Массовые появления вредных насекомых и их прогноз
Динамика популяций. Уже указывалось, что виды существуют в природе как популяции - группы близко родственных особей, образующих отдельные поселения. Численность особей в популяциях, и, следовательно, плотность особей в них не является стабильной, а изменяется под воздействием экологических факторов. Многие относящиеся сюда примеры уже были рассмотрены раньше.
Эти колебания численности особей в популяциях приобретают большое практическое значение. Прежде всего увеличение численности и плотности особей насекомого-фитофага, питающегося культурным растением, превращает это насекомое во вредителя: повреждение растений становится массовым, что приводит к снижению их продуктивности и даже к полной потере урожая. Обратный процесс - снижение количества особей насекомого-вредителя - снижает его вредоносность и при особо низкой плотности может способствовать переходу его во временно безвредное состояние. Важные практические последствия будет иметь изменение численности других вредных насекомых - кровососов, переносчиков болезней человека и домашних животных, различных паразитов и пр. Существенные практические последствия могут быть и при изменении численности полезных насекомых - энтомофагов, опылителей растений и др.
Изменения численности особей тех или иных видов стали предметом особого внимания исследователей, в результате чего были выяснены некоторые общие закономерности. Это изменение количества и плотности особей в настоящее время принято обозначать понятием динамика популяций, а раздел экологии, изучающий относящиеся сюда явления, - популяционной экологией. Действительно, эти количественные изменения того или иного вида реализуются в природе прежде всего через популяции видов.
Нередко для обозначения этих явлений применяют другое широко распространенное понятие - массовые размножения. Однако данное понятие не отражает всего существа популяционной динамики видов и не может считаться точным; в нем подчеркивается только одна сторона процесса - подъем численности вследствие увеличения плодовитости. Наконец, весь процесс изменения численности - от его начала через максимум до конца спада - иногда обозначают понятием градация.
Теоретический анализ динамики численности. Динамика популяций представляет собою сложное явление, ряд сторон которого в должной мере еще не изучен и не понят. Поэтому теоретический анализ причин изменения численности организмов и в частности их массовых появлений наталкивается на ряд трудностей. Существует ряд теорий, рассматривающих эту проблему по-разному.
Вместе с тем познание закономерностей популяционной динамики вредителей и разработка вопросов ее теории приобретает большое практическое значение для составления прогнозов возможных массовых появлений этих вредителей. На основе прогнозов могут быть приняты своевременные меры по предотвращению возможного вреда.
Чем же объясняются изменения численности вредных насекомых и каковы их причины?
В самом общем виде изменение численности может быть объяснено следующими двумя коренными причинами:
1) изменчивостью самой внешней среды, выходящей за пределы нормы требований и приспособлений вида к факторам среды;
2) изменением плодовитости и выживаемости особей под воздействием внешней среды.
При этом изменение численности может иметь двоякий характер: с одной стороны, изменяется плотность особей в популяциях (т. е. их число на 1 м2 или на одно растение), с другой стороны, изменяется число заселенных стаций. Следовательно, увеличение или уменьшение численности особей происходит не только в одном или в немногих местообитаниях вида, но может также охватить ту или иную часть его ареала, т. е. большое число популяций. Этими показателями определяются масштаб массового появления вредителя или, наоборот, его депрессии.
Плодовитость насекомых и способность их к размножению часто необычайно велика. Нередко эту способность к размножению обозначают понятием потенциал размножения, или биотический потенциал. Наиболее рационально им обозначать не плодовитость вида вообще, а теоретический максимум потомков, получаемый от одной пары особей (при партеногенезе - от одной особи) за весь год. Например, яблонная плодожорка откладывает в среднем 100 яиц, поэтому ее биотический потенциал при двух поколениях составит на одну пару особей (при равном числе самцов и самок в популяции) 502, т. е. 2500. У тлей, дающих за лето до 15 и более партеногенетических поколений при той же самой плодовитости, т. е. 50 особей на самку, биотический потенциал достигает астрономических показателей - в данном примере 5015, т. е. миллиарды миллиардов особей.
Акад. В. И. Вернадский рассматривал размножение организмов как проявление одного из свойств живой материи - способность распространяться по земной поверхности в результате производимой химической работы и создания новых количеств живого вещества. Эту способность он обозначил понятием скорость передачи жизни, которая является величиной постоянной и характерной для каждого вида организмов; она определяется размерами и массой тела, половой продуктивностью, числом поколений в тот или иной отрезок времени и требованиями к среде обитания. В целом скорость передачи жизни характеризует геохимическую энергию видов и выражается числом см/сек.
Например, скорость передачи жизни у нестадных саранчовых составляет примерно 13-15 см/сек, а у лугового мотылька 45 см/сек; это значит, что распространение этих насекомых завершилось бы на Земле, учитывая длину экватора примерно в 40 тыс. км, в первом случае в течение около 9 лет, а во втором - около 3 лет.
Биотический потенциал и скорость передачи жизни являются теоретическими абстракциями и в реальной природе размножение организмов никогда не соответствуют этим величинам. Однако оба эти понятия ценны тем, что позволяют установить для видов численные показатели их потенциальной энергии размножения.
Невозможность полной реализации в природе биотического потенциала видов - следствие ограничивающего воздействия внешней среды: под ее воздействием происходит либо снижение плодовитости, либо гибель части потомства.
По вопросу о роли факторов среды и механизма сдерживания численности особей в популяциях существуют различные точки зрения. Широким распространением пользовалась теория северо-американского эколога К. Чэпмана, который рассматривал все факторы среды как враждебные организму силы природы; эти силы он обозначил понятием сопротивление среды. Численность особей вида считалась данным исследователем лишь итогом взаимодействия между биотическим потенциалом и сопротивлением среды.
В самом общем виде эта теория в известной мере раскрывает механизм динамики численности популяций, но не может считаться удовлетворительной, так как рассматривает организм и среду лишь как враждебные системы. Между тем организм является порождением среды и вместе они выступают не как враждебные, а как эволюционно согласованные системы (см. также "Вводные понятия" к разделу "Экология").
Возникающие между организмом и средой противоречия временны, происходят вследствие изменчивости условий среды и отклонений ее от той средней нормы, к которой организм оказался приспособленным. Эти отклонения от нормальных показателей постоянно устраняются, и тем самым соответствие организма и среды постоянно восстанавливается. В конечном итоге изменение численности особей в популяциях может рассматриваться как результат тех воздействий, которые нарушают и восстанавливают согласованность системы - организм и среда.
Теория Чэпмана, как и высказанные по ее поводу положения, имеют слишком общий характер и не раскрывают сложного механизма популяционной динамики видов. В настоящее время по вопросу о динамике численности насекомых существуют две основные теории.
Одна из них главенствующую роль приписывает абиотическим факторам среды, хотя не исключает значения и биотических факторов. Случайные комбинации этих факторов определяют ход численности особей в популяциях; популяции, следовательно, не регулируются, а стихийно изменяются, варьируют. Эта точка зрения разделялась известным израильским исследователем доктором Ф. Боденхеймером.
Противоположную позицию занимают многие другие исследователи как у нас, так и за рубежом. Они считают, что численность особей в популяциях определяется двумя категориями факторов - нереактивными и реактивными; первые обозначаются также независимыми, а вторые - зависимыми от плотности популяций.
Нереактивные факторы - это преимущественно абиотические факторы; они действуют на популяции вне зависимости от плотности особей в них, т. е. при любой численности вида. Например, слишком суровые зимы вызывают гибель зимующих особей капустной белянки или озимой совки вне зависимости от того, в большом или в малом количестве особей существуют эти виды в данный зимний период. Или, наоборот, благоприятные условия зимовки могут способствовать повышению численности особей как в разреженных популяциях, так и в плотных. Следовательно, эти факторы среды могут вызывать значительные колебания численности особей.
Реактивные факторы выступают уже как регулирующие численность особей в популяциях. Это биотические факторы среды - естественные враги (хищники, паразиты, возбудители болезней) и пища. Как только плотность популяции того или иного вида вредителя достигнет для данного биоценоза своего возможного в этом биоценозе верхнего предела, начинают действовать эти реактивные факторы: обилие особей насекомых-фитофагов благоприятствует росту численности энтомофагов, тесный контакт особей благоприятствует возникновению грибных, бактериальных и других болезней, создается также недостаток пищи для фитофага. В результате численность особей насекомого-фитофага в популяциях сокращается, что сокращает и численность энтомофагов.
Без такой регулирующей роли реактивных факторов могло бы происходить несдерживаемое увеличение численности насекомых-фитофагов в популяциях и в биоценозах, что в конце концов повело бы к уничтожению кормовых растений, к гибели вследствие этого насекомого-фитофага и всего биоценоза в целом. Регуляция численности рассматривается в этой теории не как следствие сопротивления среды, а как процесс, возникший в биоценозе в результате сложившихся взаимодействий между отдельными видами на основе цепей питания.
Эта теория саморегулирования популяций, следовательно, не отрицает большого значения нереактивных факторов как случайных изменений в абиотической среде; но им приписывается строго определенная роль - эти факторы определяют лишь колебание численности и плотности особей в популяциях. Регулирующая же роль принадлежит реактивным факторам, т. е. биотической среде.
В настоящее время благодаря успехам в области кибернетики и теории автоматического регулирования и управления идея о саморегулировании популяций приобретает особый интерес. Некоторые исследователи считают возможным приложить теорию автоматического регулирования к пониманию популяционной динамики в биоценозах. Одно из положений этой теории состоит в том, что в саморегулируемых системах регулируемый процесс связан с регулятором двусторонне - прямой и обратной связью. Следовательно, регулятор - в данном случае биотические факторы - меняет свое воздействие на регулируемый процесс, т. е. численность особей, в соответствии с теми изменениями, которые происходят в численности.
Действительно, многие исследования и наблюдения показали, что при росте численности и плотности особей в популяциях происходит усиление воздействия биотических факторов, а при снижении их численности и плотности роль биотических факторов также снижается. Благодаря этому популяция не гибнет и вместе с тем ее плотность не может увеличиваться беспредельно несмотря на высокий биотический потенциал составляющих ее особей. Такое взаимодействие с обратной связью может иметь место только между живыми организмами - например, насекомыми-фитофагами и их энтомофагами; те и другие обладают способностью реагировать на изменения внешней среды, чего лишены абиотические факторы.
Таким образом, высокий биотический потенциал постоянно обеспечивает в биоценозе возможность избыточного роста численности, а биотические факторы сдерживают и регулируют этот рост, выполняя буферную функцию. Если же буферная роль биотических факторов в популяциях того или иного вида снизится, а абиотические факторы окажутся благоприятными, неизбежно произойдет резкое увеличение численности и плотности особей данного вида.
Такое ослабление регулирующей роли биотических факторов часто наблюдается во вторичных биоценозах, например, на посевах и посадках культурных растений. Применяемая агротехника может оказаться гибельной для полезных энтомофагов, а обилие пищи в виде возделываемого культурного растения исключает регулирующую роль пищевого фактора. Вот почему агробиоценозы являются той основной биоценотической средой, где особенно часто происходят массовые вспышки насекомых-фитофагов, которые и становятся здесь вредителями.
Быстрота ответа биотических факторов на изменение плотности популяции и, следовательно, регулирующая их роль неодинакова. Наиболее быстродействующей реакцией обладают многоядные энтомофаги; они способны переключаться на питание любым видом насекомого-фитофага, численность которого начинает в биоценозе возрастать. Более специализированные энтомофаги, т. е. ограниченноядные и одноядные хищники и паразиты, действуют с более замедленным эффектом - необходимо некоторое время для их размножения. При более высоких плотностях насекомого-хозяина создаются условия для действия различных патогенных, вызывающих заболевания, организмов; этому может благоприятствовать и ослабление физиологического состояния данного насекомого. Наконец, важными регулирующими факторами являются внутривидовая конкуренция и пища; они предотвращают чрезмерное размножение насекомого, а отсюда уничтожение пищевой базы и гибель популяции.
Для насекомых-фитофагов, в том числе и вредителей растений, наибольшую регулирующую роль играют их естественные враги - хищники, паразиты, возбудители болезней; благодаря этому фитофаги лишаются в природных биоценозах возможности уничтожения своих кормовых растений - энергетической основы биоценоза. У хищников, паразитов и потребителей мертвой органической продукции - сапрофагов и некрофагов - решающая регулирующая роль принадлежит пищевому фактору и внутривидовой конкуренции.
Типы динамики популяций и прогноз численности. Изменение численности особей происходит у разных видов по-разному. Установление специфики популяционной динамики отдельных видов вредителей создает научные основы для разработки методов прогноза, изменения их численности в практических целях - с целью принятия необходимых мер борьбы.
Отвлекаясь от частностей, все многообразие популяционной динамики можно, свести к трем основным типам - устойчивому, сезонному и многолетнему.
Устойчивый тип динамики численности характерен для видов с более или менее постоянной численностью в течение всего вегетационного периода. Количество особей в популяциях у таких видов мало изменяется в течение сезона вследствие наличия у них соответствующих приспособлений, обеспечивающих им высокую выживаемость на протяжении всего года. Высокая выживаемость сопряжена у видов этого типа с низкой плодовитостью; последней достаточно для поддержания более или менее постоянного уровня численности во все сезоны года.
К этому типу относятся такие вредные насекомые как хрущи, хлебные жуки, жуки-щелкуны и их личинки - проволочники, жуки-чернотелки и их личинки-ложнопроволочники.
Для прогноза вероятной численности этих видов необходимо произвести учет их плотности и распределения на различных полях в осеннее время. Исходя из типа популяционной динамики этих видов, можно с достаточной степенью вероятности сказать, что в будущем году численность и плотность данных вредителей существенно не изменится, если не будут применены меры борьбы.
Сезонный тип динамики численности (рис. 189) характерен для видов с резко возрастающей плотностью популяций в течение одного сезона. Другими словами, количество особей у видов этого типа может сильно увеличиться в течение одного вегетационного периода, т. е. с весны до осени, однообразно повторяясь ежегодно. Сюда относятся многие поливольтинные виды и некоторые высоко плодовитые моновольтинные виды: капустная моль, хлопковая совка, различные виды тлей, яблонная плодожорка, яблонная моль, боярышница, златогузка, гессенская и шведская муха, комнатная муха и др.
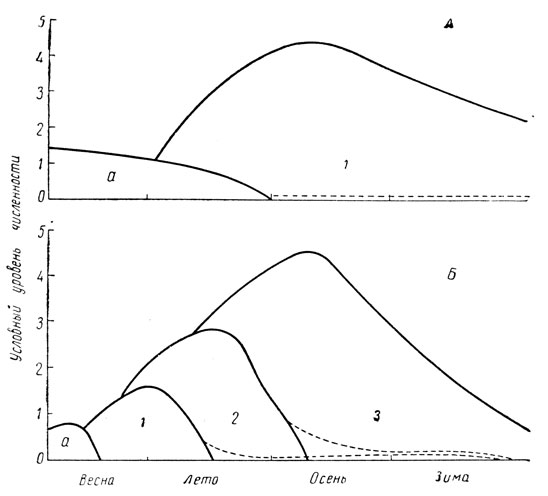
Рис. 189. Кривые изменения численности особей при сезонном типе популяционной динамики. А - у моновольтинных видов; Б - у поливольтинных видов (по И. Полякову): а - перезимовавшая часть популяции, 1-3 - соответствующие поколения
Эти виды появляются весной в виде того или иного количества перезимовавших особей - в фазе яйца (тли), личинки (яблонная плодожорка, боярышница, златогузка), куколки (капустная моль, хлопковая совка) или имаго (комнатная муха).
Большое число поколений или высокая плодовитость обеспечивают этим видам быстрое увеличение численности особей за один вегетационный период. Например, потомство одной пары капустной моли при средней плодовитости 200 яиц на самку при 3-4 поколениях могло бы достичь величины 1003 или 1004; хотя этого не происходит, но во всяком случае численность особей от поколения к поколению сильно увеличивается (рис. 189, В). Даже у таких моновольтинных видов как известные вредители плодового сада - боярышница и златогузка - численность особей значительно увеличивается за один сезон (рис. 189, А); расчет по биотическому потенциалу дает увеличение даже в 150-250 раз, что однако никогда не наблюдается.
Таким образом для видов этой группы характерно численное возрастание особей во второй половине сезона или осенью, что и должно служить основой для прогноза и планирования мер борьбы. Для большинства видов этого типа целесообразно проведение осеннего учета их численности.
Многолетний тип популяционной динамики отличается наибольшей сложностью и разнообразием проявлений. Изменение численности особей, плотности в популяциях и заселенности стаций охватывает период в несколько лет и сопровождается прохождением серии фаз популяционной динамики. Обычно различают 4-5 таких фаз, название которых и вкладываемое содержание не всегда совпадают у различных авторов. В предлагаемой ниже схеме (рис. 190) эти фазы сведены к следующим:
1) фаза минимума, или депрессии, - численность насекомого минимальная, причиняемый им вред незаметен;
2) фаза подъема, или нарастания, называемая иногда также продромальной, - численность насекомого и плотность его в популяциях под влиянием благоприятных условий увеличивается, он расселяется на новые местообитания, но вред проявляется еще незначительно или не в полной мере;
3) фаза максимума, или массовой вспышки, называемая иногда также эруптивной, - насекомое достигает наибольшей плотности в популяциях и в заселяемых стациях, находится в состоянии экологического и физиологического оптимума и причиняет особо значительный вред;
4) фаза спада, или кризиса, - численность начинает снижаться под влиянием неблагоприятных условий, в первую очередь биотических факторов (естественные враги, недостаток пищи), плотность в популяциях и число заселенных стаций уменьшается, вредоносность резко падает.
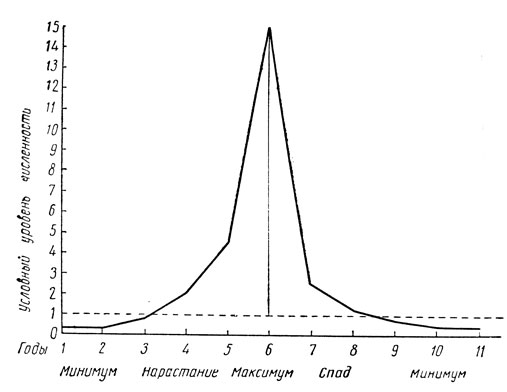
Рис. 190. Кривая изменения численности особей при многолетнем типе популяционной динамики (по Драховской)
Общая продолжительность всего полного цикла фаз популяционной динамики при многолетнем типе достигает у моновольтинных видов минимум 4 лет, а у многих видов 6-10 и более лет; у би- и тривольтинных видов весь цикл может охватить 2-3 года (рис. 190).
Виды с многолетним типом динамики численности наиболее многообразны и могут быть подразделены на ряд подтипов, подробное рассмотрение которых здесь не дается. Примерами могут служить саранчовые, вредная черепашка, свекловичный долгоносик, хлебная жужелица, озимая совка, колорадский жук и многие другие известные виды вредителей.
Так, для саранчовых характерно то, что их очаги, т. е. местообитания, где они накапливаются в большой численности, располагаются за пределами агробиоценозов - на целинных землях, залежах, пастбищах и пр. Плодовитость сравнительно невелика, в умеренных странах развивается одно поколение. Массовая вспышка при особо благоприятных погодных и других условиях может произойти в некоторых случаях после двух оптимальных лет, но нередко один период популяционной динамики растягивается на 10 и более лет. В фазу подъема и максимума происходит резкое увеличение заселенной площади, особенно у стадных видов.
Прогноз вероятной численности этих вредителей на будущий год строится на осенне-весеннем учете территорий, зараженных кубышками, на анализе предшествовавших климатических и других факторов, а у стадных видов - также на определении стадного состояния популяций, т. е. соотношений стадной, промежуточной и одиночной форм.
Вредная черепашка, свекловичный долгоносик и хлебная жужелица так же, как и саранчовые, имеют одну генерацию в году, но в отличие от них тесно связаны с обрабатываемыми землями, т. е. агробиоценозами, где имеют устойчивую кормовую базу. Вследствие этого численность этих видов на посевах всегда более или менее высокая, погодные и другие условия не могут быстро и значительно снизить плотность популяций.
Для прогноза вероятной численности видов этой группы в будущем году необходим учет их распределения и численности по полям, анализ экологических условий развития до зимовки, а также учет физиологического состояния самого вредителя.
Другие группы многолетнего типа динамики численности имеют свои специфические свойства, которые здесь не рассматриваются.
Из всего сказанного выше становится ясным, сколь важное значение для разработки надежных прогнозов имеет глубокое изучение биологии и экологии каждого вида вредителя в отдельности и установление характерных особенностей его популяционной динамики.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'