6. Механизмы опознавания коммуникационных звуковых сигналов
О том, что насекомые слышат и опознают коммуникационные звуковые сигналы, свидетельствуют реакции двух основных типов: изменение акустической активности (фонореакция) и двигательные реакции (фонотаксис). В первом случае наблюдаются активация, торможение, альтернация (чередование) или синхронизация пения. Во втором - приближение, удаление или иные перемещения насекомого относительно источника звука.
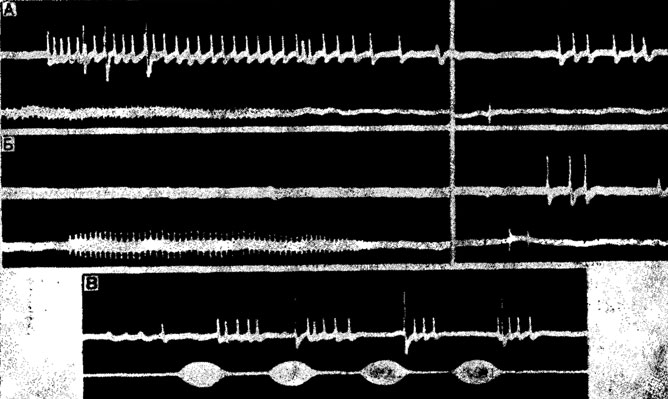
Риc. 18. Ответы восходящих слуховых интернейронов из шейных коннективов кузнечика lsophya taurica (А, Б) и сверчка Gryllus bimaculatus (В) на естественные звуковые сигналы. А - ответы тонического нейрона и детектора пульсов (импульсы большей амплитуды) на призывный (слева) и ответный (справа) сигналы; Б - ответы детектора щелчков на те же сигналы; В - ответы нейронов В и С на призывный сигнал
Избирательность акустических реакций можно легко наблюдать у видов, издающих призывные и ответные звуки. Так, самки кузнечиков рода Isophya отвечают на каждый пульс сигнала самца одним-тремя короткими щелчками (рис. 18, А, Б). Помещая перед садком с половозрелыми самками поющих самцов разных видов прямокрылых, в том числе и близких симпатрических видов Isophya, нетрудно заметить, что ответные сигналы издаются только на призывные звуки своего вида.
Многочисленные исследования положительного фонотаксиса у разных групп прямокрылых дают убедительные доказательства того, что самок привлекают видоспецифические призывные сигналы. Реакции обоих типов сохраняются и при воспроизведении сигналов, записанных на магнитную ленту, поэтому нет оснований сомневаться в том, что насекомые реагируют именно на звук, а не на какие-то иные факторы.
Лабораторные эксперименты показывают, что насекомые не только отличают видоспецифические сигналы от других звуков, предъявляемых последовательно, но и опознают их при одновременном звучании, т. е. выделяют полезный сигнал из шума. В результате специального изучения помехоустойчивости акустической системы связи у кузнечиков рода Metrioptera было установлено, что самки проявляют положительный фонотаксис даже в том случае, если шум (звук симпатрического вида) превышает по интенсивности сигналы "своих" самцов в 3 раза. Совершенно очевидно, что такая помехоустойчивость обеспечивает надежную связь в естественных биотопах, где одновременно издают звуки многие виды животных.
Как ранее отмечалось, некоторые насекомые издают не один, а несколько (до семи) коммуникационных сигналов. Их воспроизведение вызывает разные реакции, поэтому есть основания считать, что слуховая система не только выделяет эти сигналы из шума, но и отличает их друг от друга.
Как же насекомые опознают звуковые сигналы?
Для того чтобы ответить на этот вопрос, необходимо прежде всего выявить те признаки, или параметры, сигналов, которые используются для их различения и опознавания. Результаты сравнительного акустического анализа показывают, что частотные спектры сигналов близких, и в том числе симпатрических, видов широко перекрываются, а их временные характеристики (длительность и закономерности чередования пульсов), напротив, обладают высокой видоспецифичностью и хорошо различаются (см. рис. 12, А). Это дает основание предполагать, что насекомые используют для опознавания акустических сигналов не частотные, а временные параметры.
Тщательный анализ сигналов, зарегистрированных при разных условиях или полученных от разных особей одного вида, свидетельствует о том, что временные параметры существенно различаются по степени изменчивости. Поэтому для опознавания сигналов, по-видимому, должны использоваться не все, а только некоторые, наиболее стабильные элементы.
Для выяснения реальной роли тех или иных акустических компонентов в процессе опознавания проводят специальные этологические эксперименты, в которых сравнивают реакцию насекомых на естественные и искусственные звуки. Обычно для этого используют одну из описанных выше реакций - звуковые ответы или фонотаксис. Сначала активным насекомым воспроизводят естественные (чаще всего призывные) сигналы, а затем их определенным образом трансформируют или заменяют имитациями (моделями) с изменяемыми параметрами. Так, например, сверчкам, издающим призывные звуки, которые состоят из четырехпульсовых серий, предъявляют модельные сигналы; в них произвольно изменяются длительность пульсов, частота их повторения, число в серии, длительность серий и т. д.
Результаты таких экспериментов, проведенных на разных группах прямокрылых, показали, что временная организация сигнала действительно играет решающую роль в опознавании, но разные виды насекомых могут использовать для этого разные компоненты. Так, самки симпатрических видов рода Homorocoryphus различают призывные трели самцов только по частоте повторения пульсов. У некоторых стеблевых сверчков (Oecanthus) сигналы опознаются главным образом по частоте повторения серий. Сверчки рода Gryllus используют для этой цели частоту повторения пульсов и длительность серий, а некоторые виды кузнечиков рода Isophya - длительность пульсов.
В течение длительного времени принято было считать, что насекомые не могут распознавать звуки по частоте. Однако результаты новейших исследований свидетельствуют о том, что частотные параметры все же используются для опознавания некоторых коммуникационных сигналов. При рассмотрении этого вопроса необходимо прежде всего помнить, что слуховые органы обладают определенной частотной избирательностью, поэтому диапазон воспринимаемых ими частот может не перекрываться со спектрами сигналов некоторых групп насекомых. Так, например, многие кузнечики, по-видимому, почти не слышат призывных сигналов сверчков, что, несомненно, облегчает их звуковую коммуникацию в естественных условиях. Некоторые листовые кузнечики (Isophya) издают звуки в сравнительно узкой полосе частот и обладают соответствующим образом "настроенными" рецепторами, позволяющими использовать частоту для опознавания видоспецифических призывных звуков. Их самки отвечают на модельные сигналы только в том случае, если их частотные характеристики приближаются к естественным.
Этологические опыты, проводившиеся на сверчках из родов Oecanthus и Scapsipedus, показали, что изменение частоты в диапазоне, доступном для слуховых рецепторов, не оказывает существенного влияния на привлекательность искусственных призывных сигналов для самок, но аналогичные опыты, поставленные на представителях других родов Grylloidea (Acheta, Gryllus, Teleogryllus, Scapteriscus), дали противоположные результаты. Причем в некоторых случаях повышение частоты призывного звука до 12-13 кГц приводило к смене положительного фонотаксиса на отрицательный. По-видимому, для устранения этих противоречий потребуются дополнительные исследования, но результаты последних экспериментов и электрофизиологические данные (см. далее) указывают на то, что у сверчков и медведок эти параметры играют существенную роль в процессе опознавания.
Для двукрылых частота, по-видимому, служит единственным критерием, по которому они могут различать звуки, возникающие в полете. Хорошо известно, например, что самцов комаров привлекают тональные сигналы, если их частота приближается к частоте биения крыльев летящей самки.
Физиологические механизмы, обеспечивающие опознавание звуковых сигналов, находятся в стадии интенсивного изучения. Накопленные к настоящему времени факты пока не позволяют составить цельного представления о работе слухового анализатора, но они отражают существенные закономерности функционирования некоторых его отделов.
Обработка акустической информации проходит в слуховых анализаторах три основных этапа. Первый из них осуществляется в слуховых органах*, второй - в интернейронах грудных ганглиев, третий - в слуховых центрах надглоточного ганглия.
* (Здесь и далее речь идет только о тимпанальных органах.)
Рецепторы отвечают на естественные сигналы синхронными залпами импульсов, передающими информацию о длительности звуковых посылок и закономерностях их повторения, т. е. с большей или меньшей четкостью отражают временной рисунок ("pattern") сигнала. Частота импульсов в разряде зависит от интенсивности звука. У прямокрылых характер ответа определяется не только временными, но и частотными параметрами сигнала. Так, у сверчков призывные и агрессивные сигналы возбуждают преимущественно низкочастотный отдел слухового органа, оптимум активности которого совпадает с основной частотой этих звуков, а прекопуляционные сигналы, содержащие ультразвуковые компоненты, воспринимаются в основном высокочастотным отделом. Следовательно, в данном случае первый этап различения внутривидовых сигналов осуществляется уже на уровне рецепторов.
Слуховые интернейроны, расположенные в грудных ганглиях, обладают разными функциональными характеристиками и в соответствии с этим по-разному отвечают на одни и те же сигналы.
Регистрируя активность восходящих волокон в шейных коннективах, можно отметить следующие основные закономерности.
Интенсивность сигнала влияет не только на частоту импульсации в разрядах, но и на число работающих нейронов, причем ее увеличение приводит либо к возрастанию, либо к подавлению активности некоторых клеток.
У прямокрылых насекомых отдельные интернейроны связаны только с высокочастотными или низкочастотными сенсиллами, поэтому информация о частоте звука может передаваться по разным каналам. Наряду с этим существуют клетки, имеющие двухвершинные характеристики, указывающие на то, что они возбуждаются обоими отделами тимпанального органа.
Быстро адаптирующиеся клетки реагируют только на начало звука или отвечают на каждый пульс одним-двумя импульсами, подчеркивая тем самым ритмическую организацию сигнала. Тонические нейроны, напротив, отражают не только частоту повторения, но и длительность звуковых посылок (рис. 18, В).
Сравнение реакций комплекса восходящих интернейронов на разные призывные сигналы свидетельствует о том, что в грудных ганглиях акустическая информация претерпевает заметные преобразования, приводящие к выделению и контрастированию многих параметров сигнала. Такая предварительная обработка, по видимому, необходима для успешного функционирования высших отделов анализатора. Однако на этом этапе еще не происходит детектирования видоспецифического призывного сигнала.
Сопоставляя далее ответы грудных интернейронов на внутривидовые сигналы, можно заметить, что каждый из них вызывает специфическую реакцию определенных клеток. Так, у сверчков на призывные сигналы отвечают нейроны С. или (при больших интенсивностях) В и С, на прекопуляционные - преимущественно В и В1, а на звуки агрессии - только В. Еще большая селективность наблюдается у некоторых кузнечиков (Isophya), издающих длительные пульсы и короткие щелчки. В грудном отделе их слухового анализатора обнаружены "детекторы пульсов" и "детекторы щелчков" - нейроны, реагирующие только на один из этих сигналов (рис. 18, А, Б).
Приведенные, а также некоторые другие факты дают основание предполагать, что в грудных ганглиях осуществляется важный этап различения и, возможно, опознавания внутривидовых сигналов.
Изучение высших отделов слухового анализатора насекомых только начинается. Судя по предварительным данным, в надглоточном ганглии не удается обнаружить детекторы, отвечающие только на видоспецифические призывные сигналы, но у саранчи все же найдены клетки, обладающие достаточно высокой избирательностью. Некоторые из них реагируют на сравнительно длинные (20 мс) низкочастотные посылки, повторяющиеся с частотой 20 в секунду. Другие возбуждаются очень короткими (0,5-5 мс) высокочастотными сигналами. Сравнение ответов этих нейронов на искусственные и естественные звуки показало, что они приспособлены для выделения стридуляционных (призывных) и мандиоулярных (возникающих при щелкании мандибул) звуков. Таким образом, в надглоточном ганглии продолжается процесс различения внутривидовых сигналов. Механизмы выделения и опознавания видоспецифических призывных сигналов пока остаются неизученными, но есть все основания предполагать, что они будут исследованы в ближайшем будущем.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'