровых каналов толстостенных обонятельных сенсилл клеща Amblyomma americanum. Вместе с тем обонятельные сенсиллы органа Галлера этого клеща, видимо, имеют гидрофобное содержимое пор, так как гранулы серебра скапливаются лишь в начале поры. Длительная экспозиция в растворе протаргола приводила к возникновению электронноплотных филаментов из мелкодиспергированного материала в порах, по размерам и расположению очень похожих на поровые трубочки насекомых, но без центральной светлой части. Эти филаменты могли достигать мембран разветвлений периферического отростка чувствительной клетки сенсиллы.
Присутствие тяжей, или филаментов, в порах тонкостенных обонятельных сенсилл было показано у таракана Arenivaga sp. Тяжи имеют липидную природу, так как растворяются в ацетоне, исчезают при изменении режима фиксации, ведущему к вымыванию липидов, не пропускают в полость волоска через поры водорастворимые красители.
Существуют две точки зрения на связь поровых трубочек, или филаментов, с кутикулой. С одной стороны, считают, что трубочки представляют собой вещество непрерывной мембраны, подстилающей кутикулу волоска сенсиллы изнутри. С другой стороны, вещество, из которого состоят трубочки, отождествляют с эпикутикулой волоска. Так, обонятельные сенсиллы тутового шелкопряда имеют трехслойную эпикутикулу. Наружный электронноплотный слой не заходит в пору. Средний слой, прозрачный на электронных микрофотографиях, заполняет сердцевину микротрубочки, а внутренний слой эпикутикулы, по плотности равный наружному, формирует стенки микротрубочки. Поскольку материал, составляющий содержимое и стенки микротрубочек, у этого насекомого не растворяется в органических растворителях, ему приписывают свойства "стабилизированных липидов", состоящих из полимерных единиц. Если учесть, что значительная часть поровых трубочек (на некоторых срезах волоска до 40%) вступает в контакт с мембранами периферических отростков чувствующих клеток, то можно говорить о прямой связи в виде липидного мостика поверхности кутикулы волоска, куда попадают молекулы пахучих веществ, с рецепторной мембраной. Установлено, что сенсиллы на антеннах самца тутового шелкопряда адсорбируют 80-100% бомбикола попавшего на антенну, хотя их поверхность составляет лишь около 13% площади антенны. Количественные измерения порога чувствительности самца тутового или непарного шелкопрядов к феромонам самки, полученные при использовании как поведенческого, так и электрофизиологического критериев, показали, что возбуждение самца наступает, когда на антенны действует струя воздуха, содержащая около 100 молекул аттрактанта в 1 см3 воздуха. Если допустить, что все молекулы адсорбируются и попадают на поверхность волосков и учесть площадь поперечного сечения поровых каналов, которая составляет 0,03% от площади волосков, то за 30 с (максимальное время латентного периода реакции) при скорости потока 1 см/с вероятность прямого попадания хотя бы одной молекулы в пору равна 0,1. Расчеты приводят к выводу, что стимулопроводящая система должна активно участвовать в транспортировке молекул раздражителя не только внутри порового канала и поровых трубочек, но и на поверхности волоска.
Несомненная общность строения стимулопроводящей системы в обонятельных сенсиллах у большого числа исследованных представителей различных отрядов насекомых говорит об ее важной роли в первичных процессах обонятельной рецепции членистоногих. Очевидно, что функциональная роль поро-трубчатой системы у насекомых тесно связана с явлением сверхвысокой чувствительности к специфическим запахам, поскольку в обонятельных органах позвоночных аналогичных морфологических образований не обнаружено.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'