8. Виброрецепция
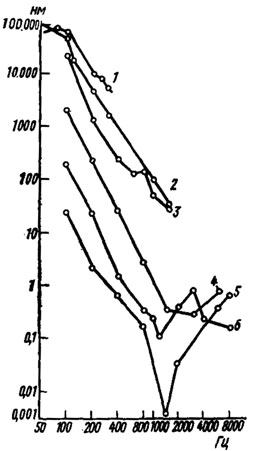
Рис. 21. Частотно-пороговые характеристики ножных виброрецепторов (по Autrum, Schneider, 1948): 1 - Carabus, 2 - Vespa, 3 - Bombus, 4 - Pyrantels, 5 - Periplaneta, 6 - Liogryllus. По оси абсцисс - частота, Гц; по оси ординат - амплитуда вибрации, нм
Многочисленные наблюдения и электрофизиологические данные свидетельствуют о способности насекомых воспринимать вибрации субстрата. Рецепторы, реагирующие на эти колебания, локализуются в ногах*. Регистрируя суммарную активность сенсорных нейронов в ножных нервах, можно получить представление о чувствительности этих рецепторов. За порог реакции принимается то минимальное значение амплитуды смещения субстрата или ускорения, при которых наблюдаются ответы (рис. 21).
* (Исключение составляют только трихоидные сенсиллы на стернитах саранчовых и, возможно, булавовидные хеты на церках сверчковых.)
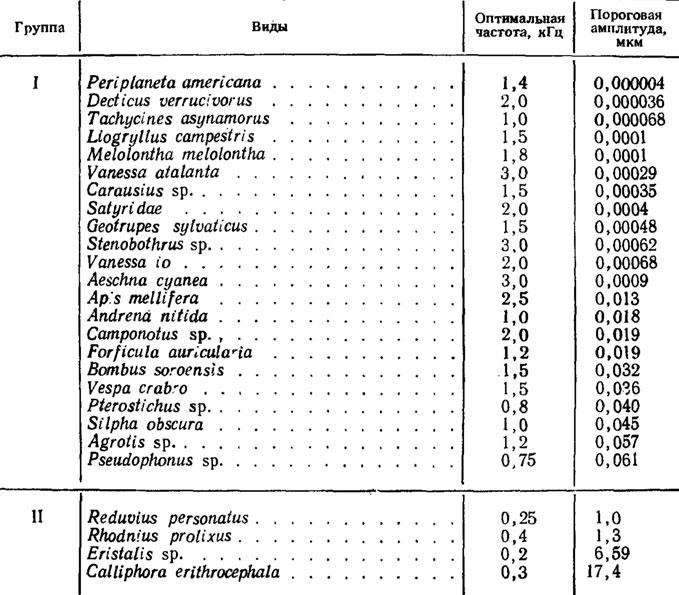
Таблица 2 Чувствительность насекомых к вибрациям (по Schneider, 1950)
В результате сравнительного изучения достаточно большого числа видов насекомых было установлено, что они могут быть условно разделены на две группы: высокочувствительные и малочувствительные (табл. 2). К первым относятся тараканы, прямокрылые, палочники, перепончатокрылые, жесткокрылые и другие, ко вторым - полужесткокрылые, двукрылые и некоторые жесткокрылые. Различия между этими группами касаются не только порогов реакции, но и оптимальных частот. В первом случае они обычно лежат в диапазоне 1-3 кГц, во втором - не превышают 0,4 кГц.
Сравнительно-анатомические материалы и результаты экстирпаций различных ножных рецепторов дают основание считать, что у чувствительных видов восприятие вибраций осуществляется преимущественно субгенуальными органами, а у малочувствительных - ти-биогарзальными хордотональными органами и трихоидными сенсиллами на лапках. Такая закономерность прослеживается при сравнении многих групп насекомых, но ее все-таки нельзя считать универсальной, так как отсутствие субгенуального органа не исключает достаточно высокой чувствительности к вибрациям (Melolontha, Geotrupes), а при его наличии восприятие может осуществляться преимущественно другими рецепторами (см. ниже). Точное выяснение роли отдельных сенсилл или органов в этом процессе сопряжено с большими трудностями, поэтому достоверных данных получено пока немного.
У тараканов, сверчков и термитов колебания субстрата воспринимаются субгенуальными органами. Степень их развития и детали строения сильно варьируют у разных насекомых. Обычно они представляют собой 1-2 группы удлиненных хордотональных сенсилл, расположенных между трахеей и кутикулой в проксимальной части голени. Число сенсилл в органе колеблется от 10 до 40. У тараканов и прямокрылых шапочковые клетки прикрепляются к кутикуле в одной точке, а тела биполярных нейронов образуют дугу, поэтому в целом орган имеет веерообразную форму (см. рис. 13,Б). Несколько наружных, более или менее обособленных сенсилл часто рассматриваются как самостоятельный прилежащий орган.
У тараканов (Periptaneta) ответы субгенуального органа регистрируются в диапазоне 0,025-6 кГц. Оптимум чувствительности лежит в области 1-5 кГц. Минимальные пороги реакции достигают 10-7-10-10 см. Прилежащий орган воспринимает только низкочастотные колебания в пределах 30-700 Гц и отличается меньшей чувствительностью (пороги-10-6 см). Помимо хордотональных органов на вибрации субстрата отвечают также кампаниформные сенсиллы, расположенные в проксимальной части голени, но они возбуждаются достаточно сильными колебаниями с частотой, не превышающей 100 Гц.
Сходные результаты были получены при изучении ножных рецепторов сверчков (Gryllus)*. Оптимум чувствительности субгенуальных органов передних ног лежит в диапазоне 0,8-1 кГц, а задних - 0,4-0,5 кГц. Минимальные пороги (по амплитуде смещения) во всех трех случаях различались мало (1,4Х10-8-2,2Х10-8 см), но передние рецепторы оказались менее чувствительными к ускорению. Искусственное утяжеление задних ног не влияло на функции рецепторов, поэтому можно считать, что их характеристики определяются только механическими свойствами самих субгенуальных органов. Сравнение органов трех пар ног показывает, что их оптимальные частоты находятся в обратной зависимости от собственной массы.
* (Помимо реакции субгенуальных органов в передних ногах регистрировали так же ответы каких-то других низкочастотных механорецепторов, расположенных проксимальнее тибиотарзального сочленения.)
Эти данные находятся в соответствии с модельными представлениями, согласно которым субгенуальный орган рассматривается как эластичная мембрана с массой в центре, натянутая в сосуде с жидкостью. Сотрясение такой системы вызывает смещение массы и колебания мембраны, регистрируемые измерителем (сенсиллами).
У муравьев-листорезов (Atta) ножные рецепторы реагируют на звуки в диапазоне 0,05-4 (7) кГц. Оптимум чувствительности по амплитуде смещения лежит в области 1-3 кГц (минимальный порог - 1,ЗХ10-7 см), а по ускорению - 0,1-2 кГц (порог - 2,5 см/с2). Наиболее чувствительные сенсиллы находятся в передних ногах, причем солдаты менее восприимчивы к вибрациям, чем рабочие. Селективное разрушение разных рецепторов показало, что на колебания субстрата реагируют главным образом кампаниформные сенсиллы, расположенные в области сочленения вертлуга и бедра. Роль субгенуальных органов в этом случае остается неясной.
Вибрации, воспринимаемые насекомыми в естественных условиях, возникают под действием абиотических факторов (ветер, дождь и т. п.) и в результате активности животных (в том числе и особей своего вида). Обычно сотрясения субстрата происходят при передвижении, питании, прокладывании ходов, строительстве и других видах жизнедеятельности. Но кроме того, многие насекомые специально стучат по окружающим предметам различными частями тела или царапают их мандибулами*. В некоторых случаях субстрат (например сухие листья) служит только для усиления звука, в других же он используется как канал связи, а распространяющиеся в нем колебания служат сигналами, несущими определенную информацию. Такая коммуникация хорошо развита у общественных насекомых (термитов, муравьев, пчел, ос), а также у веснянок (Plecoptera), сеноедов(Psocoptera), жуков-точильщиков (Anobiidae) и др. У одиночно живущих насекомых эти сигналы используются для привлечения особей противоположного пола. Термиты стучат по стенкам гнезда в случае опасности. Воспроизведение этих колебаний вызывает у них серию защитных реакций. Голодные личинки ос получают пищу, если они царапают стенки ячеек мандибулами. Некоторые муравьи (Atta, Megaponera), оказавшись под слоем почвы, издают "сигналы бедствия", привлекающие внимание особей, находящихся на поверхности. Виды, живущие в древесине (Catnponotus), стучат по субстрату при появлении опасности или повреждении гнезда. Очень большую роль подобные сигналы играют в жизни пчел. Малейшие сотрясения улья вызывают у них активную защитную реакцию. Доказано также, что матки медоносной пчелы отвечают на "кваканье" других маток или его имитацию только в том случае, если сигнал передается через субстрат. У безжалых пчел (Melipona) разведчица стимулирует фуражиров на вылет за пищей, издавая специальные звуки. Их восприятие осуществляется виброрецепторами, так как покрытие пола улья резиной полностью исключает реакцию.
* (В некоторых случаях (например у муравьев) через субстрат передаются также колебания, издаваемые фрикционными органами.)
Анализ функциональной организации виброрецедторов свидетельствует о том, что они адаптированы для восприятия коммуникационных сигналов. Так, у термитов (Zootermopsis angusticollis) оптимальная частота сенсилл субгенуальных органов (1,15 кГц) почти совладает с доминирующей частотой сигнала тревоги (1,14 кГц). Аналогичное соответствие имеется между частотным оптимумом рецепторов муравьев (Atta) и спектром прошедших через почву "сигналов бедствия".
Помимо вибраций твердого субстрата насекомые могут воспринимать колебания поверхностной пленки воды. Водомерки (Gerridae) используют такие волны не только для обнаружения добычи (насекомых, плавающих на поверхности), но и для внутривидовой коммуникации в период размножения. Восприятие колебаний осуществляется различными механорецепторами ног. Минимальные пороги их реакций достигают 1 мкм на частоте 200-300 Гц.
У жуков-вертячек (Gyrinidae) эти функции выполняет Джоистонов орган. Второй членик их антенн скользит по воде, а подвижно сочлененный с ним утолщенный жгутик располагается над поверхностью. Колебания второго членика вызывают смещение его относительно жгутика, что приводит к возбуждению хордотональных сенсилл. В целом антенна представляет собой вибрационную систему с резонансной частотой, близкой к 250 Гц. В области частот, не превышающих 150 Гц, она работает как измеритель ускорений. Согласно электрофизиологическим данным, рецептор реагирует на колебания, амплитуда которых достигает нескольких микрон, причем его чувствительность возрастает с увеличением частоты. Во время движения жуки могут определять искривления поверхностной пленки (мениск) вблизи преград с расстояния 0,5- 1,5 см и избегать столкновений.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'