7. Пространственный слух
Слуховые рецепторы. В 1940 г. Памфри высказал предположение, что все слуховые рецепторы насекомых реагируют не на звуковое давление (как у позвоночных животных), а на смещение, или градиент давления. Приемники такого типа обладают направленной чувствительностью (направленностью), что создает теоретическую возможность монауральной локализации источника звука. Бели звуковые колебания воспринимаются более или менее открытой с двух сторон мембраной (тимпанальные органы), то слуховой орган должен обладать независимой от частоты звука двусторонней характеристикой направленности (при длинах волн, не сравнимых с его величиной). В тех случаях, когда звуковые колебания приводят в движение не мембрану, а легкий подвижный стержень (трихоидные сенсиллы, Джонстоновы органы), характеристики направленности приемника должны существенно зависеть от его конструктивных особенностей (формы, способов прикрепления подвижных структур я др.).
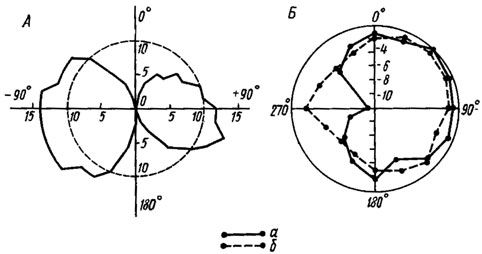
Рис. 19. Характеристики направленности слуховых органов: А - изолированного тимпанального органа саранчи (по Pumphrey, 1940); Б - тимпанального органа сверчка Gryllus bimaculatus (по Жатнневу и др., 1975). а, б - до и после удаления контралатеральной передней ноги. Частота звука 5 кГц, интенсивность 84 дБ. По радиусам отложена амплитуда ответов в дБ относительно максимума
При экспериментальной проверке этой гипотезы вначале были получены положительные результаты. Так, оказалось, что комары могут находить источник звука с помощью одного слухового органа. Ранее аналогичные опыты были поставлены на сверчках. Электрофизиологичеокие исследования показали, что изолированный тимпанальный орган саранчи обладает двусторонней направленностью (рис. 19, А) и, следовательно, может рассматриваться как приемник градиента давления. Такие же результаты были получены при изучении слуховых органов кузнечиков. Правда, изменение их чувствительности было незначительным, но ее уменьшение при фронтальной стимуляции препарата позволяло думать, что и эти слуховые органы реагируют на градиент давления.
В дальнейшем, однако, стали накапливаться данные, противоречащие упомянутой гипотезе Памфри или ставящие под сомнение ее универсальность. Так, в поведенческих экспериментах не подтвердилась способность некоторых сверчков и кузнечиков к монауральной локализации источника звука. Исследования зависимости направленности тимпанальных органов саранчи от частоты звука дали противоречивые результаты. В некоторых экспериментах диаграммы направленности изменялись с изменением частоты, в других такой закономерности не наблюдалось. Специальное изучение этого вопроса показало, что тело саранчи оказывает существенное влияние на прохождение звуковых волн, в результате чего тимпанальные органы на высоких частотах (10-20 кГц) работают как приемники давления, а на низких (около 3,5 кГц) в какой-то мере реагируют и ,на градиент давления.
Некоторые особенности строения и положения тимпанальных органов кузнечиков давали основания предполагать, что они в значительно большей степени соответствуют требованиям, предъявляемым к идеальным приемникам градиента давления. Однако исследования зависимости характеристик их направленности от частоты звука показали, что в диапазоне 5-8 кГц они имеют почти круговые диаграммы и, следовательно, ведут себя как приемники давления, а на частотах 10-20 кГц диаграммы приобретают форму эллипса, что указывает на возможность восприятия градиента давления.
Характеристики направленности тимпанальных органов сверчков изучали только на оптимальных частотах - в области 5 и 16 кГц. При вращении препарата в звуковом поле перепады чувствительности достигали 6-11 дБ. Полученные диаграммы приближались по форме к кардиоиде (5 кГц) или эллипсу (16 кГц). После удаления контралатеральной ноги и закрывания отверстия трахеи в трохантере чувствительность ипсилатерального органа на 5 кГц возрастала и асимметрия диаграмм уменьшалась (рис. 19, Б). Отсюда возникло предположение, что звук, приходящий по трахее с контралатеральной стороны, ослабляет амплитуду колебаний тимпанальной мембраны.
Для снятия характеристик направленности тимпанальных органов чешуекрылых (Noctuidae) использовали короткие щелчки. Полученные диаграммы показали, что пороги реакции снижались при стимуляции рецепторов под углом 90° к плоскости мембраны и значительно возрастали при переносе источника звука на противоположную сторону тела. Учитывая частотные характеристики тимпанальных органов, можно предполагать, что их направленность возникает в результате экранирующего влияния тела бабочки. В других - опытах было установлено, что ответы этих рецепторов зависят не только от угла, под которым приходит звук, но и от положения крыльев.
Суммируя результаты описанных выше экспериментов, можно прийти к выводу, что тимпанальные органы насекомых не подчиняются закономерностям, характерным для простых приемников градиента давления.
В отличие от тимпанальных органов трихоидные сенсиллы и Джонстоновы органы в силу своих конструктивных особенностей могут реагировать только на смещение. Действительно, звуковые колебания воспринимаются в этих органах тонкими волосками или жгутиками антенн, подвижно сочлененными с кутикулой с помощью эластичной мембраны. В звуковом поле переменный ток воздуха захватывает эти образования внутренним трением и увлекает их в направлении колебания частиц. Если бы такие органы были снабжены рецепторами, реакция которых отражала направление колебаний подвижных элементов, то с их помощью можно было бы успешно локализовать источник звука. Как ранее отмечалось, в трихоидных сенсиллах на церках тараканов имеются структуры, ограничивающие колебания волоска в определенной плоскости, а возбуждение нейрона наступает только в том случае, если волосок отклоняется в одном из двух возможных направлений. Имея набор по-разному ориентированных сенсилл такого типа, можно довольно точно определить направление на источник стимуляции (колебания воздуха и, возможно, низкочастотные звуки).
Значительно больший интерес в этом отношении представляют Джонстоновы органы, в состав которых входит несколько групп хордотональных сенсилл. Результаты их исследования у комаров электрофизиологическими методами в основном сводятся к следующему.
При помещении электрода во второй членик антенны и стимуляции препарата звуками низких частот (60-600 Гц) удается зарегистрировать суммарный синхронный ответ. Амплитуда регистрируемого потенциала в определенных пределах зависит от интенсивности звука и его частоты. Максимальная амплитуда ответа наблюдается при частотах, близких к резонансной частоте антенны. Регистрируемый потенциал иногда помимо основной частоты содержит еще и первую гармонику. Специально проведенные эксперименты показали, что гармоника появляется в том случае, когда звук падает на антенну под углом. Если же фронт звуковой волны перпендикулярен к антенне, то ответ содержит только основную частоту. При стимуляции антенн комара иглой, соединенной с прибором, сообщавшим ей синусоидальные колебания, было установлено, что амплитуда первой гармоники и ее отношение к амплитуде основной частоты зависят от угла, под которым совершаются движения иглы.
На основании этих данных была предложена следующая теория работы Джонстонова органа. При движении антенны вдоль ее продольной оси базальная пластинка (и отходящие от нее отростки) стимулирует сенсиллы радиальной серии, в результате чего возникает суммарный потенциал, частота которого соответствует частоте стимулирующего звука. Если фронт звуковой волны параллелен оси антенны, то она совершает качательные движения, а базальная пластинка попеременно стимулирует две группы хордотональных сенсилл, что приводит к удвоению частоты регистрируемого потенциала. В промежуточных случаях, когда звук падает на антенну под углом меньше 90°, в этом потенциале наблюдаются колебания двух частот (основной частоты и первой гармоники), причем соотношение их амплитуд прапорционально углу, под которым действует стимул.
В процессе дальнейших исследований было установлено, что соотношение амплитуд основной частоты и первой гармоники зависит не только от направления стимуляции, но и от частоты звука, а также от положения электрода во втором членике антени. Поэтому данная теория в настоящее время нуждается в серьезных поправках.
Центральные слуховые нейроны. Тимпанальные органы у насекомых разделены расстоянием от нескольких миллиметров до 3-4 см, поэтому существует возможность восприятия и оценки различий в их стимуляции. При этом теоретически могут быть использованы фазовые, временные и амплитудные различия.
Исследование функциональной организации тимпанальных органов показало, что они не имеют механизмов, необходимых для кодирования и передачи информации о фазовых изменениях звука, поэтому для предположений об использовании этого параметра в процессе локализации нет никаких оснований.
Интерауральные различия по времени могут достигать у наиболее крупных насекомых 0,1-0,15 мс. В большинстве же случаев они, повидимому, не превышают 10-30 мкс. Возможность использования столь малых временных различий* в пространственном слухе насекомых по ряду причин представляется маловероятной и требует специального доказательства.
* (Значения интерауральных временных различий даны для случая, когда звук падает под углом 90° к продольной оси тела насекомого. При локализации источника звука отклонения от курса обычно не достигают 30-40°, поэтому эти различия будут еще меньшими.)
Различия в интенсивности стимуляции симметричных тимпанальных органов возникают по двум причинам: из-за их направленной чувствительности и в результате экранирующего влияния тела насекомого. Влияние последнего фактора становится заметным при длинах волн, сравнимых с телом насекомого, и возрастает с ростом частоты.
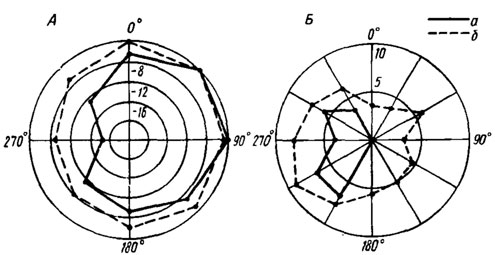
Рис. 20. Изменения ответов слуховых ин-тернейронов в грудном отделе ЦНС при вращении насекомого в звуковом поле. А - ответы нейрона С сверчка Gryllus bimaculatus (по Жантневу и др., 1975): а, б - до и после перерезки контралатерального тимпанального нерва. Частота 5 кГц, интенсивность 57 дБ. По радиусам отложена интенсивность ответа в дБ относительно максимума. Б - ответы нейрона F саранчи Locusta migratoria (по Kalmring et al., 1972): а, б - до и после разрушения контралатерального тимпанального органа. Частота звука 12 кГц, интенсивность 63 дБ. По радиусам - число импульсов в ответе на звуковую посылку длительностью 20 мс
Электрофизиологические исследования, выполненные в течение (последнего десятилетия, позволили установить наличие перекрестных тормозных и возбуждающих связей между симметричными отделами слуховых анализаторов в грудных и головных ганглиях (рис. 20). Впервые такие связи были обнаружены при изучении слуховой системы кузнечиков, впоследствии их удалось найти у саранчовых, сверчков, медведок и чешуекрылых.
Сравнение полученных к настоящему времени фактов позволяет наметить некоторые общие закономерности в реакциях центральных слуховых нейронов при изменении интерауральных различий в стимуляции.
Прежде всего следует отметить, что ответы многих интернейронов зависят от положения источника звука в пространстве и закономерно изменяются при его перемещении* (или вращении насекомого), в результате чего могут быть получены соответствующие диаграммы (см. рис. 20). Наблюдающиеся при этом различия в ответах касаются как частоты импульсации, так и величины скрытых периодов реакции. Различия в порогах реакции симметричных нейронов колеблются в значительных пределах и могут достигать 40-60 дБ. Наряду с этим встречаются клетки, совершенно не реагирующие на вращение насекомого в звуковом поле.
* (Здесь и далее речь идет о перемещениях в горизонтальной плоскости.)
Контралатеральный тимпанальный орган оказывает на активность нейронов как тормозное, так и возбуждающее воздействие.
Обычно одно из них преобладает, а их соотношение зависит не только от интерауральных различий в стимуляции, но и от интенсивности, а иногда и частоты звука. В некоторых случаях наблюдаются "парадоксальные" реакции, когда усиление стимуляции ипсилатерального органа вызывает повышение импульсации контралатеральных интернейронов.
Значительный интерес представляют клетки, ответы которых изменяются только при изменении интерауральных различий стимуляции и не зависят (в широких пределах) от интенсивности звука.
Использование ритмично повторяющихся звуковых посылок, особенно следующих с частотой повторения пульсов в видоспецифических сигналах, существенно усиливает различия в ответах симметричных нейронов.
Сравнение реакций слуховых интернейронов грудных и головных ганглиев свидетельствует о том, что между ними нет принципиальных различий, но асимметрия в активности протоцеребраль ных нейронов может быть выражена в меньшей степени, чем у элементов из грудных ганглиев.
Исследование центральных отделов слуховых анализаторов насекомых началось сравнительно недавно, поэтому многие особенности их функционирования при локализации источника звука остаются еще неизученными. Тем не менее проведенные эксперименты позволили выявить два важных факта: существование перекрестных связей на нескольких уровнях слуховых анализаторов и участие в них многих нейронов с разными характеристиками. Сопоставление этих данных с результатами этологических опытов дает основание предполагать, что реципрокные влияния (особенно тормозное, см. рис. 20) повышают контрастность в реакциях симметричных нейронов, в результате чего, по-видимому, увеличиваются разрешающая способность системы и точность локализации, а использование нескольких каналов с разными перекрестными связями повышает пластичность этого механизма и помехоустойчивость при изменяющихся условиях ориентации.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'