3. Физиологические механизмы сенсорной системы и методы их изучения
Последовательность процессов, протекающих в возбужденных рецепторах насекомых, сводится к следующей схеме. Адекватный раздражитель, взаимодействуя с рецептирующими элементами дендрита чувствующей клетки, вызывает такие локальные процессы, которые изменяют проницаемость их поверхностной мембраны. В результате возникают ионный ток и, как следствие, деполяризация клеточной мембраны, регистрируемая в виде медленного рецепторного потенциала (рис. 4, А, Б). Когда деполяризация достигает некоторого критического уровня, в определенной зоне клетки, как полагают, в аксонном холмике, возникают импульсы, или спайки, распространяющиеся по аксону в направлении ганглия. Мембрана хеморецепторных клеток под действием одних веществ деполяризуется, под влиянием других - гиперполяризуется. В последнем случае импульсный ответ в аксоне клетки затормаживается. Градуальный деполяризационный потенциал клетки называют также генераторным*, поскольку он приводит к генерации быстрого ответа клетки в виде импульсов, возникающих, как известно, по закону "все или ничего" (рис. 4, В). Импульсы бывают двухфазными или многофазными, как, например, в аксоне вкусовых и многих механорецепторных клеток. Монофазные импульсы механорецепторных нейронов отличаются увеличением их амплитуды по мере нарастания генераторного потенциала.
* (Во вторичночувствующих рецепторах позвоночных рецепторнын потенциал отличают от генераторного.)
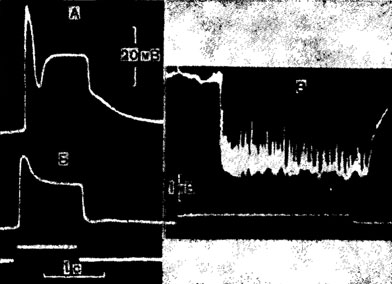
Рис. 4. Ответы рецепторных клеток. А, Б - рецепторный потенциал зрительной клетки мухи Luci-liа, зарегистрированный внутри-клеточно; запись на А сделана при освещении в тысячу раз выше, чем запись Б (по Naka, 1961). В - рецепторный потенциал с наложенными на нем импульсами, зарегистрированный вне-клеточно от вкусовой сенсиллы мухи Phormia regina; стимуляция 0,5 М раствором глюкозой (по Uehara, Morita, 1972)
Процессы трансформации энергии стимула, приводящие к изменению проницаемости рецепторной мембраны, пока изучены не настолько, чтобы можно было говорить о них с уверенностью и подробно (в какой-то мере исключение составляют фоторецепторы позвоночных животных).
Итак, если мы будем считать, что рецепторы насекомых являются первичными, то, следовательно, у них одна и та же сенсорная клетка и рецептирует стимул, и генерирует залп импульсов. Нет сомнений, что именно так обстоит дело в хеморецепторных или механорецепторных сенсиллах. Но зрительные клетки насекомых следует отнести скорее всего ко вторичным рецепторам, как и у высших животных. При их возбуждении, в период увеличения деполяризации, наблюдаются так называемые миниатюрные потенциалы, происхождение которых объясняют выделением порций медиатора из рабдомера, представляющего собой рецептор, объединенный с сенсорным нейроном - зрительной клеткой.
Деполяризационный ответ рецепторной клетки, как и частота импульсации в ее аксоне, в определенных пределах пропорционален логарифму интенсивности стимула. Максимальная амплитуда медленного ответа близка к величине потенциала покоя клетки - 30-70 мВ. В аксонах разных рецепторов импульсы могут следовать как в течение всего периода действия раздражителя, так и только после его включения (оn-ответ) или выключения (off-ответ); помимо оn-оff-ответов в волокнах часто регистрируется спонтанная (фоновая) импульсация. Соответственно различают тонические рецепторы, медленно адаптирующиеся к воздействию или неадаптирующиеся совсем, и фазовые, реагирующие только на изменение стимула. Фазовые рецепторы обычно быстро адаптируются - на повторные воздействия отвечают постепенно уменьшающимся ответом.
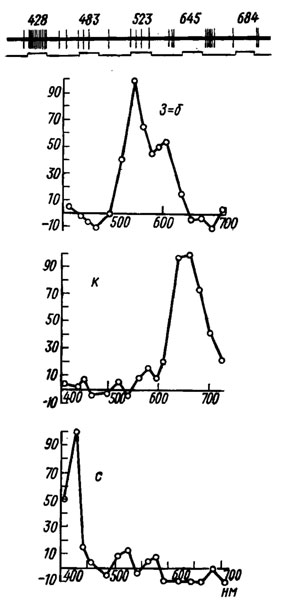
Рис. 5. Примеры ответов на спектральные стимулы мозговых интернейронов бабочки Рарillо troilus (по Swihart, 1970). Вверху: ответы с-нейрона; отклонение вверх нижней линии - период стимуляции, числа - длина волны света, нм. Внизу: графики зависимости им-пульсащии ют длины волны света; по оси ординат - импульсация в процентах от максимальной; по оси абсцисс - длина волны света, нм. з-б - 'зелено-белый' оппонентный нейрон: зеленые и белые лучи его возбуждают, а синие и красные частично тормозят его фоновую активность; к - 'красный' нейрон, возбуждающийся преимущественно оранжево-красными лучами; с - 'синий' нейрон, возбуждающийся преимущественно фиолетово-синими лучами
Интернейроны насекомых, как и других животных, активируются возбуждающими постсинаптическими потенциалами и соответственно затормаживаются тормозными постсинаптическими потенциалами. При возбуждении оптического интернейрона насекомых его мембрана, как правило, гиперполяризуется на десятки милливольт. Межнейронное взаимодействие осуществляется с помощью медиаторов - ацетилхолина, адреналина и, в виде исключения, посредством электрических импульсов. По типу ответа интернейроны могут быть весьма разнообразными, особенно в зрительном анализаторе. Встречаются on- или off-единицы, а также on-off-единицы; одни нейроны имеют фоновую активность, тормозящуюся при стимуляции рецепторов, у других она отсутствует; бывают адаптирующиеся к непрерывной стимуляции и неадаптирующиеся нейроны (рис. 5).
О сложном межнейронном взаимодействии в мозге насекомых при различных входных (рецепторных) сигналах можно судить по такому примеру. Замечено, что у мухи Boethcherisca peregrina с закрашенными фасеточными глазами на активность одиночных интернейронов переднего, среднего и заднего мозга, связанных с обонятельными сенсиллами и механорецепторами антенн, влияют сигналы от простых глазков. При освещении глазков мухи спонтанная активность одних единиц усиливается, а других тормозится. Выключение света по-разному сказывается и на ответах разных интернейронов, вызванных, например, пахучим стимулом: одни из них усиливаются, другие ослабляются. Первые встречаются преимущественно в среднем мозгу, вторые же приурочены к дорсальной области переднего и вентральной части среднего и заднего мозга.
Для того чтобы проникнуть в механизмы работы сенсорной системы, необходим разносторонний подход к ее изучению. Физиологию в широком смысле интересуют следующие важнейшие моменты:
- способы, которыми рецептор детектирует адекватные стимулы, т. е. выделяет и поглощает энергию адекватного раздражителя;
- молекулярные и ионно-обменные процессы в рецепторной клетке, приводящие в конечном итоге к генерации импульсов;
- способы кодирования рецепторной информации;
- механизм межнейронного взаимодействия и связи в ассоциативных центрах, а также способы обработки ими рецепторной информации.
Завершает эту цепь поиска вопрос о соответствии между видами поведения целого организма и детектируемой им информацией*.
* (В последнее время этот вопрос относят к области этологии животных.)
Коротко остановимся на подходах и методах, которыми располагает современная физиология органов чувств.
Морфофункциональный подход, традиционный для биолога, приобрел теперь более широкие и даже новые возможности благодаря использованию не только разных вариантов световой, но и электронной, в том числе сканирующей, микроскопии. Возможность различать внутриклеточные и межклеточные структуры, измеряемые ангстремами, служит хорошей основой для суждения о механизмах поглощения и трансформации внешней энергии в рецепторе и передачи нервного сигнала по аксону. Здесь велика роль гистохимии, особенно много давшей для расшифровки молекулярных процессов в возбужденных фоторецепторах.
Электрофизиологический подход. Успех в понимании работы сенсорной и всей нервной системы насекомых, достигнутый в последние 10 лет, обязан, пожалуй, в первую очередь именно электрофизиологическому подходу, в особенности микроэлектродному методу регистрации активности нервных клеток. Для внутриклеточного отведения электрической активности применяют тончайшие стеклянные микропипетки (диаметр кончика 0,6-0,3 мкм), заполненные 2М КСl. Такой микроэлектрод вводят в сенсиллу или нервную ткань до тех пор, пока скачком не изменится потенциал покоя на 40-60 мВ, что свидетельствует о проколе клеточной мембраны*. Внутриклеточным микроэлектродом можно регистрировать как генераторный потенциал клетки, так и ее импульсную активность. Активность нейронов может быть зарегистрирована и внеклеточным электродом. В зависимости от положения его кончика и точек приложения второго (индифферентного) электрода отводят либо медленный суммарный потенциал возбужденных клеток в виде, например, электроретинограмм (ЭРГ) или электроантеннограмм (ЭАГ), либо быстрый потенциал - экстраклеточные импульсы. Экстраклеточные электроды используют не только для регистрации ответа нейрона, но и для его стимуляции, например, током или химическим агентом, как при излучении вкусовых волосков. Через регистрирующий электрод специальной краской можно пометить место отведения, что особенно важно при внутриклеточной регистрации.
* (Поверхность клеточной мембраны имеет отрицательный знак по отношению к содержимому клетки. Поэтому введенный внутрь клетки микроэлектрод будет положительным относительно индифферентного электрода.)
Моделирование. Моделирование как способ изучения сенсорных органов и нейронных сетей все шире входит в арсенал физиолога. Исходя из параметров системы, создают либо физическую (например электронную), либо математическую модель ее. В последнем случае используют ЭВМ, для которой разрабатывают последовательность специальных команд-программу. Так, известны физические модели отдельных нейронов и целых систем, например цветового зрения некоторых животных. На ЭВМ многие исследователи пытаются моделировать механизм периферического кодирования признаков и центрального опознавания образов.
Поведенческий подход. Наблюдение за поведением животного следует считать, строго говоря, единственным адекватным способом для суждения о назначении и различительных возможностях его органов чувств. Биолога интересуют прежде всего те свойства сенсорных систем, которые реализуются в поведении, а точнее, те рецепторные сигналы, которые контролируют реакцию животного. Поэтому наблюдение за врожденными или приобретенными в ходе научения реакциями в норме и после нарушения целостности рецепторных органов или ассоциативных центров дает очень много для понимания их функции. Изменение поведения при частичной искусственно вызванной деафферентации позволяет выявить роль тех или иных рецепторов в управлении реакциями. Регистрируют как реакции отдельных частей тела, например выдвижение хоботка в ответ на стимуляцию вкусовых сенсилл или отдергивание ноги при ее механической стимуляции, так и моторные ответы целого организма - приближение к стимулу или уход от него. Реакция может быть как безусловнорефлекторной, так и на основе условного рефлекса, замыкающегося в простейшем случае, как показано известным физиологом Дж. Хорриджем (1962), на уровне грудных ганглиев нервной цепочки. Подключив последовательно к источнику тока симметричные лапки у двух обезглавленных тараканов Periplaneta americana, он заметил, что то насекомое, которое в процессе обучения получало удар тока лишь при опускании ноги, замыкавшей цепь "наказания", в контрольных опытах чаще держало ее приподнятой, чем партнер, получавший удар вне зависимости от положения своей ноги.
Особенно легко вырабатывается условный рефлекс у общественных перепончатокрылых пчел, муравьев, ос, чем исследователи широко пользуются для изучения механизмов рецепторного управления поведением. Например, кардинальные сведения о цветовом зрении и обонянии медоносной пчелы были получены именно в опытах с обучением насекомого.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'