Глава I. Общая характеристика нервной системы и органов чувств насекомых
1. Краткие сведения об анатомии и гистологии нервной системы
По особенностям организации и функции нервная система насекомых может быть подразделена на центральную, симпатическую и периферическую.
Центральная нервная система имеет метамерное строение: она состоит из серии нервных узлов, или ганглиев, соединенных в нервную цепочку межсегментными стволами - коннективами. Каждый ганглий парный, его левая и правая половины соединены более или менее выраженными комиссурами. Передний отдел нервной цепочки из надглоточного и подглоточного ганглиев называют головным мозгом, а остальные ганглии - брюшной нервной цепочкой. Число ганглиев в брюшной цепочке у разных филогенетических групп насекомых и даже разных стадий их индивидуального развития различно, однако степень интеграции ганглиев, как правило, не коррелирует с уровнем поведения насекомого.
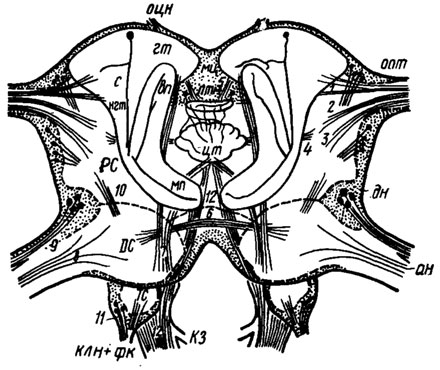
Рис. 1. Схематизированная картина фронтального среза через надглоточный ганглий сверчка Gryllus campestris(по Huber, 1960). Границы нейропиля прото-(PC), дейто-(ДС) и тритоцеребрума (ТС) обведены прерывистой линией. Ассоциативные ядра протоцеребрума обведены сплошными линиями. ан - антеннальный нерв; вп - возвратный придаток ножки грибовидного тела: гт - грибовидное тело; дн - двигательные нейроны дейтоцеребрума; клн+фк - клинео-лабральный нерв и фронтальный коннектив; кЗ - комиссура тритоцере-брума; мп - медиальный придаток ножки грибовидного тела; мц - межцеребральная часть; нгт - ножка грибовидного тела; опт - оптические доли; оцн - оцеллярный нерв; цт - центральное тело; птм - протоцеребральный мост. Нервные пути: 1-3 - оптический тракт; 4 - оптическая комиссура; 5 - оцеллярный тракт; 6 - комиссура дейтоцеребрума; 7- ольфакторно-глобулярный тракт; 8 - афферентные нервы антенн; 9 - эфферентные аксоны, идущие к мышцам антенн; 10 - связь между прото- и дейтоцеребрумом; 11 - моторные аксоны, идущие к верхней губе; 12-нервные пути, связывающие надглоточный ганглий с подглоточным
Надглоточный ганглий - самый крупный и наиболее сложный нервный узел, сходный по функциям с головным мозгом позвоночных. Его образуют, по крайней мере, три слившихся сегментальных ганглия: прото-, дейто- и тритоцеребрум. Более крупный протоцеребрум (рис. 1, PC) тесно связан с фасеточными глазами (или стеммами у личинок Holometabola) и дорсальными глазками, а его грибовидные тела (гт) и примыкающие к ним мозговые ядра служат высшим ассоциативным центром, где замыкаются условные связи. Грибовидные тела наиболее развиты у общественных насекомых, ведущих, в сравнении с остальными, более сложный образ жизни и обладающих высокой способностью к научению. Двустороннее разрушение грибовидных тел влечет выпадение приобретенных навыков, но безусловные реакции на те же стимулы у насекомого сохраняются. Дейтоцеребрум (DC) связан с антеннами, их сенсорным аппаратом и мышцами, поэтому считается обонятельным центром. Миниатюрный тритоцеребрум (ТС) иннервирует верхнюю губу, а также связывает надглоточный ганглий с симпатической нервной системой.
Подглоточный ганглий, также состоящий из трех сегментальных нервных масс, обслуживает сенсиллы и мышцы ротовых частей - соответственно мандибул, максилл и лабиума, а также рецепторы и эффекторы шейной области.
Брюшная нервная цепочка начинается тремя грудными ганглиями, которые у одних насекомых лежат раздельно, у других - сливаются вместе (иногда только мезо- и метаторакальный ганглии) или даже объединяются с несколькими (и изредка со всеми) брюшными ганглиями. Грудные ганглии ответственны в первую очередь за управление движениями ног и крыльев. В них могут также замыкаться условные локомоторные рефлексы.
Менее сложно устроены брюшные ганглии, число которых не превышает 8, иннервируют мышцы брюшных сегментов и их придатков и получают от них сенсорные нервы. Представление о том, в каких комбинациях могут сливаться ганглии брюшной нервной цепочки, дает рис. 2.
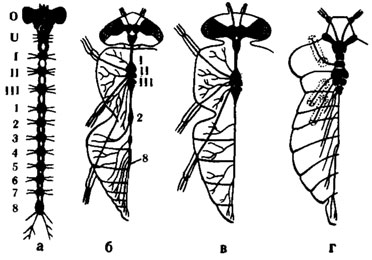
Рис. 2. Различные ступени концентрации центральной нервной системы насекомых(по Eidmann, Kuhlhorn, 1970): а - гипотетическое исходное состояние; б - слепень Tabanus bovinus; в - муха Sarcophaga; г - клоп семейства Pentatomidae. О - надглоточный ганглий; U - подглоточный ганглий; I-III - грудные ганглии; 1-8 - брюшные ганглии
Симпатической, или вегетативной, нервной системы, регулирующей работу внутренних органов, и связанного с ней нейросекреторного аппарата насекомых мы практически не будем касаться в дальнейшем изложении.
Периферическая нервная система состоит из афферентных и эфферентных нервов, чувствующих клеток со вспомогательным аппаратом и эфферентных окончаний. Наше последующее описание будет посвящено в основном именно этой части нервной системы.
Основная структурная единица нервной системы насекомых, как и всех высокоорганизованных животных, - нервная клетка, или нейрон. От тела нейрона, в котором заключено ядро, отходят обычно один длинный неразветвленный аксон и несколько коротких ветвящихся дендритов. В зависимости от числа и взаимного расположения этих отростков говорят о моно-, би- и мультиполярных нейронах. Дендрит служит входом нейрона, а аксон - выходом. Морфологически и функционально различают три главных типа нейронов: чувствующие, двигательные и вставочные. Нас в первую очередь будут интересовать чувствующие и вставочные.
Чувствующие нейроны. Тела чувствующих, или сенсорных, клеток обычно биполярной или мультиполярной формы* лежат всегда вблизи чувствующего органа или иннервируемой ткани. Дендриты одних нейронов, чаще всего биполярных, связаны с кутикулярными образованиями, других, всегда мультиполярных, - с тканями полости тела или же они образуют субэпидермальную сеть, как у личинок с мягкой кожей.
* (Некоторые авторы относят чувствующие клетки к особой категории нервных клеток и не называют их нейронами.)
Соответственно различают две большие категории чувствующих клеток. Клетки первого типа отличаются тем, что практически всегда связаны с кутикулой или ее впячиваниями: аподемами, трахеями, выстилкой предротовой и ротовой полостей и т. п. К ним принадлежат разнообразные экстерорецепторные клетки, в том числе и зрительные, хотя их дендриты выражены неясно. Клетки второго типа никогда не связаны с кутикулой и лежат только на внутренней поверхности тела, стенках пищеварительного тракта, в мышечной и соединительной тканях. Электрофизиологически показано, что они принадлежат интеро или проприоцепторам.
Аксоны чувствующих клеток идут непосредственно в соответствующие ганглии ЦНС, иногда находящиеся непосредственно в головном мозгу, например оптические или обонятельные центры. Вопрос о каналах связи рецепторных клеток с нервным центром чрезвычайно важен для правильной интерпретации работы анализатора и механизма управления поведением насекомого. Теперь, по-видимому, все признают несостоятельным прежнее мнение о том, что в некоторых рецепторных системах, например в антеннах клопа Rhodnius, имеет место срастание аксонов нескольких чувствующих клеток в единое волокно. Но замыкание группы рецепторов на один периферический нейрон второго порядка, т. е. потеря "адреса" входного сигнала, характерно для первого оптического ганглия насекомых. Смысл такого способа связи с центром, ведущего к частичной потере информации от совокупности датчиков, пока не всегда ясен (см. ниже).
Нервная ткань, в том числе и сенсорные клетки, происходят из эктодермы. Их принадлежность покрову тела выражается и в том, что связь чувствующего органа с ЦНС устанавливается центростремительно. Так, В. Вигглесворс показал на клопе Rhodnius, что перерезанный афферентный нерв регенерирует в направлении к ЦНС. Точно так же во время каждой линьки, когда образуются дополнительные рецепторы, чтобы обслуживать увеличивающуюся поверхность тела, их чувствующие клетки посылают аксоны центростремительно.
Выявленный на гистологических препаратах факт центростремительного развития аксона может стать одним из оснований для важного заключения о том, что путь от чувствующей клетки до ЦНС прямой, без синаптического переключения. Вблизи рецепторных клеток и афферентных нервов встречаются другие, например, нейроглиальные (питающие) клетки, но они не имеют отношения к передаче рецепторного сигнала.
Двигательные (моторные) нейроны. Тела этих обычно униполярных нейронов лежат всегда в сегментальном ганглии, чаще с его вентральной стороны и с боков. Их дендриты густо ветвятся в нейропиле ганглия, вступая в синаптические контакты с вставочными нейронами или даже непосредственно с разветвлениями аксонов чувствующих клеток. Аксоны двигательных нейронов уходят за пределы ганглия к мышцам, где заканчиваются множеством моторных концевых пластинок, или бляшек. Одну и ту же мышцу иннервируют так называемые быстрые и медленные двигательные волокна: по первым идут импульсы, вызывающие быстрые сокращения мышцы, по вторым - вызывающие тоническое сокращение. При этом тело иннервирующего двигательного нейрона может лежать в ганглии как того же сегмента, которому принадлежит мышца, так и соседнего. Часто встречается контралатеральное положение тела двигательного нейрона по отношению к эффектору.
Вставочные нейроны, или интернейроны, выполняют функцию передачи возбуждения с одного нейрона на другой. Они связывают чувствующие клетки с моторными, причем в последовательную цепь может быть включено несколько клеток, и тогда говорят об интернейронах 2-го, 3-го и более высокого порядка. Их обычно униполярные небольшие (от 2-3 мкм и меньше) тела лежат всегда по периферии ганглия, а отростки, в числе которых аксон, нередко т-образный, вступают внутрь ганглия, в его нейропиль. Интернейроны могут быть сегментальными или мультисегментальными, с восходящими и нисходящими волокнами. Последние участвуют, например, в управлении локомоторными реакциями. Благодаря множеству связей интернейроны образуют коммутационные сети для выработки нервных команд, направляемых по специальным трактам в составе комиссур и коннектив.
Среди восходящих волокон у насекомых обращают на себя внимание аксоны так называемых гигантских интернейронов, в частности, берущие начало в последнем брюшном ганглии и идущие по цепочке до грудных и даже до головных центров. Они отличаются большой длиной и толщиной (до 30 мкм и более), а главное, высокой скоростью проведения сигналов. Так, если в обычных афферентных двигательных нервах импульсы распространяются со скоростью 1,5-3,0 м/с, то в гигантском волокне, например таракана Periplanefa, скорость проведения может достигать 5-7 м/с. Считалось, что с помощью таких "быстрых" интернейронов осуществляется реакция убегания, когда опасность грозит насекомому сзади, но теперь этот взгляд признан несостоятельным.
Очень сложна область ганглиозного нейропиля, в котором генерируются нервные команды, управляющие поведением насекомого. Ее образует переплетение отростков дендритов моторных нейронов с разветвлениями ассоциативных и афферентных волокон, связанных друг с другом множеством синаптических контактов. Волокна здесь особенно тонкие: от 0,1 мкм по сравнению 1-3 мкм на выходе из дорсальных (двигательных) и вентральных (чувствующих) корешков. Они содержат протонейрофибриллы диаметром 100-200 А, митохондрии, гранулярный эндоплазматический ретикулум, нейросекреторные включения и прочие элементы. Синаптическая передача основана на поперечных и продольных контактах волокон или на контакте типа терминальной пуговки. Ультраструктура синаптических образований напоминает таковую синапсов у позвоночных: пресинаптическое волокно в районе контакта содержит синаптические пузырьки, наполненные химическим медиатором. Между трехслойными пре- и постсинаптическими мембранами находится синаптическая щель шириной 100-300 А. Как полагают, у насекомых имеется не только холинэнергический, но и адренэнергический механизм синаптической передачи. Не исключается также возможность электрической передачи возбуждения с помощью соответствующих синапсов, в которых импульсы передаются в результате электрического воздействия пресинаптического тока на мембрану постсинаптического волокна. Такие электрические сикапсы недавно обнаружены между зрительными интeрнейронами - детекторами движения, где они обеспечивают максимально высокую скорость проведения сигналов.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'