4. Функции слуховых органов
Как известно, приборы для регистрации звука реагируют на один из трех его параметров: звуковое давление, градиент давления или смещение частиц среды.
Прибор, регистрирующий звуковое давление, представляет собой толстостенную камеру, одна из стенок которой замещена туго натянутой мембраной. Заключенный в этом резервуаре воздух сообщается с наружной средой через небольшое отверстие, благодаря чему внутри камеры поддерживается атмосферное давление. В звуковом поле на диафрагму прибора действует сила, пропорциональная разности между давлением звука и атмосферным давлением. Если длина волны значительно превышает размеры такого приемника, то его чувствительность не зависит от направления прихода звука.
Приемники градиента давления представляют собой натянутую на жесткую рамку и открытую с двух сторон мембрану (например ленточный микрофон). Сила, действующая на мембрану в звуковом поле, зависит от угла, под которым звуковые волны падают на ее поверхность, поэтому такой приемник является направленным.
Колебания частиц воздуха можно наблюдать под микроскопом, если ввести в звуковое поле аэрозоль или какие-нибудь тела с минимальной массой (например очень тонкую кварцевую нить).
В течение длительного времени предполагалось, что все слуховые органы насекомых воспринимают смещение или градиент давления. Однако в последние годы были получены данные, свидетельствующие о том, что, по крайней мере, тимпанальные органы саранчовых и кузнечиков представляют собой сложные приемники, реагирующие как на давление, так и на градиент давления.
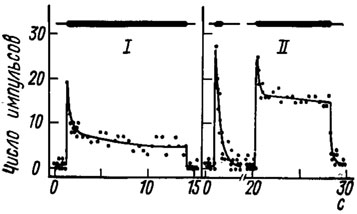
Рис. 15. Адаптация двух а-клеток из тимпанального органа саранчи Schistocerca gregaria к звуковому раздражителю (3 кГц) интенсивностью 80 дБ (I) и 90 дБ (II) (по Michelsen, 1966). По оси абсцисс - время, с; по оси ординат - число импульсов за 96 мс; толстые линии - длительность стимуляции
Анализ импульсной активности одиночных нейронов показал, что в разных слуховых органах и даже в пределах одного некоторые сенсиллы могут адаптироваться по-разному. Чаще всего через несколько миллисекунд после включения звука частота импульсов быстро возрастает до максимума, а затем более или менее быстро падает до определенной величины, после чего либо остается в течение длительного времени на этом уровне, либо постепенно снижается (рис. 15). Реже наблюдаются медленное возрастание и: быстрый спад частоты импульсации. Первые несколько импульсов: обычно генерируются в сенсиллах тимпанальных органов синхронно, в результате чего в их суммарных ответах регистрируется так называемая синхронная фаза, состоящая из нескольких волн большой амплитуды. При длительной стимуляции она сменяется асинхронной фазой, а после выключения звука наблюдается небольшое последействие.
При стимуляции Джонстоновых органов и церкальных трихоидных сенсилл низкочастотными звуками регистрируются суммарные потенциалы, представляющие собой гармонические колебания, частота которых равна или кратна частоте звука.
Абсолютная чувствительность большинства слуховых рецепторов насекомых невелика: пороги реакции, регистрируемые обычными методами, лежат в области 30-40 дБ. Использование некоторых специальных методов позволяет обнаруживать слабые ответы при УЗД, не превышающем 7-10 дБ. Максимальной чувствительностью обладают Джонстоновы органы (порог около 0 дБ).
Частота импульсации отдельных сенсилл тимпанальных органов возрастает при увеличении интенсивности стимула до 30-40 дБ над порогом реакции. Наличие в сложных слуховых органах сенсилл с разной чувствительностью значительно расширяет их динамический диапазон.
Важной особенностью тимпанальных органов насекомых следует считать чрезвычайно малую временную константу. У большинства изученных видов она составляет всего 2-3 мс. Это позволяет насекомым различать отдельные звуковые пульсы, следующие с частотой 100-300 в секунду.
При изучении функций слуховых органов насекомых большое внимание всегда уделялось их частотным характеристикам. На первых этапах этих исследований была выдвинута гипотеза, отрицавшая способность слуховых рецепторов дискриминировать звуки по частоте. Иначе говоря, считалось, что в отличие от позвоночных у насекомых все сенсиллы слухового органа "настроены" на один диапазон частот. В ходе дальнейших работ, проводившихся на представителях разных отрядов насекомых, были получены многочисленные подтверждения этой гипотезы. Однако в последние годы удалось установить, что тимпанальные органы прямокрылых имеют более сложную функциональную организацию, чем предполагалось ранее, и, в частности, содержат группы сенсилл, по-разному реагирующих на звуки разных частот.
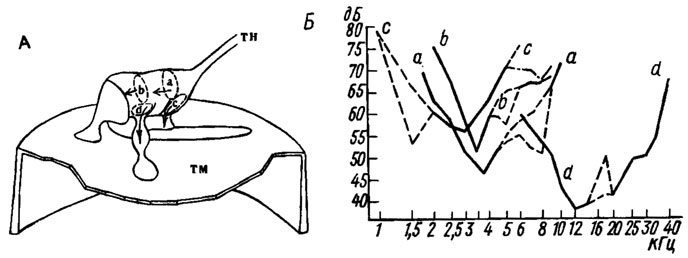
Рис. 16. Строение и функциональные характеристики тимпанального органа саранчи. А -схема строения тимпанального органа: а-d - четыре группы хордотональных сенсилл; тн - тимпанальный нерв; тм - тимпанальная мембрана (по Gray, 1960). Б - частотно-пороговые характеристики четырех групп хордотональных сенсилл в изолированном тимпанальном органе (по Michelsen, 1969). По оси абсцисс - частота звука, кГц; по оси ординат - его интенсивность, дБ
У саранчовых в изолированных тимпанальных органах выделены четыре группы сенсилл, обладающих разными функциональными свойствами (рис. 16). Три из них (а, b, с) реагируют преимущественно на звуки низких частот - от 1 до 10 кГц (оптимум чувствительности - 2-4 кГц), а одна (d) воспринимает высокочастотные колебания в диапазоне 6-40 кГц (оптимум чувствительности-12 кГц). Исследование механических свойств тимпанальной мембраны с помощью метода лазерной голографии показало, что частотные характеристики четырех групп сенсилл определяются резонансными свойствами тех специализированных участков тимпанальной мембраны, к которым они прикрепляются.
У кузнечиков и сверчков изучение тимпанальных органов затрудняется тем, что к ним примыкают сенсиллы подколенного органа, часть из которых посылает волокна в тимпанальный нерв. Тем не менее, сочетая электрофизиологические методы с перерезками веточек тимпанального нерва, удалось получить частотные характеристики этих органов. Так, было показано, что у кузнечиков подколенный орган не реагирует на звуковые колебания, превышающие 1 кГц. Промежуточный орган отвечает на звуки в диапазоне от 1 до 15 кГц (оптимум на 5-7 кГц), а слуховой гребень воспринимает высокочастотные колебания от 1 до 100 кГц (оптимум на 12-30 кГц).
Аналогичные результаты были получены при изучении функциональной организации тимпанальных органов сверчков. Низкочастотные звуки (1 -15 кГц) вызывают ответы проксимальной части трахеального органа, гомологичной промежуточному органу кузнечиков. Оптимум чувствительности этой группы сенсилл лежит в области 4-5 кГц. Дистальная часть трахеального органа воспринимает звуки более высоких частот - от 10 до 50 кГц (оптимум чувствительности- 16 кГц).
У медведок, как и у сверчков, на частотно-пороговых кривых тимпанальных органов выделяются две зоны повышенной чувствительности: на 1,5-2 кГц и на 16-18 кГц, - поэтому можно предполагать, что сенсиллы у этих насекомых также образуют два функциональных отдела, по-разному реагирующих на звуки высоких и низких частот.
В отличие от прямокрылых в тимпанальных органах цикад и чешуекрылых пока не удается обнаружить сенсилл с разными частотными характеристиками. Оптимум их чувствительности у цикад лежит в области 3-7 кГц, а у чешуекрылых он смещен в ультразвуковой диапазон- 15-60 кГц. Джонстоновы органы и трихоид-ные сенсиллы реагируют на звуки сравнительно низких частот (до 1-3 кГц).
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'