3. Физиологические особенности проприоцепторов
Проприоцепторами называют механорецепторы, посылающие в ЦНС информацию о положении, деформации и смещениях различных частей тела*. Их функционирование обеспечивает координацию всех подвижных органов и тканей насекомого в состоянии покоя и во время любых двигательных актов. При экспериментальном выключении проприоцепторов насекомые теряют способность поддерживать естественные позы, двигаться и целесообразно реагировать на внешние воздействия.
* (Некоторые проприоцепторы, воспринимающие механическое состояние внутренних органов (пищеварительной, кровеносной систем и др.), называют интеро-рецепторами.)
Функции проприоцепторов у насекомых выполняют чувствительные элементы всех описанных ранее типов (стр. 31): трихоидные, кампаниформные, хордотональные сенсиллы и рецепторы растяжения.
Трихоидные сенсиллы, выполняющие функции проприоцепторов, обычно образуют очень густые скопления (так называемые волосковые пластинки) в местах контакта подвижно сочлененных склеритов - на переднем крае груди, на щупиках, тазиках, вертлугах и т. п. Возбуждаются они при соприкосновении с межсегментной мембраной или поверхностью близлежащего склерита. Так, волоски, покрывающие передний край груди, реагируют на движения головной капсулы, а аналогичные сенсиллы тазика стимулируются при его перемещении относительно плейрита.
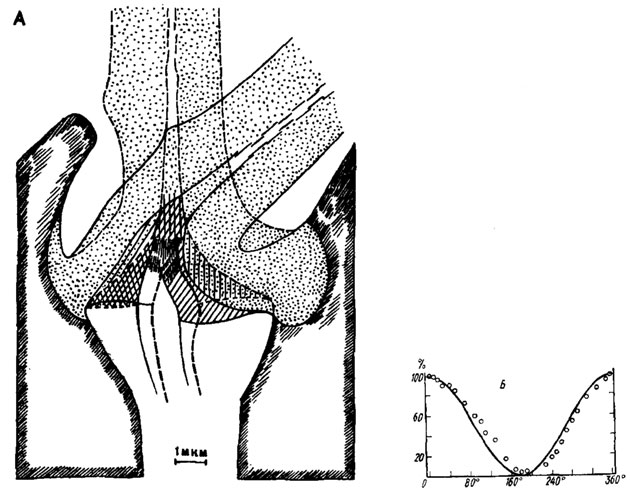
Рис. 9. Функциональные характеристики трихоидной сенсиллы пчелы Apis mellifera (noThurm, 1963, 1964). А - положение волоска в состоянии покоя (пунктир) и при его максимальном отклонении (непрерывная линия); Б - зависимость амплитуды рецепторного потенциала от направления действия стимула. По оси абсцисс - угол отклонения волоска (0° соответствует направлению, при котором наблюдается максимальный ответ); по оси ординат - относительная амплитуда рецепторного потенциала, %
Электрофизиологические эксперименты свидетельствуют о том, что уровень импульсной активности этих рецепторов пропорционален степени изгиба волосков. Асимметрия сочленовной области волосков (рис. 9, А) оказывает значительное влияние на амплитуду ответа. Зависимость реакции сенсиллы от направления отклонения волоска представлена на рис. 9, Б. Рецепторный потенциал одиночной сенсиллы после отклонения волоска быстро достигает максимума, затем падает до определенного уровня и сохраняется почти без изменений вплоть до конца стимуляции. Такой характер адаптации позволяет отнести эти сенсиллы к фазово-тоническим рецепторам.
Трихоидные сенсиллы играют очень важную роль в процессе локомоции и ориентации насекомых. Ранее (см. гл. I) были рассмотрены функции шейных волосков у богомолов. Другие примеры участия этих рецепторов в сложных двигательных реакциях дают некоторые летающие насекомые. Так, у стрекозы Апах голова обладает определенным моментом инерции, поэтому в полете при вращении туловища вокруг продольной оси она может смещаться относительно груди и возбуждать проторакальные сенсиллы. Поступающая от них информация используется для коррекции работы крыльев, компенсирующих вращательные движения.
Аналогичным образом контролируется работа ног. Многие насекомые, передвигаясь по твердому субстрату, удерживают тело на некотором расстоянии от его поверхности и сохраняют эту дистанцию даже при дополнительной нагрузке, в несколько раз превышающей их собственный вес. Однако если у них удаляют трихоидные сенсиллы в сочленении тазика и вертлуга, они полностью утрачивают способность приподниматься над субстратом.
Участие трихоидных сенсилл в механизмах, обеспечивающих ориентацию в гравитационном поле, будет рассмотрено ниже.
Кампаниформные сенсиллы рассеяны по всей поверхности тела, но, как правило, они концентрируются на тех участках кутикулы, которые испытывают более или менее значительную деформацию, в сочленениях, на придатках и т. п. местах. Подавляющее большинство исследователей считают, что эти сенсиллы реагируют на изгибы, сжатие или растяжение покровов. Анализ их ультраструктурной организации дает основание предполагать, что деформация кутикулы передается колпачку и заключенной в нем шапочке, что приводит к поперечному сжатию или растяжению вершины реснички и возбуждению нейрона.
Специальные электрофизиологические исследования показали, что вблизи сенсиллы регистрируется значительный трансэпителиальный потенциал (от +50 до +100 мВ), который возникает, по-видимому, в результате деятельности калиевых насосов тормогенной клетки, повышающих концентрацию ионов К+ в экстраклеточной полости вокруг дистальной части дендрита и его отростка. При механической стимуляции (сгибании кутикулы) происходит уменьшение величины этого потенциала, сходное с фазово-тоническим изменением рецепторного потенциала других механорецепторов. Предполагается, что амплитуда трансэпителиального потенциала контролируется сопротивлением мембраны реснички, изменяющимся при деформации ее дистальной части.
На различных участках тела насекомого наряду с одиночными круглыми сенсиллами обычно встречаются группы сенсилл с эллиптическими колпачками. Их продольные оси в пределах группы параллельны друг другу и направлению основных сил, деформирующих данный участок кутикулы. На крыльях эти сенсиллы обычно располагаются вдоль жилок, а на ногах - параллельно их продольной оси. Благодаря такой конструкции и положению эллиптические сенсиллы избирательно реагируют на механические воздействия, в результате чего несколько групп по-разному ориентированных сенсилл могут передавать информацию в ЦНС не только об интенсивности стимула, но и о его направленности. Электрофизиологические данные указывают на то, что сенсиллы одной группы функционируют как единое целое и представляют собой более или менее изолированный проприоцептивный орган.
Многочисленные наблюдения и эксперименты свидетельствуют о том, что кампаниформиые сенсиллы участвуют в регуляции разных двигательных актов. Обычно количество и степень развития этих рецепторов находятся в прямой зависимости от подвижности и маневренности насекомых. Так, например, у летающего долгоносика Sitophilus orizae на крыльях гораздо больше таких сенсилл, чем у нелетающего Sitophilus granarium.
У тараканов стимуляция кампаниформных сенсилл на вертлугах вызывает рефлекторное опускание ног. Сенсиллы, расположенные на вертлуге и в проксимальной части бедра, принимают участие в тарзальном рефлексе, т. е. запускают полет при потере контакта конечностей с субстратом.
Очень важную роль играют кампаниформиые сенсиллы в регуляции полета. Поступающая от них информация влияет на работу мышц, обеспечивающих изменение подъемной силы крыльев, как это показано на пустынной саранче. При выжигании этих рецепторов на крыльях существенно нарушается способность насекомого маневрировать в полете. Изучение импульсной активности сенсилл на задних крыльях саранчи показало, что при периодической деформации крыла ответ сенсилл синхронизируется с ритмом раздражения вплоть до частоты 65-70 Гц, причем уровень реакции зависит от ускорения деформации.
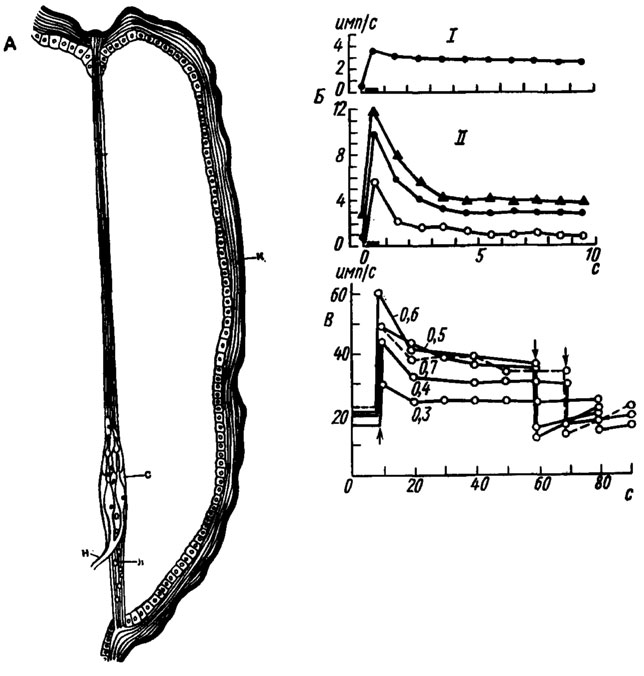
Рис. 10. Строение и функции проприоцепторов. А - схема строения туловищного хордотонального органа личинки жука-усача Ergates (no Hess, 1917): к - кутикула; л - лига-мент; н - нерв; с - сенсиллы. Б - адаптация тонических (I) и фазово-тонических (II) хордото-нальиых сенсилл в тибиотарзальном сочленении таракана Periplaneta americana (no Young, 1970). По оси абсцисс - время, с; по оси ординат - частота импульсов, имп/с. Длительность стимуляции (отклонение лапки) показана линией над осью абсцисс. Три кривых на графике II получены при последовательном увеличении угла отклонения лапки. В - зависимость частоты импульсации рецептора растяжения гусеницы Antheraea pernyi от интенсивности стимула (по Finlayson, Lowenstein, 1958). По оси абсцисс - время, с; по оси ординат - частота импульсов, имп/с. Стрелками показаны начало и конец стимуляции. Цифры под кривыми - удлинение рецептора, мм
Хордотональные органы состоят из нескольких (как исключение, одиночных) сенсилл, натянутых между подвижно сочлененными сегментами или частями тела (рис. 10, А). Их шапочковые клетки обычно прикрепляются к кутикуле или сочленовным мембранам, а проксимальные отделы фиксируются лигаментами или прилежащими тканями.
Множество таких органов найдено в антеннах, ротовых придатках, в ногах, у основания крыльев, в груди, в брюшке и других частях тела. Их общее число у одного насекомого может достигать нескольких десятков: у личинок Drosophila, например, обнаружено 90 таких образований. Обычно хордотональные органы состоят из однотипных сенсилл, но в некоторых из них встречаются сенсиллы с одним и двумя нейронами (см. стр. 34). В настоящее время можно считать доказанным, что эти органы фукционируют как проприоцепторы, воспринимающие смещение между органами и тканями под действием внешних и внутренних сил. Механизм работы отдельных сенсилл не вполне ясен, но большинство исследователей считает, что возбуждение их нейронов происходит при деформации сенсилл, приводящей к взаимодействию шапочки и дистальной (возможно, и расширенной) части реснички. Не исключена возможность, что характер этого взаимодействия различен в разных органах.
Электрофизиологические эксперименты показали, что у ракообразных и насекомых в ножных хордотональных органах имеются мелкие тонические и крупные фазовые (или фазово-тонические) элементы (рис. 10, Б). Следовательно, в пределах одного органа существует определенная функциональная специализация сенсилл. Одни из них передают информацию преимущественно о положении сочлененных участков ноги, а другие - об их движении.
Тщательное морфологическое и физиологическое исследование тибиотарзальных хордотональных органов у таракана позволило получить некоторые представления о механизме их работы. В частности, было установлено, что они реагируют на опускание и отведение лапки назад, но не проявляют активности при ее возвращении в исходное положение. Учитывая конструктивные особенности сочленения, можно с достаточной уверенностью предполагать, что возбуждение сенсилл происходит при их растяжении, а не при расслаблении или сжатии. На основании структурных различий сенсиллы, входящие в состав этого органа, были разделены на три группы. Одну из них, состоящую из сравнительно крупных, проксимально расположенных сенсилл, удалось идентифицировать с фазово-тоническими элементами, а другую, дистально лежащую группу мелких сенсилл - с тоническими.
Один из наиболее сложных хордотональных органов - Джонстонов орган, помещающийся во втором членике антенн. После первых морфологических работ стало очевидным, что его радиально расположенные сенсиллы должны реагировать на смещение третьего членика (и связанного с ним жгутика) относительно второго. Последующие физиологические исследования подтвердили это предположение. Стимуляция Джонстонова органа происходит как при активном ощупывании антеннами различных предметов, так и во время движения насекомого в воздушной или водной среде. Доказано также, что этот орган воспринимает звуковые колебания и силу тяжести.
Электрофизиологические исследования показали, что у мух Caltiphora отклонение аристы и ее возвращение в исходное положение вызывают появление в электроантеннограмме двух значительных по амплитуде волн (до +5 мВ), возникающих, по-видимому, в результате суммации ответов множества синхронно реагирующих сенсилл Джонстонова органа. Амплитуда первой волны зависит от интенсивности стимула. Анализ электрической активности дает основание предполагать, что некоторые сенсиллы отвечают на любые колебания аристы, в то время как другие возбуждаются только при определенном направлении стимула. Подобная избирательность была обнаружена и при микроэлектродной регистрации ответов одиночных сенсилл в Джонстоновом органе саранчи. Помимо фазовых реакций Джонстонова органа в антеннах наблюдаются и тонические ответы каких-то иных рецепторов (возможно, трихоидных сенсилл), передающих информацию о положении жгутика относительно второго членика.
Рецепторы растяжения, несмотря на их морфологическое разнообразие, обладают сходными физиологическими характеристиками. В состоянии покоя (при минимальном натяжении) спонтанная импульсная активность обычно поддерживается на уровне 5-10 имп/с. При растяжении рецептора импульсацчя быстро возрастает до максимума, а затем снижается до определенного уровня, на котором она может оставаться в течение длительного времени (рис. 10,В). В момент расслабления импульсная активность резко падает (иногда до 0), после чего наблюдается постепенное восстановление исходного уровня импульсации. Благодаря такой фазово-тонической реакции рецептор растяжения передает информацию как о переходных процессах, так и о стационарных смещениях близлежащих структур. Частота импульсов в начальной фазе разряда возрастает пропорционально увеличению интенсивности стимула, но при экстремальном растяжении рецептора эта зависимость нарушается. На частоту импульсации оказывает влияние также скорость, с которой изменяется длина соединительнотканного тяжа или мышцы. Так, в рецепторах, лежащих у основания крыльев саранчи, частота разрядов и темп ее нарастания максимальны при быстрых перемещениях крыла. Во время ритмического качания крыловой пластинки рецептор отвечает синхронными залпами импульсов только на ее подъем, причем увеличение частоты колебаний приводит к уменьшению числа импульсов в разрядах. Специальные эксперименты показали, что удаление рецепторов растяжения или искусственная стимуляция идущих от них аксонов оказывают существенное влияние на ритм работы крылового аппарата саранчи. Анализ полученных результатов свидетельствует о том, что эти органы выполняют функцию обратной связи, стабилизируя уровень активности центральных нейронов, управляющих работой крыловых мышц.
По-видимому, не менее важную роль рецепторы растяжения играют в регуляции других локомоторных реакций, и в частности червеобразных движений личинок. Кроме того, есть основания думать, что они участвуют в контроле ритмических движений брюшка в процессе дыхания. Известно также, что у клопа Rhodnius растяжение абдоминальных рецепторов тормозит кровососание и стимулирует выделение гормона линьки.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'