5. Центральные отделы слуховых анализаторов
Афферентные волокна, идущие от тимпанальных органов, входят в грудные ганглии, а волокна Джонстонова органа поступают непосредственно в головной мозг (надглоточный ганглий).
Исследование функциональной организации высших отделов слуховых анализаторов проводилось до настоящего времени только на прямокрылых и чешуекрылых насекомых.
У кузнечиков в первом грудном ганглии сначала были найдены 2 крупных Т-образных нейрона, посылающих свои ветви как в надглоточный ганглий, так и во II и III грудные ганглии. Чаще всего эти клетки дают фазовые ответы, т. е. реагируют на включение или быстрое изменение амплитуды звука. Контралатеральные рецепторы оказывают на них слабое возбуждающее и сравнительно сильное тормозное влияние. Последующие эксперименты показали, что в грудном отделе имеется несколько Т-образных слуховых волокон, а также восходящие и нисходящие волокна. Среди них были найдены фазовые, тонические, фазово-тонические нейроны, а также клетки, характер ответов которых определялся условиями стимуляции. Уровень импульсной активности почти всех нейронов зависит от интенсивности и частоты звука, достигая максимума только при определенном их сочетании. На реакции большинства изученных элементов влияет также направление прихода звука, причем форма полученных диаграмм изменяется при изменении интенсивности и частоты сигнала.
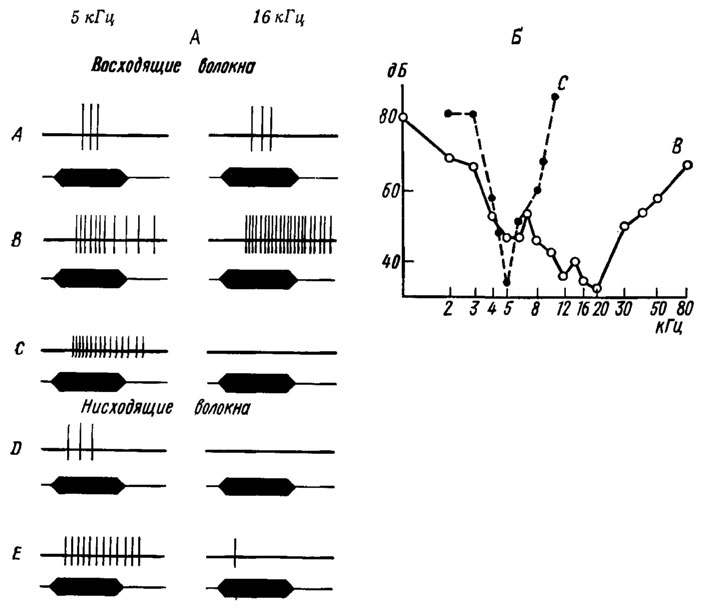
Рис. 17. Функциональные характеристики слуховых интернейронов из I грудного ганглия сверчка Gryllus bimaculatus (по Жантиеву, Чуканову, 1972). А - схема ответов 5 слуховых ни-тернейронов (А-D) на звуковые сигналы 5 и 16 кГц (длительность 50 мс, интенсивность 90 дБ). Б - частотно-пороговые характеристики двух слуховых нитерней-ронов (В и С). По оси абсцисс - частота звука, кГц; по оси ординат - звуковое давление, дБ
У сверчков в первом грудном ганглии лучше других изучены 5 центральных слуховых нейронов (рис. 17): 3 восходящих (А, В, С) и 2 нисходящих (D, Е). Все они возбуждаются низкочастотным отделом тимпанального органа (оптимум на 5 кГц), а клетки Л, В и Е, кроме того, связаны с высокочастотными сенсиллами (оптимум на 16 кГц). Вследствие этого на частотно-пороговых кривых нейронов С и D имеется только один оптимум, а у нейронов А, В и Е достаточно отчетливо выражены две зоны повышенной чувствительности. По характеру реакций на длительные сигналы эти клетки могут быть разделены на фазовые [A, D) и тонические (В, Е). При определенных условиях стимуляции почти все они испытывают тормозное влияние со стороны других нервных элементов. Контралатеральные рецепторы оказывают на некоторые нейроны только тормозное (С), а на другие - тормозное и возбуждающее воздействие (В).
Слуховые анализаторы медведок изучены еще недостаточно, но имеющиеся данные позволяют думать, что они имеют много общего с гомологичными анализаторами сверчков. В коннективах, отходящих от I грудного ганглия, у двух видов медведок удалось проанализировать ответы пяти клеток. По функциональным харак-теристикам некоторые из них близки к слуховым нейронам сверчков (А и С), но другие существенно отличаются от них. Сопоставление амплитудно-частотных спектров звуковых сигналов медведок с частотно-пороговыми характеристиками слуховых нейронов первого грудного ганглия свидетельствует о том, что этот отдел слухового анализатора, несомненно, приспособлен для восприятия и дискриминации внутривидовых сигналов.
У саранчовых, в отличие от длинноусых прямокрылых, тимпанальный нерв входит не в первый, а в третий грудной ганглий. Здесь его волокна синаптически связаны с несколькими восходящими центральными нейронами. Некоторые из них прослеживаются во всех коннективах вплоть до надглоточного ганглия. Другие достигают только проторакального ганглия. Известны также восходящие волокна, начинающиеся в мезоторакальном ганглии. Все эти нейроны хорошо различаются по основным функциональным характеристикам. Максимум их импульсной активности обычно наблюдается при определенном (оптимальном) сочетании интенсивности и частоты звука. Исключение составляет только одна клетка, которая отвечает единичными импульсами на стимулы в очень широком диапазоне частот и интенсивностей. Комплекс изученных к настоящему времени восходящих нейронов обеспечивает передачу информации об акустических сигналах в диапазоне от 3 до 40 кГц при уровнях звукового давления от 40 до 110 дБ..Приблизительно половина этих клеток испытывает влияние со стороны контралатеральных рецепторов. Кроме того, на реакции некоторых слуховых интернейронов значительное воздействие оказывают афферентные волокна, не связанные с тимпанальными органами, но реагирующие на низкочастотные колебания.
У чешуекрылых тимпанальный нерв входит в третий грудной ганглий, но его волокна прослеживаются и в других грудных ганглиях. В мезоторакальном ганглии найдены две группы восходящих нейронов второго порядка. Ипсилатеральные волокна идут отсюда в подглоточный ганглий, а контр ал атеральные достигают только проторакального ганглия. Ответы этих клеток, получивших название "релейных", мало чем отличаются от реакций афферентных нейронов. Более существенное преобразование информации производят нейроны другого типа, обнаруженные в мезоторакальном ганглии. Они отвечают на звуковые посылки любой длитель-ности только одним импульсом. В грудном отделе ЦНС чешуекрылых регистрируются ответы и некоторых других слуховых волокон, но они изучены еще недостаточно. Несмотря на сравнительную простоту организации, восходящие слуховые пути могут передавать значительный объем информации об основных параметрах звуковых сигналов (интенсивности, частоте повторения, направленности и др.).
Высшие отделы слуховых анализаторов, расположенные в надглоточном ганглии, исследованы у саранчи, кузнечиков и некоторых чешуекрылых (Noctuidae).
В первом случае нейроны, реагирующие на звук, были обнаружены в задней латеральной части протоцеребрума и в передней латеральной части дейтоцеребрума. Тщательное изучение их функциональных характеристик показало, что некоторые клетки возбуждаются сигналами, обладающими . определенным сочетанием интенсивности, частоты, длительности и частоты повторения. Иначе говоря, эти нейроны ведут себя хак фильтры, выделяющие звуки с определенными параметрами. Сопоставляя активность некоторых клеток до и после отключения одного из тимпанальных органов, удалось установить, что контр ал атеральные рецепторы оказывают на них преимущественно тормозное воздействие: На основании этого было высказано достаточно обоснованное предположение об участии нейронов надглоточного ганглия не только в распозна-вании, но и локализации звуковых сигналов.
У кузнечиков слуховые нейроны также расположены преимущественно между прото- и дейтоцеребрумом. По большинству характеристик они оказались сходными с интернейронами грудных ганглиев. Различия между ними проявляются главным образом в характере изменений активности во время стимуляции, длительности ответов и зависимости их реакций от ориентации насекомого в звуковом поле.
Исследование слухового центра в надглоточном ганглии чешуекрылых прежде всего показало, что и высший отдел слухового анализатора представлен у этих насекомых сравнительно небольшим количеством элементов. С каждой стороны протоцеребрума удалось обнаружить по одному фазовому и по два тонических нейрона. Активность одного из них подавлялась звуком больших ин-тенсивностей, что указывает на его возможную связь с низкочувствительной клеткой тимпанального органа и позволяет рассматривать этот нейрон как важный элемент механизма, обеспечивающего изменение поведения бабочек при приближении летучей мыши.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'