2. Морфологическая характеристика контактных хеморецепторных сенсилл
Локализация. Основное назначение контактных хеморецепторных органов насекомых - определение пригодности субстрата для питания и яйцекладки. Поэтому они встречаются на тех участках тела насекомого или его придатках, которые используются для отгрызания и размельчения пищи, а также для ощупывания поверхности предметов, т.е. на челюстях, околоротовых придатках, яйцекладах и дистальных члениках ног. Для того чтобы определить локализацию контактных хеморецепторов, растворы различных вкусовых веществ наносили на различные участки тела насекомого и регистрировали его ответную реакцию, которая проявлялась в характерных рефлекторных движениях ротовых частей, хоботка, околоротовых придатков. В случае, если отдельные сенсиллы были хорошо различимы при малых увеличениях микроскопа (лабеллярные волооки высших мух), производили раздражение отдельных чувствительных волосков растворами соли или сахара, заключенными в микропипетку. Окончательная идентификация чувствительных волосков как контактных хеморецепторов может быть осуществлена по методике, предложенной в 1954 г. американскими исследователями Е. Ходжсоном и К. Редером, когда микропипетка с раздражающим раствором одевается на вершину волоска и одновременно служит электродом, регистрирующим биоэлектрическую активность чувствующих клеток сенсиллы.
Многочисленные исследования показали, что контактные хеморецепторы у насекомых из разных отрядов в основном локализованы на лабиальных и максиллярных пальцах, челюстях, лигуле и гипофарингсе. У насекомых, имеющих хоботок, контактные хеморецепторы хорошо развиты на дистальных члениках ног, преимущественно на лапках. Они обнаружены также на антеннах пчел, муравьев, кузнечиков, термитов, бабочек из семейства совок, на яйцекладах мух, орехотворок и наездников-ихневмонид.
У насекомых с ортоптероидным типом ротового аппарата (тараканы, прямокрылые, многие жуки) все его части - мандибулы, макоиллы, верхняя и нижняя губы - участвуют в откусывании и размельчении пищи. Хеморецепторы в этом случае диффузно и равномерно распределены по внутренней поверхности ротовых придатков. У саранчи Schistocerca gregarina мелкие трихоидные сенсиллы размером 35 мкм присутствуют на всех частях ротового аппарата, а базиконические сенсиллы найдены на клипеолабральном комплексе и гипофарингсе. На эпифарингсе личинок стрекоз обнаружено множество сенсилл, представляющих собой выпуклые участки мезокутикулы с центральной порой, куда подходят периферические отростки чувствующих клеток. У имаго стрекоз поверхность эпифарингса покрыта многочисленными базиконическими сенсиллами.
У гусениц бабочек контактные хеморецепторы сконцентрированы на дистальных участках максиллярных и лабиальных пальп и представлены базиконичеекими и стилоконическими сенсиллами. Аналогичное распределение вкусовых сенсилл было обнаружено у личинок щелкунов рода Agriotes и личинок хлебного жука Anisoplia austriaca. В последнем случае стилоконичеокие сенсиллы присутствуют также на максиллах и на внутренней поверхности верхней губы личинки жука. У имаго бабочек контактные хеморецепторы найдены на лабеллах и лапках ног и отсутствуют на лабиальных и максиллярных пальпах. Сходная картина наблюдается у Thysanura и Diptera.
У высших двукрылых вкусовые хеморецепторы расположены на вентролатеральной поверхности лабелл и представлены трихоидными сенсиллами разного размера. Самые длинные волоски у мух Calliphora erythrocephala достигают 300 мкм и расположены по периферии орального диска. Волоски (внутренних рядов имеют меньшие размеры, а на вентральной поверхности хоботка встречаются маленькие, около 10 мкм, "интерпсевдотрахеальные палиллы". Такое различие в длине волосков рассматривается как приспособление для увеличения числа контактов рецепторов с поверхностью во время приема пищи, когда лабеллы находятся в развернутом состоянии. Общее количество лабелляряых трихоидных сенсилл у мух колеблется от 220 у Calliphora vomitoria до 28-30 у Stomoxys calcitrans.
Еще в 1926 г. Д. Минничем было показано, что мухи вытягивают хоботок при контакте дистальных сегментов йог с раствором сахара. Позже тарзальная чувствительность была обнаружена у комаров, слепней, клопов, бабочек, саранчовых и многих других насекомых. По своей морфологии тарзальные хеморецепторы мало отличаются от трихоидных лабеллярных сенсилл. Однако число контактных хеморецепторов на лапках трех пар ног неодинаково. Так, тарзусы передних ног мухи Phormia regina несут 303 сенсиллы, а тарзусы задних ног - 147 сенсилл. Такая же особенность распределения тарзальных сенсилл установлена для осенней жигалки Stomoxys calcitrans, кровососущих комаров Aedes aegypti, некоторых видов слепней. Увеличение числа сеноилл на лапках передних ног свидетельствует об их большей чувствительностя к контактным раздражителям, что также подтверждается физиологическими тестами.
У насекомых с колющесосущим ротовым аппаратом (комары, тли, клопы) контактные хеморецепторные органы можно разделить на две группы соответственно выполняемым функциям. Это, во-первых, трихоидные и базиконические сенсиллы внешней поверхности нижней губы, используемые при ощупывании поверхности субстрата и при питании свободными жидкостями, и, во-вторых, сенсиллы колющих стилетов и стенок пищевого канала, функционирующие при проколе тканей.
Контактные хеморецепторные органы насекомых представлены четырьмя типами сенсилл: трихоидными, базиконическими и стилоконическими, а также чувствительными папиллами.
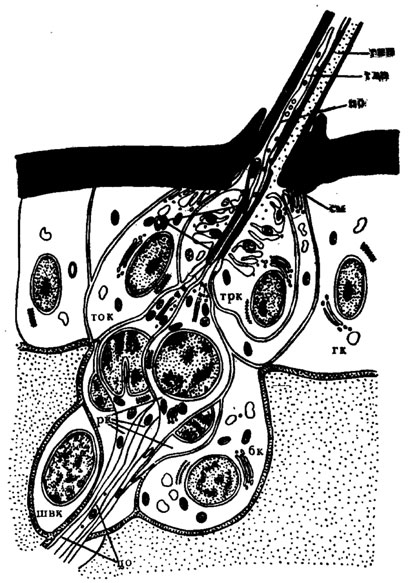
Рис. 24. Схема строения контактной хеморецепторной трихоидной сенсиллы мошки Boophthora erythrocephala (по Елизарову, Чайке, 1975): по - периферический отросток; рк -рецепторная клетка; см - сочленовная мембрана; со - сколопоидная оболочка; тлп - толстостенная полость; тнп - тонкостенная полость; ток - тормогенная клетка; трк - трихогенная клетка; цо - центральный отросток; швк - Шванновская клетка
Ультраструктура контактных хеморецепторных сенсилл. Наиболее полно строение и функциональные свойства трихоидных хеморецепторных сенсилл исследованы на примере лабеллярных хеморецепторов двукрылых (рис. 24, 25).
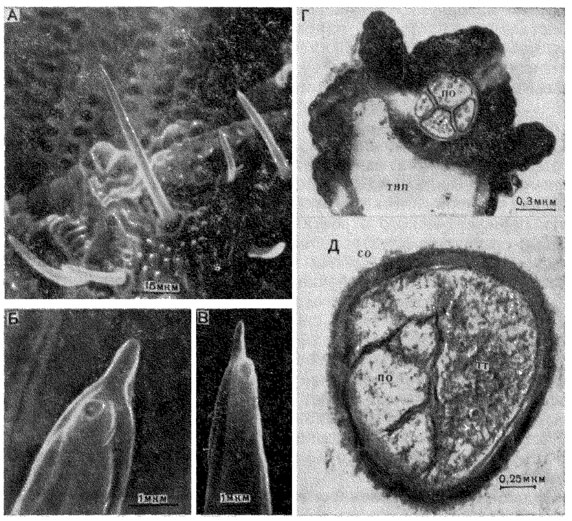
Рис. 25. Контактные хеморецепторные сенсиллы. А - трихоидные лабеллярные сенсиллы на краю орального диска хоботка мухи Protophormia terraenovae; Б - пора на кончике трихоидной сенсиллы; В - вязкое содержимое, выступающее из поры; Г - поперечный срез волоска на лабеллуме комара Culex pipiens; Д - поперечный срез лабеллярной трихоидной сенсиллы мошки Boophthora erythrocephala на уровне трубчатого тела (тт) механорецептора. Обозначения, как на рис. 24
Кутикулярный отдел сенсиллы представляет собой толстостечный волосок, который разделен продольной перегородкой на две полости, названные по толщине ограничивающей их кутикулы: толстостенной и тонкостенной. Перегородка волоска, по-видимому, сколопоидная оболочка, окружает периферические отростки чувствующих клеток и одной стороной опаяна с внутренней поверхностью стенки волоска. В толстостенной полости к вершине волоска проходят периферические отростки клеток. Тонкостенная полость волоска сообщается с межклеточной полостью сенсиллы. Основание волоска связано с кутикулой лабеллума тонкими тяжами, имеющими, по-видимому, кутикулярную природу. Эти тяжи обеспечивают отклонение волоска при контакте с субстратом, а также приведение его в исходное шстояние.
На вершине волоска имеется одна или несколько пор диаметром около 500 Å, соединяющих толстостенную полость с внешней средой. Иногда поры смещены несколько ниже вершины волоска, имеющей в этом случае форму папиллы (рис. 25,Б). Тонкостенная полость заканчивается слепо под вершиной волоска на расстоянии нескольких микрон.
При электронномикроскопическом исследовании вершины хеморецепторной сенсиллы у мух Phormia regina и Calliphora vicina обнаружены два состояния поры: открытое и закрытое. Водные растворы и влажный воздух вызывают открывание поры, тогда как действие сухого воздуха, фиксаторов и прикосновение к волоску приводят к закрыванию поры. Механизмы, осуществляющие открывание и закрывание пор в трихоидных сенсиллах, пока остаются невыясненными. Видимо, они связаны с изменением давления внутри волоска, так как сократительных волокон или каких-либо тяжей в полости волоска не обнаружено. Предполагается, что в кончике волоска в состав кутикулы входит структурный белок "резилин", обеспечивающий подвижность складок, закрывающих пору.
На вершине волоска могут наблюдаться капли вязкой жидкости, выступающей из поры (рис. 25, В). Вязкий субстрат служит посредником между растворами вкусовых веществ и чувствительными мембранами периферических отростков. В отдельных трихоидных сенсиллах отмечаются пучки филаментов из электронно-плотного материала, соединяющиеся с мембраной периферического отростка, что говорит об определенном сходстве с поротрубчатой системой обонятельных сенсилл.
Число рецепторных клеток в контактных хеморецепторных трихоидных сенсиллах не превышает пяти, хотя в отдельных сенсиллах их может быть и меньше. Периферические отростки доходят до вершины волоска, кроме отростка одной из клеток, который заканчивается у основания волоска характерным образованием, называемым "трубчатым телом" (рис. 25, Д). Как показали электрофизиологические исследования, этот отросток принадлежит механорецепторной клетке. Трубочки, содержащиеся в дистальной части периферического отростка механорецептора, имеют диаметр 150 Å и состоят из электронноплотного материала. В центре трубчатого тела они расположены гексагонально, на периферии - палисадно. Между собой трубочки связаны перпендикулярными слоями электронноплотного вещества. Стимуляция механорецепторной клетки происходит в момент сжатия трубчатого тела, возникающего при движении волоска.
Кутикулярный отдел базиконических сенсилл с контактной хеморецепторной функцией представляет собой полый толстостенный конус, погруженный в углубление кутикулы и соединенный с верхним краем расположенного под ним канала сочленовной мембраной. Между основанием конуса и стенками канала может быть натянута тонкая дополнительная мембрана, состоящая из отдельных тяжей.
Отличительной особенностью строения кутикулярного отдела базиконических сенсилл служит отсутствие продольной перегородки в его полости. Периферические отростки чувствующих клеток, окруженные сколопоидной оболочкой, проходят в центре кутикулярного отдела к терминальной поре, диаметр которой около 300 Å. Сколопоидная оболочка прикреплена к стенкам конуса только в области его вершины. Внутри футляра, образованного сколопоидной оболочкой, отходят продольные выросты, частично разделяя периферические отростки клеток.
Базиконические сенсиллы иннервируются тремя-пятью чувствующими клетками. В отдельных сенсиллах, например в терминальных сенсиллах максиллярных пальп саранчи Schistocerca gregarina, их число может доходить до 11 на одну сенсиллу.
Вкусовые папиллы. Интерпсевдотрахеальные папиллы высших мух представляют собой небольшие конусы, не превышающие по высоте 7 мкм. Они находятся в кутикулярных ямках на такой глубине, что вершина конуса слегка выступает над поверхностью кутикулы. Папиллы встречаются у многих мух на вентральной поверхности хоботка в промежутках между псевдотрахеями.
Исследование ультраструктуры папилл мух Boettcherisca peregrina показало большое сходство в строении папилл и контактных хеморецепторных базиконических сенсилл. Элидермальный клеточный комплекс папиллы содержит четыре чувствующие клетки, две трихогенные и одну тормогенную. Одна из чувствующих клеток служит механорецептором и образует трубчатое тело в основании папиллы. Периферические отростки клеток имеют структуру ресничек. Как и в предыдущих типах сенсилл, обкладочные клетки образуют две полости: базальную, где расположены основания ресничек, и межклеточную полость сенсиллы, доходящую до основания кутикулярногр конуса. Дистальные части периферических отростков чувствующих клеток заключены в сколопоидные оболочки, заканчивающиеся у основания папиллы. В узком ее просвете сколопоидных оболочек не обнаружено, дендриты лежат свободно в жидком содержимом кутикулярного отдела. Характер возникновения возбуждения при раздражении клеток папиллы поэтому должен несколько отличаться от процессов, происходящих при возбуждении клеток в двуполостной трихоидной контактной хеморецепторной сенсилле.
Другие типы контактных хеморецепторных сенсилл. Контактная хеморецепторная функция свойственна стило-коническим сенсиллам личинок чешуекрылых и жуков, мелким чувствительным шипам и порам, обнаруженным на клипеолабруме саранчи и в цибариальной полости мух и комаров, а также чувствительным бугоркам на антеннах бабочек-совок, куполообразным сенсиллам на яйцекладах наездников и т.п. Все эти органы имеют поры на вершине кутикулярного отдела, 3-5 чувствующих клеток с цилиарной структурой периферических отростков, обкладочные клетки, образующие межклеточные полости сенсиллы, сколопоидные оболочки, окружающие периферические отростки целиком или в отдельности. Механорецепторов в таких сенсиллах не обнаружено.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'