2. Функции тактильных рецепторов
При соприкосновении насекомых с твердым субстратом или при их перемещении в воздушной или водной среде возбуждаются механорецепторы, но наиболее важную роль в восприятии контактных стимулов играют рассеянные по всему телу трихоидные сенсиллы. Обычно они концентрируются на тех склеритах и придатках, которые чаще всего соприкасаются с окружающими предметами или интенсивней обдуваются во время полета.
Морфологические данные свидетельствуют о том, что, несмотря на сравнительно простую организацию и однотипность строения, эти сенсиллы часто обладают признаками достаточно выраженной специализации, указывающими на возможность избирательной рецепции определенных механических стимулов. Результаты физиологических и этологических экспериментов позволяют разделить трихоидные сенсиллы наземных насекомых на две группы. Представители одной из них возобуждаются при контакте с твердыми телами и могут быть названы собственно осязательными, в то время как другие приспособлены для восприятия движения воздуха и получили название ветрочувствительных. У водных форм аналогичные рецепторы должны возбуждаться во время плавания, но они изучены еще недостаточно, поэтому мы ограничимся рассмотрением двух упомянутых выше групп.
Детали строения осязательных рецепторов варьируют в значительных пределах, но почти все сенсиллы этого типа обладают сравнительно жесткими толстостенными волосками и мало-эластичными сочленовными мембранами. Ломимо простых механо-рецепторов у насекомых встречаются сенсиллы со смешанными функциями, реагирующие на механические и химические стимулы. В полость их волосков входят отростки хеморецепторных нейронов (см. стр. 83).
Согласно существующим представлениям, при отклонении волоска из положения равновесия его основание или сочленовная мембрана деформируют шапочку и заключенную в ней вершину реснички, что и вызывает возбуждение нейрона (см. рис. 9,А).
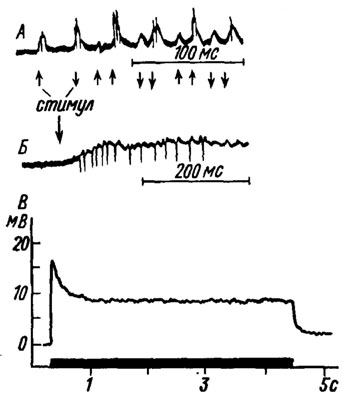
Рис. 8. Ответы трихоидных сенсилл на механические стимулы: А - расположенных на крыльях мухи Sarcophaga sp. (no Wolbarsht, 1960); Б - расположенных на антеннах саранчового Melanoplus femur rubrum (no Wolbarsht, I960); В - ветрочувствительиых сенсилл саранчи Locusta migratoria (изменение рецепторного потенциала (по Smola, 1970). По оси абсцисс - время, с; по оси ординат - амплитуда рецепторного потенциала, мВ. Продолжительность стимуляции (отклонение волоска) показана линией над осью абсцисс
Исследование трихоидных сенсилл у двукрылых и прямокрылых показало, что сгибание волоска вызывает появление отрицательного рецепторного потенциала, амплитуда которого достигает нескольких милливольт (рис. 8, А, Б). По характеру реакции эти сенсиллы были разделены на фазовые и тонические. У первых потенциал возникает только во время движения волоска (рис. 8,А), а у вторых он регистрируется в течение всего периода стимуляции, т. е. отклонения волоска из положения равновесия (рис.8,Б). Если рецепторный потенциал достигает пороговой величины, нейрон начинает генерировать импульсы положительной полярности. Их амплитуда находится в прямой зависимости от величины рецепторного потенциала. Частота импульсации также увеличивается с ростом амплитуды этого потенциала, но прямая зависимость между ними сохраняется только в определенных пределах.
Для фазовых рецепторов характерна быстрая и полная адаптация, а интенсивность реакции зависит не только от амплитуды, но и скорости изменения стимула. Тонические сенсиллы, напротив, адаптируются очень медленно, и их импульсная активность обычно не снижается до нуля даже при очень длительной стимуляции. Эти различия дают основание предполагать, что сенсиллы первого типа играют существенную роль при кратковременных соприкосновениях с субстратом - при движении, ощупывании и т. п., а тонические элементы функционируют главным образом во время длительных контактов - при копуляции, яйцекладке и др.
Используя информацию от рассеянных по всему телу трихоидных сенсилл, насекомые получают представление о субстрате и окружающих предметах практически при всех видах жизнедеятельности. Без тактильной рецепции невозможны ни простые двигательные акты, ни сложные формы поведения. Особенно велико значение осязания у насекомых, активных в ночное время или живущих в темноте - в норах, скважинах, пещерах, в толще субстрата и других подобных местах.
Ветрочувствительные сенсиллы в отличие от осязательных обладают более тонкими волосками и чрезвычайно эластичной сочленовной мембраной, обеспечивающей исключительно высокую чувствительность этих рецепторов к перемещениям окружающего воздуха. Такие сенсиллы встречаются на многих частях тела насекомых. Лучше других в настоящее время изучены рецепторы, расположенные на голове у саранчи и на церках многих ортоптероидных насекомых, поэтому, говоря о ветрочувствительных сенсиллах, мы будем иметь в виду прежде всего эти рецепторы.
Морфологические исследования показали, что основание волоска заканчивается двумя небольшими отростками, которые входят в соответствующие ямки в кутикуле, в результате чего образуется сочленение, облегчающее качание волоска в определенной плоскости. Отклонение волоска в одном из двух возможных направлений вызывает деполяризацию мембраны нейрона и появление медленного рецепторного потенциала. Его амплитуда в течение нескольких миллисекунд достигает максимума, а затем (если волосок не возвращается в исходное положение) падает до определенного уровня, на котором остается до конца стимуляции (рис. 8, Б). Такой фазово-тонический характер изменения потенциала позволяет сенсилле реагировать как на колебания, так и на стационарные отклонения волоска. У каждой сенсиллы деполяризация мембраны наступает только при смещении волоска в определенном направлении. Отклонение его в противоположную сторону (от положения равновесия) вызывает гиперполяризацию. Эти различия можно объяснить тем, что вершина отростка дендрита не совпадает с продольной осью волоска или (в случае церкальных сенсилл) окружена кутикулярными складками только с трех сторон, поэтому качание волоска оказывает на отросток неэквивалентное воздействие. По-видимому, одно из движений приводит к сжатию, а противоположное - к растяжению его дистальной части. При увеличении угла отклонения волоска до 12-25° происходит возрастание амплитуды как фазового, так и тонического компонентов рецепторного потенциала, но в первом случае наблюдается логарифмическая, а во втором - линейная зависимость. Эти закономерности нарушаются, если волосок отклоняют за пределы, ограниченные сочленовным валиком.
При достижении пороговой величины рецепторный потенциал вызывает появление в аксоне импульсных разрядов. Их частота пропорциональна амплитуде потенциала в области плато. Интенсивность ответа рецептора зависит не только от величины силы, отклоняющей волосок, но и от скорости его смещения. Так, в сенсиллах на голове саранчи максимальная частота импульсации и наибольшая скорость ее нарастания наблюдаются при очень быстром отклонении волоска, если же смещение волоска замедлено, то импульсация нарастает постепенно, и фазовая часть ответа может вообще отсутствовать.
Хотя каждая сенсилла обладает направленной чувствительностью, ее возможности в этом отношении очень ограничены. Сравнительные данные свидетельствуют о том, что плоскости качания близлежащих сенсилл ориентированы по разному, поэтому при изменении направления стимула возбуждаются разные группы рецепторов. Совокупность таких элементов, очевидно, представляет собой рецептивное поле, передающее информацию о направлении потока воздуха, воздействующего на данный участок кутикулы.
Тщательные исследования трихоидных сенсилл на голове саранчи показали, что эти рецепторы участвуют в запуске и поддержании полета. Аксоны их биполярных нейронов входят в головные ганглии, где они синаптически связаны с нисходящими фазовыми и тоническими (фазово-тоническими) интернейронами, влияющими на работу нейронов крылового аппарата.
Возбуждение трихоидных сенсилл на церках тараканов и сверчков, происходящее при обдувании насекомого или быстром приближении к нему какого-нибудь предмета, обычно вызывает реакцию убегания. В течение длительного времени считалось, что афферентные импульсы возбуждают в последнем брюшном ганглии несколько гигантских нейронов, идущих через все брюшко в метаторакальный ганглий, где они запускают реакцию мотонейронов третьей пары ног. Новейшие исследования показали, однако, что мотонейроны связаны синаптически не с гигантскими волокнами, а с идущими параллельно небольшими нейронами, обладающими меньшей скоростью проведения. Функции гигантских волокон, продолжающихся до надглоточного ганглия, пока остаются не совсем ясными.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'