2. Особенности сенсорной системы
Сенсорные органы представляют собой специализированные образования, предназначенные улавливать энергию внешних или внутренних стимулов для активации их нервных клеток, чьи сигналы информируют ЦНС о характере стимуляции. К таким стимулам, посредством которых организм регулирует свои отношения с окружающей средой, относится прежде всего энергия частиц и молекул, слагающих мир. Так, зрительные клетки поглощают фотоны определенной энергии, отраженные окружающими предметами или исходящие непосредственно от их источника. Лучистое тепло улавливают другие клетки путем абсорбции фотонов более широкого энергетического уровня. Вкус и запах улавливаются путем утилизации потенциальной энергии, заложенной во взаимном притягивании и отталкивании частиц, образующих атом. Звук обнаруживается посредством использования энергии движения частиц моле кулярного уровня.
Все клетки организма работают за счет внешней энергии, получаемой в конечном счете в виде молекул или частиц. Но чувствующие клетки отличаются от остальных, по крайней мере, в двух отношениях. Во-первых, энергия внешнего воздействия служит для чувствующей клетки только стимулом, или триггером, для запуска процессов, совершаемых за счет потенциальной, накопленной через механизм обмена энергии самой клетки. Поэтому энергия, необходимая для возбуждения рецептора, всегда меньше той, которую несет его выходной сигнал. Во-вторых, чувствующая клетка имеет на выходе электрическую энергию, обязательно передаваемую другим клеткам, которые сами не способны воспринимать энергию данного внешнего воздействия.
Обратимость изменения состояния в ответ на внешнее воздействие позволяет чувствующей клетке служить датчиком информации, однако в этой роли к ней предъявляются очень жесткие требования. Рецептор не сможет дать много полезной организму информации, если он так груб, что не чувствителен даже к большим изменениям энергии на входе, или, наоборот, так "совершенен", что реагирует на незначительные флюктуации входного сигнала. Кроме того, польза рецептора была бы небольшой при одинаковой реакции его на разные виды энергии или простой сигнализации о наличии или отсутствии стимула. С точки зрения запросов организма у рецептора должны быть:
- некоторый оптимум чувствительности;
- способность выделять определенные виды стимулов;
- способность улавливать не только появление и исчезновение стимула, но и величину, направление и скорость его изменения.
Природа позаботилась об удовлетворении этих запросов. Хотя для клеточной протоплазмы характерна умеренная чувствительность ко многим видам энергии и реакция на многие параметры стимулов, рецепторы специализировались на восприятие определенного вида энергии. Специализация рецептора выражается в повышенной чувствительности каждого к "своему" виду стимула и его определенным параметрам. Это оказалось возможным, например, за счет приобретения рецепторными клетками специальных свойств на молекулярном уровне. Так, приобретение зрительными клетками фотолабильного пигмента позволило поглощать лучистую энергию в определенном диапазоне длин волн и отличать свет от теплового воздействия лучей.
В дополнение к специализированной чувствительности как таковой рецепторные клетки приобрели окружение в виде вспомогательных структур, предопределяющих род воспринимаемой энергии. В простейшем случае это - изгибающийся под действием внешних сил кутикулярный волосок, в более сложном - хрусталик, ирисовые клетки и другие элементы глаза, обслуживающие фоторецепторную часть - сетчатку, или тимпанальная мембрана органа слуха. Названные структуры могут не только влиять на чувствительность такой комбинированной рецепторной сенсиллы (например удаление хрусталика ухудшает зрение, а прободение тимпанальной мембраны ухудшает слух), но и предопределяют, какие параметры стимула (величина, скорость изменения и т. п.) могут быть использованы. Можно сказать вслед за известным американским физиологом В. Дж. Детье, что "очищение" рецепторных органов от генерализованных признаков и неизбирательности ответа прослеживается на всем эволюционном пути как их сенсорных клеток, так и дополнительных структур.
В чем же специфика сенсорной системы насекомых? Два типичнейших признака насекомых - малая величина тела и наличие экзоскелета - накладывают на их сенсорную систему и весь нервный аппарат определенные ограничения. Прослеживаемая в филогенезе насекомых миниатюризация тела ведет к укорочению нервных путей и сокращению времени поступления нервных импульсов от рецептора к нервному центру и от последнего к эффектору. Это может повлечь за собой, при прочих равных условиях, более высокую реактивность и подвижность малого по размерам организма по сравнению с крупным. И действительно, среди класса насекомых у мелких форм тараканов, прямокрылых, жуков, перепончатокрылых замечена тенденция реагировать быстрее по сравнению с крупными формами. Кроме того, малая величина тела неизбежно вызывает уменьшение числа нейронов, возможных в данном объеме. Если мозг млекопитающих содержит более 1010 нейронов, то во всей нервной системе таракана Periplaneta число нейронов - порядка 105, а в каждом отдельном ганглии его брюшной цепочки - от 102 До 103 нейронов. При небольшом числе нейронов уменьшается информационная емкость нервной системы, но это обстоятельство, как мы увидим ниже, не ограничивает процветания насекомых. Более того, редукции числа нейронов у них способствует образование специальных гигантских волокон, так характерных для этих членистоногих. Например, гигантские волокна у Реriplaneta занимают 12% площади поперечного сечения брюшной нервной цепочки.
По мнению физиолога Редера (1959), относительное преимущество нервной системы из малого числа крупных единиц перед нервной системой, образованной многими небольшими единицами, легко понять, если допустить, что скорость реакции для ее обладателя важнее, чем детали информации. Так, чтобы обнаружить и успеть избежать врага, высокая скорость реагирования дает насекомому большую выгоду, чем подробная информация о враге. Для насекомого-хищника скорость броска также важнее подробного представления о жертве. С этой точки зрения вполне понятно развитие системы гигантских волокон, проводящих нервные команды быстрее, хотя бы и ценой потери потенциальной информации, которую мог бы передать нерв того же сечения, но составленный из большого числа обычных (тонких) волокон.
Пример экономности нервных элементов мы находим и в моторной системе насекомых. Мышцы насекомых иннервируются не сотнями двигательных волокон, а единицами: их может быть четыре, два и даже одно на целую мышцу. Всю мускулатуру второй пары ног личинки стрекозы Aeschna обслуживают только 6 мотонейронов. У пустынной саранчи такую сложную мышечную систему, как крыловая, обслуживают не более чем по 80 мотонейронов в мезо- и метаторакальном ганглиях. Однако миниатюрный двигательный аппарат насекомых работает не менее прецизионно, чем даже у позвоночных, а по скорости сокращения мышц насекомые намного превосходят их.
Очевидно и ограничение числа нейронов в собственно сенсорной системе насекомых. Оно связано не только с миниатюрностью тела, но и с тем, что тело заключено в жесткую, лишенную чувствительности кутикулу. Если у млекопитающих с их мягкими покровами кожные рецепторы исчисляются миллионами, то покров насекомых относительно малочувствителен, поскольку энергия внешних стимулов может достичь чувствующих клеток только в отдельных точках тела, специально предусмотренных природой. Например, на вентральной поверхности каждого брюшного кольца личинки четвертого возраста клопа Rhodnius находится до 420 рецепторов; на всей ноге мухи Phormia - менее 500 рецепторов; общее число тактильных рецепторов на теле трутня медоносной пчелы не превышает 3000. И только на высокоспециализированных сенсорных областях - в глазах и на антеннах - число рецепторов велико, хотя и здесь их все же меньше, чем в органах зрения и обоняния позвоночных. В фасеточном глазу максимальное число рецепторных (зрительных) клеток обнаружено у стрекоз - приблизительно до 210 000. Обычно же их всего несколько тысяч или даже сотен, тогда как в сетчатке позвоночных - миллионы. На каждой антенне трутня медоносной пчелы до 500 тыс. сенсорных клеток.
С экономностью числа нервных клеток связана интересная особенность нервной системы насекомых, заключающаяся в том, что для совершения рефлекторного ответа организма достаточно возбудить одну-единственную чувствующую клетку. Так, стимуляция одного чувствующего нейрона волоска на оральном диске мух вызывает движение хоботка, т. е. запускает один из начальных этапов пищевой реакции. Сходным образом, раздражение одного тактильного нейрона может вызвать в ряде случаев последовательность поведенческих ответов - от простого отдергивания придатка до убегания насекомого.
Экономность сенсорной системы насекомых выражается, наконец, и в том, что, по-видимому, подавляющее большинство их рецепторных клеток, независимо от модальности или расположения на теле, - первичночувствующие. Так называемые "первичные" (первичночувствующие) рецепторные клетки выполняют двойную функцию: они не только воспринимают энергию внешнего стимулу, но и генерируют и сами же передают импульсы в ЦНС. "Вторичными" (вторичночувствующими) рецепторами называют такие, у которых чувствующий нейрон возбуждается не внешним стимулом, а через посредство неразрывно связанной с ним одной или нескольких собственно воспринимающих клеток. К вторичным рецепторам у высокоразвитых животных относят рецепторы органов слуха, зрения, вкуса, боковой линии, а к первичным - обонятельные и тактильные клетки, мышечные веретена.
Чувствующий нейрон (или группа нейронов) у насекомых, как правило, объединен с дополнительными обслуживающими клетками в элементарный орган, называемый сенсиллой. Как нейрон, так и функционально связанные с ним клетки, дающие выход к кутикулярным структурам или внутренним стенкам тела, происходят путем деления одной материнской эктодермальной клетки. Полагают, что за возможным исключением фоторецепторов все сенсиллы, столь характерные насекомым, представляют собой гомологи кожного волоска, или хеты. Они состоят минимум из трех клеток: волоскообразующей (трихогенной), гнездообразующей (тормогенной) и чувствующего нейрона. Сенсиллы рассеяны по телу поодиночке или образуют поля, либо объединены в сложные органы, такие, как, например, фасеточные глаза и тимпанальные (слуховые) органы.
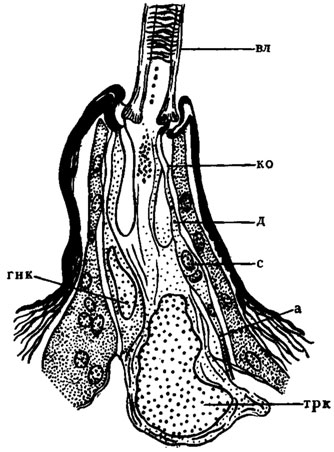
Рис. 3. Схема осязательного волоска гусеницы бабочки-крапивницы Vanessa urtiсае (по Hsu, 1938): а - аксон чувствующей клетки; вл - волосок; гнк - гнездообразующая клетка; д - дендрит чувствующей клетки; кокутикулярная оболочка; с - сома чувствующей клетки (биполярного нейрона); трк - трихогенная клетка
Общий план строения сенсилл рассмотрим на примере наиболее простой - трихоидной - сенсиллы, или кожного волоска (рис. 3). Его функция полимодальна: осязание и восприятие колебания воздуха. Полый кутикулярный волосок (вл) сидит в гибкой мембранозной впадине. С утолщенным основанием волоска контактирует вершина короткого неразветвленного дендрита (д) чувствующей клетки, одетого в своей терминальной части кутикулярным футляром (ко)*. Длинный аксон (а) на противоположном конце чувствующей клетки связывает ее с соответствующим сегментальным ганглием. Волосок настолько упруг, что всякое прилагаемое к нему усилие передается впадине. В результате деформируется вершина сколопоидного футляра и в дендрите возникает рецепторный потенциал.
* (Разные авторы по-разному называют кутикулярный футляр: сколопоидная оболочка, штифт, сколопс, кутикулярная шапочка. Последнее название не следует путать с резилиновой шапочкой, в которую входит в механорецепторных сенсиллах вершина кутикулярного футляра (см. гл. II).)
Простейший кожный волосок имеет только один нейрон, но бывают сенсиллы с несколькими (даже до нескольких десятков) чувствующими клетками. Кроме того, в состав сенсиллы часто входят обкладочные и другие клетки, назначение которых будет разъяснено ниже.
В зависимости от формы и положения волоска или его производного различают базиконические, целоконические, плакоидные и многие другие типы сенсилл (см. гл. гл. II и III). Единой же морфофункциональной классификации сенсилл не существует, и эта задача едва ли выполнима на современном уровне знания их тонкой организации. Дело в том, что при сходстве формы внешних кутикулярных придатков (а именно по их виду именуют сенсиллы) функция сенсилл может и совпадать и резко отличаться, а среди морфологически непохожих сенсилл встречаются такие, которые выполняют одинаковые функции. Например, волосковидные сенсиллы могут быть у разных видов насекомых или на разных участках тела того же индивида механо-, гигро-, термо- либо даже хеморецепторами. Если же это механорецепторный волосок, то в зависимости от положения на теле или видовой принадлежности он будет улавливать ветер (встречный поток воздуха), прикосновение к телу, его сотрясения либо даже звук в воздухе. Элементарными обонятельными органами могут оказаться базиконические и плакоидные, целоконические и некоторые другие сенсиллы. Даже такие специализированные сенсиллы, как хордотональные, реагируют на звук, вибрацию, растяжение частей тела. Наконец, известны и такие сенсиллы, которые одновременно служат двум целям. Например, волоски на хоботке мухи Calliphora - одновременно механо- и хеморецепторы. Одни чувствующие клетки таких сложных сенсилл возбуждаются при сдавливании, а другие - исключительно химическими стимулами.
Представление о многообразии видов чувствительности насекомых дает следующая классификация, в которой, повторяем, пока только отдельным модальностям стимулов удается указать морфологически строго определенный тип сенсиллы, например восприятию фотонов света - светочувствительную клетку с рабдомом, содержащим фотопигмент.

Помимо этого насекомые реагируют также на силу гравитации, на магнитные и электрические поля, но специфических рецепторов, предназначенных исключительно для восприятия названных стимулов, у них пока не обнаружено. Реакция обеспечивается прежде всего возбуждением разнообразных механорецепторов, локализованных в местах сочленений тела: головы с грудью, переднегруди со среднегрудью и, конечно, отдельных частей ног. Что касается чувствительности к магнитному полю и умеренному по силе полю статического электричества, то скорее всего и она связана с возбуждением проприорецепторов, контролирующих растяжение и взаимное положение частей тела.
Таким образом, большинство типов рецепторов насекомых "обращено" во внешнюю среду. Это - экстерорецепторы, воспринимающие внешние стимулы изначально контактным способом, а по мере специализации - и на расстоянии, т. е. дистантно. Но столь же важны для жизни насекомых интеро- и проприорецепторы, сигнализирующие о раздражителях внутренней среды и двигательной системы организма. Интеро- и проприорецепторы, хотя и "обращены внутрь" организма, но при определенных условиях, как, например, в случае восприятия гравитации или магнитного поля, могут давать информацию и о внешней среде. В последнем случае ЦНС отличает (опознает) реакцию, например, проприорецепторов при активном сгибании суставов от реакции на гравитацию. Совпадение сигналов от рецепторов в разных частях тела истолковывается как действие силы тяжести, а несовпадение - как следствие локомоции.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'