Местообитание и ареал как экологические явления
Каждый вид и слагающие его популяции занимает в природе ту или иную территорию, т. е. владеет определенным пространством. Пространственное распределение вида в значительной степени или полностью определяется экологическими условиями среды и избирательным отношением вида к ней. Основными формами пространственного распределения организмов на земной поверхности является заселение местообитаний и географическое распространение.
Распределение по местообитаниям. Избирательное отношение видов к факторам среды порождает избирательность и к местообитаниям, т. е. экологическую специализацию по отношению к участкам территории, которые он пытается занять и заселить. Участок территории, занятый популяцией вида и характеризующийся определенными экологическими условиями, называется местообитанием вида, или стацией.
Так, пшеничное поле является стацией пшеничного трипса, шведской мухи, гессенской мухи, разных видов клопов-черепашек и хлебных жуков, хлебной жужелицы и других видов. Стациями азиатской саранчи будут пониженные, обычно поросшие тростником болотистые участки по берегам рек, озер и морей на юге СССР, а стациями майского хруща - сосновые боры и другие лесные массивы, поляны, вырубки.
Каждая из таких стаций характеризуется своим растительным покровом, свойствами почвы, особенностями микроклимата и пр. Следовательно, каждая стация является как бы суммированным показателем условий среды, свойственных определенному участку земной поверхности.
Как оказалось, каждый вид насекомого имеет свой характерный набор заселяемых стаций; у одних видов этот набор разнообразен, у других - строго ограничен определенным типом местообитания. Между этими крайними показателями избирательности вида к местообитаниям существуют разнообразные переходы. Например, для азиатской саранчи характерна, как указывалось, строгая связь с болотистыми стациями, а итальянская саранча встречается в более разнообразных местообитаниях - на целинных степных участках, залежных землях, пастбищах и пр. Пшеничный трипс, шведская и гессенская муха встречаются лишь на посевах хлебных злаков и на злаковниках с участием луговых злаков, например, на лугах или пырейных залежах. А капустная совка может встречаться не только на овощных полях и огородах с капустой и брюквой, но также на посевах свеклы, гороха, подсолнечника, на плантациях табака, клеверищах и в других местообитаниях.
Набор заселяемых тем или иным видом стаций столь характерен для каждого вида, что может служить важнейшим видовым отличительным признаком, не менее значимым, чем морфологические и другие отличия. Более того, исследования показали, что в пределах родственной группы насекомых (представителей одного и того же рода или семейства) невозможно подобрать пару видов, которые бы заселяли один и тот же комплекс стаций.
Это свойство видов избирательно заселять те или иные стации представляет собою важнейшую экологическую закономерность, обозначаемую как принцип постоянства в выборе местообитаний, или принцип стациальной верности. Очевидно изученность и характеристика любого вида насекомого, особенно имеющего практический интерес, не может считаться удовлетворительной без сведений о распределении его по стациям.
Итак, стация есть местообитание вида. Однако в некоторых случаях местообитание вида обозначают понятием биотоп. Такое смешение терминов нельзя считать целесообразным; под понятием биотоп лучше подразумевать участок территории, заселенный сообществом организмов, т. е. их естественным комплексом. Такие комплексы, т. е. сообщества взаимосвязанных между собою организмов, обозначаются понятием биоценоз (см. следующий раздел "Биоценология насекомых"). Следовательно, биотоп представляет собою единицу местообитания биоценоза, а не вида.
Принцип смены местообитаний. Установление избирательной привязанности организмов к стациям явилось важным итогом полевого экологического изучения видов. Однако дальнейшие исследования показали, что принцип стадиальной верности оказывается действительным лишь в условиях ограниченного диапазона пространства и времени. В широком их диапазоне возникает диаметрально противоположное явление - закономерное изменение видами своих местообитаний. Эта закономерность обозначена нами как принцип смены местообитаний.
Смена местообитаний как экологическая закономерность представляет собою сложный комплекс явлений, протекающих частью в пространстве и частью во времени. В широком диапазоне пространства смена местообитаний проявляется в виде зональной и вертикальной смены стаций и зональной смены ярусов; а в широком диапазоне времени - в виде сезонной и годичной смены стаций.
Зональная смена стаций характерна для видов широко распространенных, заселяющих ряд природных зон, т. е. для трансзональных видов. Проявляется зональная смена стаций как закономерно направленное изменение местообитаний при переходе вида из одной природной зоны в другую: при продвижении к северу избираются более сухие, хорошо прогреваемые открытые стации с разреженным растительным покровом, часто располагающиеся на легких песчаных или каменистых почвах; при продвижении к югу тот же вид заселяет более увлажненные и тенистые стации с густым растительным покровом и с глинистыми почвами.
Примеры зональной смены стаций у трансзональных видов приведены в таблице 8.

Таблица 8
Из таблицы видно, что перелетная саранча в средней полосе Европейской части СССР заселяет песчаные места, а на юге - болотистые стации. Итальянская саранча близ северной границы распространения (например, в центральных черноземных областях и в Западной Сибири) живет на склонах сухих меловых обнажений с редким травостоем и на песчаных местах, а в Средней Азии и в Закавказье заселяет хорошо орошаемые участки с густым травостоем, например, люцерновые поля в оазисах; в Средней Азии этот вид даже называется оазисным прусом.
Сходным образом происходит смена стаций и у других приведенных в таблице видов - майского и июньского хрущей и озимой совки. Изменение стаций у майского хруща при продвижении с севера Европейской части СССР к центру и далее к югу показано также на карте (рис. 183).
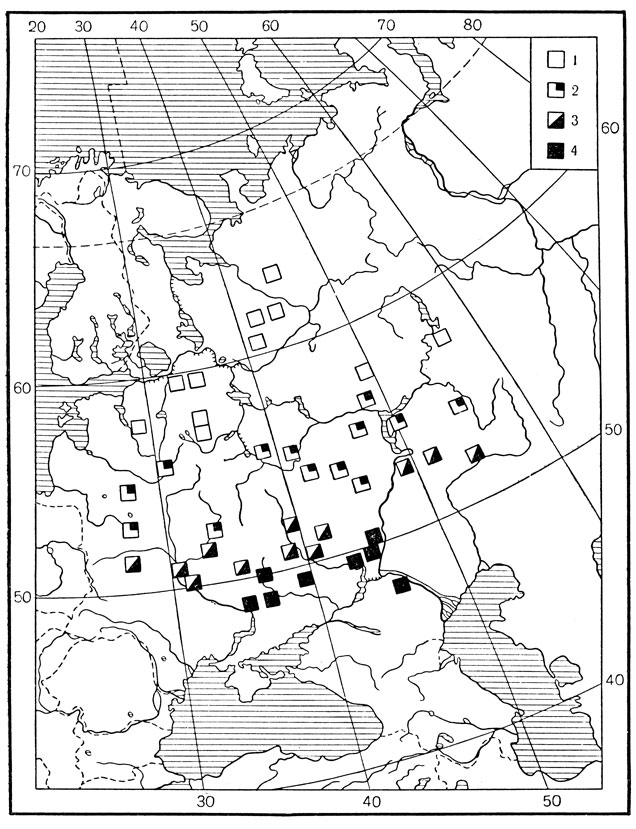
Рис. 183. Зональная смена местообитаний у восточного майского хруща в Европейской части СССР (по Березиной): 1 - встречается вне лесного полога, 2 - вне лесного полога и под слабым затенением, 3 - под пологом леса и редко вне его, 4 - под пологом леса
Графически зональная смена пояснена на специальной диаграмме (рис. 184). Все стации на этой диаграмме распределены по характеру растительного покрова и микроклимату на три экологические группы - ксерофитные (сухие теплые стации с разреженным травостоем), мезофитные (умеренно или средне увлажненные) и гигрофитные (сильно увлажненные с густым травостоем). Смещение видов в более влажные стации при продвижении к югу показано косыми стрелками; при этом влаголюбивые виды лесной и отчасти лесостепной зоны лишаются возможности проникнуть в более южные зоны, так как более влажные стации, чем гигрофитные, физически и экологически немыслимы (на рис. 184 показано прерывистыми стрелками).
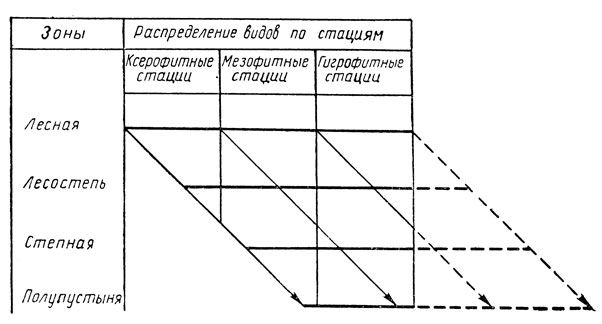
Рис. 184. Диаграмма зональной смены стаций (по Бей-Биенко)
Зональная смена стаций является экологическим следствием закона географической зональности и объясняется возрастанием по мере движения на юг количества тепла, получаемого земной поверхностью. Так, количество солнечного тепла (эффективной радиации) за вегетационный период, т. е. с апреля по сентябрь включительно, воспринимаемое 1 см2 земной поверхности, изменяется по географической широте следующим образом (в больших калориях):

Резкое возрастание эффективной радиации по мере движения на юг ставит расселяющийся здесь вид в условия избытка тепла. Внешне одинаковые стации на севере и юге в действительности резко различаются между собою по тепловому режиму; например, температура на поверхности песчаного участка в лесной зоне летом обычно не превышает 35-40°С, а в пустыне она достигает 65°С и выше.
Этот избыток тепла на сухом юге может быть устранен путем перехода в более увлажненные и тенистые стации; высокая влажность в таких стациях при сухом воздухе способствует охлаждению через усиленное испарение, а тень умеряет прогревающий эффект южного солнца. В конечном итоге в таких влажных стациях создается более прохладный микроклимат, что и позволяет здесь жить некоторым северным видам. С другой стороны, недостаток тепла и избыток влаги на севере компенсируется переходом на более сухие и теплые стации.
Вертикальная смена стаций аналогична зональной, но проявляется в горных условиях. Наиболее обычной формой вертикальной смены стаций является переход видов на более ксерофитные стации по мере повышения вертикального уровня. Например, серый кузнечик в лесном поясе Кавказа держится на мезо- и гигрофитных стациях, а в альпийском - в ксеро- и мезофитных.
Зональная смена ярусов проявляется в том, что трансзональные виды, как показал М. С. Гиляров и другие авторы, в разных зонах занимают неодинаковые ярусы. Так, в более сухих зонах они нередко из наземных видов становятся хотя бы частично почвенными; с другой стороны, некоторые виды при продвижении на север перемещаются из более высокого растительного яруса в более низкий (табл. 9).

Таблица 9
Самым необычным и удивительным из приведенных примеров является переход таких, казалось, наземных видов как короеды к жизни на корнях, т. е. к подземному образу жизни.
В итоге зональная смена стаций и ярусов и вертикальная смена стаций ставит вид в двойственные и противоречивые условия. С одной стороны вид предъявляет к среде определенные требования, вытекающие из его наследственных физиологических свойств; с другой стороны, при успешном расселении в другие зоны и в горах вид вынужден переместиться на другие стации или даже сменить ярус. Все это приводит к важным последствиям: изменяется экология, физиология и биология видов, что стимулирует внутривидовую дифференциацию; смена стаций становится одним из факторов эволюции.
Зональная и вертикальная смена местообитаний имеет существенный интерес и в практическом отношении. Она должна учитываться при разработке зональных мер борьбы с вредителями, может дать основания для понимания изменения экологии вида в разных зонах и прогноза этих изменений при таких мероприятиях как орошение земель и создание лесных массивов на юге, как осушение земель, возделывание пропашных культур, вырубка леса, применение черных и занятых паров на севере и пр.
Смена местообитаний во времени связана с изменениями микроклимата в течение одного сезона или в течение ряда лет. При изменениях микроклимата в течение одного и того же сезона происходит сезонная смена стаций. Она бывает особенно резко выраженной в областях с жарким и сухим климатом.
Одной из наиболее обычных форм сезонной смены стаций является переселение степных и пустынных видов в период летней засухи и выгорания природной растительности на более увлажненные местообитания с зеленым и густым растительным покровом - на посевы, на луга и в западины, под полог леса. Так, в Нижнем Поволжье многие виды насекомых летом мигрируют в лиманы - затопляемые участки с более богатой, чем в степи, растительностью; другой пример - переселение вредных кузнечиков в Крыму и на Кавказе с мест, где травостой выгорел на посевы, с последующим повреждением этих посевов. Переселение насекомых в период сенокоса со скошенной площади на посевы и другие участки с густым растительным покровом также может рассматриваться как одна из форм сезонной смены стаций.
Отклонение погодных условий от средней нормы в ту или иную сторону отражается на микроклимате занимаемых насекомыми местообитаний и приводит к годичной смене стаций. В более сухие и теплые годы происходит переселение видов на влажные стации, а во влажные годы - на более сухие стации.
Так, перелетная саранча в южном Казахстане в сухие годы концентрируется на западинах с более влажной почвой и относительно густым растительным покровом; здесь же происходит и откладка кубышек. В более влажные годы, наоборот, избираются более возвышенные сухие места, где откладываются и кубышки.
В центральной Украине переселение в сухие жаркие годы на более влажные стации отмечено у большого и разнообразного комплекса видов насекомых; при этом на влажных стациях происходит значительное увеличение количества встречающихся видов и их обилия (численности на 1 м2), за исключением видов северного происхождения. В сырые и прохладные годы происходит не только обогащение фауны сухих стаций, но и расширение заселяемой территории у мезо- и гигрофилов при сокращении обилия ксерофилов и видов южного происхождения.
Таким образом, путем смены местообитаний виды в изменяющихся условиях среды изыскивают для себя наиболее подходящие условия жизни. В сущности смена местообитаний позволяет этим видам сохранять свой экологический стандарт, свои требования к гидротермическим факторам даже в изменяющейся среде.
В целом, принцип постоянства в выборе местообитаний, как и его антипод - принцип смены местообитаний, свидетельствуют о сложности взаимоотношений вида с той средой, которая создается на заселяемых видами участках территории земной поверхности. Без учета и понимания этих взаимоотношений вида с местообитанием невозможно понять его экологию и некоторые стороны его биологии, что будет затруднять и разработку необходимых практических мероприятий - мер борьбы с вредными видами и мер содействия полезным видам.
Ареал вида. Область географического распространения вида нередко обозначается кратким понятием ареал. Последний в сущности является географически увеличенной стаций и, следовательно, зависит как от избирательных требований вида к среде, так и от суммы экологических условий, слагающихся на территории ареала, - климата, растительного покрова и пр.
Однако по сравнению со стацией ареал вида представляет собою еще более широкое и сложное явление. Он определяется не только современными экологическими условиями, но и геоисторическими воздействиями, т. е. изменениями физико-географических условий в прошлом - климата, растительного покрова, формы земной поверхности и пр. Следовательно, ареал вида есть суммированный эффект современных и предшествующих условий.
Процесс изменения ареалов во времени изучается исторической зоогеографией и здесь не рассматривается. Что касается современных факторов, влияющих на географическое распространение видов, то из них наибольшее значение имеют термические условия, влажность среды и наличие подходящих кормовых ресурсов.
Роль климатических факторов в распространении насекомых проявляется разнообразно. Прежде всего это сказывается на числе видов: более теплые и влажные области нашей планеты имеют наиболее богатую фауну. Особенного расцвета и исключительного разнообразия насекомые достигают в тропиках, тогда как в умеренном климате они значительно менее разнообразны, а в полярной зоне представлены сравнительно немногими видами.
Если разделить земной шар на три климатические зоны - жаркую (тропики и субтропики), умеренную и холодную (полярные области), то число видов в этих зонах среди, например, таракановых будет примерно таковым: 2500-150-2. Эти цифры с очевидностью указывают на исключительное богатство тропической и субтропической фауны данного отряда и о почти полном отсутствии названных насекомых в полярных странах.
На территории СССР (площадь 22,4 млн. км2), лежащего почти целиком в умеренной и холодной зонах, встречается около 500 видов саранчовых, тогда как в Австралии (8,9 млн км2), которая в 2,5 раза меньше СССР, но лежит целиком в жаркой зоне, найдено 800 видов саранчовых. Из Африки (около 30 млн. км2) известно до 2000 видов саранчовых, хотя площадь этого континента только на 1/3 больше территории СССР.
Фауна насекомых полуострова Индостан вместе с прилежащей Бирмой (площадь 3 млн. км2), лежащих целиком в жаркой зоне, по старым и несомненно преуменьшенным данным насчитывает 30000 видов, а территория Гренландии (около 2,2 млн. км2), располагающаяся полностью в холодной зоне, имеет только около 450 видов.
Приведенные данные показывают выдающееся значение климатических факторов в распространении насекомых и в определении числа и состава встречающихся в той или иной стране видов. Из этих данных также вытекает, что в определении северной или южной границ распространения видов весьма существенную роль должен играть термический фактор. Особенно это касается северной границы распространения видов на континенте Евразии.
Одной из важнейших климатических границ на территории Евразии является январская изотерма в -20°С и изотерма средних абсолютных минимумов температуры -25°С (рис. 185). Эти две изотермы ограничивают распространение к северу и к востоку многих членов недостаточно холодостойкой европейской фауны насекомых, таких как капустная белянка, озимая совка, кольчатый шелкопряд и многие другие виды. Поэтому они широко распространены в Европейской части СССР, но отсутствуют на северо-востоке ее, не заходят в Западную Сибирь, но некоторые встречаются в Средней Азии и на юге Дальнего Востока. Объясняется это тем, что указанные изотермы от северо-запада Европы сильно спускаются вниз к юго-востоку ее и далее на востоке проходят южнее Западной Сибири. Благодаря этому фауна насекомых Западной Сибири имеет обедненный состав европейских видов, но вместе с тем включает ряд более холодостойких сибирских видов, хотя бы частью отсутствующих в Европейской части СССР, например сибирскую кобылку из саранчовых.
Характерно, например, что весь отряд таракановых, насчитывающий в СССР около 60 видов, совсем не представлен в Сибири, если не считать единственного европейского вида, проникающего лишь на юг Западной Сибири. Северная граница ареала этого отряда в Евразии проходит чуть южнее январской изотермы в -20°С. Следовательно, громадная территория Северной Сибири, лежащая к северу от этой изотермы, полностью лишена природной фауны таракановых; но на юге советского Дальнего Востока представители этого отряда вновь появляются в природе. В целом северная граница распространения отряда таракановых в Евразии является ярким свидетельством того, насколько велика роль зимних температур в ограничении продвижения к северу недостаточно холодостойких насекомых (рис. 185).
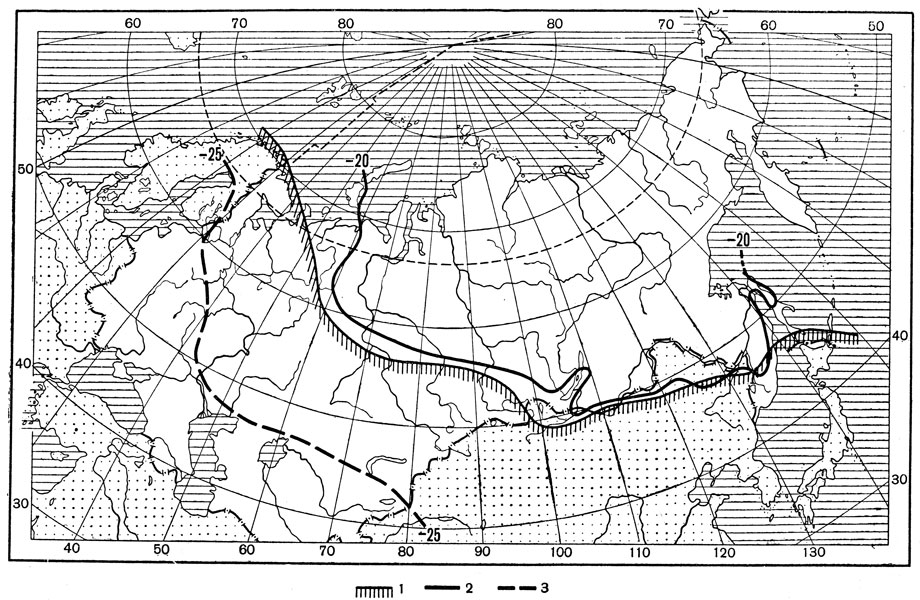
Рис. 185: Карта умеренной Евразии с показом северной границы распространения отряда таракановых и важнейших изотерм (по Бей-Биенко с изменениями): 1 - северная граница отряда таракановых, 2 - январская изотерма -20° на уровне моря, 3 - изотерма средних абсолютных минимумов температуры -25°
У особо теплолюбивых и нехолодостойких видов северная граница ареала может определяться даже сравнительно незначительными зимними температурами. Например, кровяная тля не заходит в Западной Европе и в СССР севернее январской изотермы - минус 3 -4°С; поэтому ее северная граница проходит через Варшаву, далее к востоку спускается к Каменец-Подольску, Херсону, проходит по северным склонам Кавказа и далее идет по северу Средней Азии.
Однако северная граница распространения может определяться не только минимальными зимними температурами, но и количеством тепла за вегетационный период, т. е. суммой эффективных температур. В этих случаях сумма эффективных температур, необходимая для развития одного поколения, может оказаться тем пределом, за который вид не может выйти. Так, например, северная граница ареала стеблевого мотылька в Евразии определяется суммой эффективного тепла в +710°С при пороге 9°С (рис. 186).
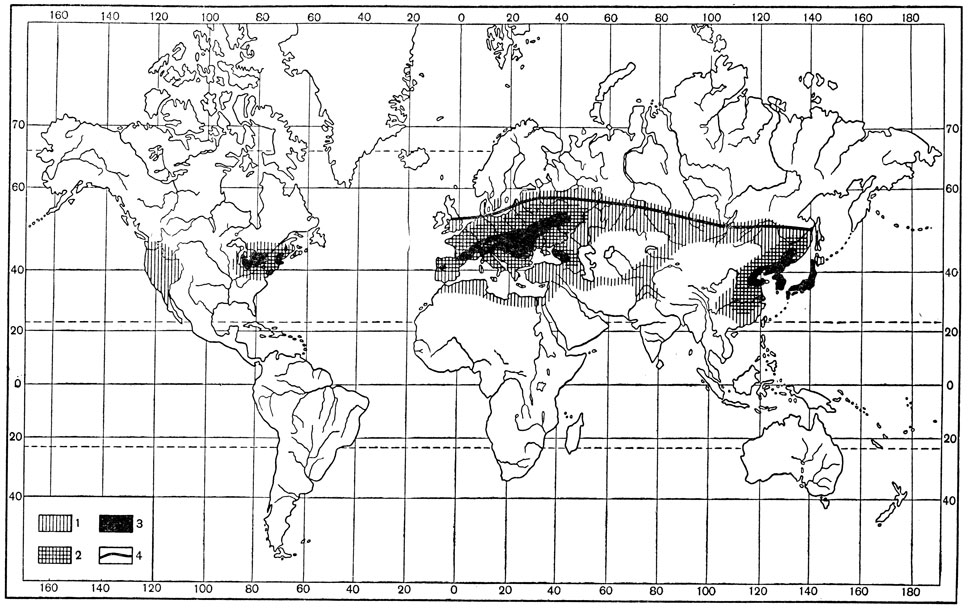
Рис. 186. Карта распространения стеблевого мотылька (по Кожанчикову): 1 - общий ареал, 2 - частая встречаемость, 3 - зона массовых размножений, 4 - изолиния суммы эффектных температур в 710°С
Роль влажности в ограничении распространения также существенна, но не всегда может быть выявлена с должной наглядностью. В отношении вышеупомянутого стеблевого мотылька установлено, что как влаголюбивый вид он в наибольшей численности встречается только в достаточно увлажненных частях Евразии и Северной Америки, совпадающих преимущественно с областями распространения широколиственных лесов. В СССР это северная Украина, средняя полоса Европейской части. Кавказ и Дальний Восток (рис. 180); здесь стеблевой мотылек выступает как вредитель растений. В засушливых степных и пустынных районах этот вид отсутствует или встречается в небольшой численности и как вредитель значения не имеет. Следовательно, во влажных областях умеренного климата создается тот экологический оптимум, который обеспечивает данному виду наибольшее процветание, повышенную численность и высокую вредность.
Примером тесной зависимости ареала вида от особенностей увлажнения является ареал мароккской саранчи (рис. 187). Он охватывает в СССР крайний юг Украины, включая Крым, далее на востоке заходит в Предкавказье, отсюда простирается до Закавказья, юга Средней Азии и юго-восточного Казахстана. Как оказалось, вся эта обширная и разнородная территория отличается особенностями весеннего увлажнения: за три весенних месяца (март, апрель, май) здесь выпадает примерно 100 мм осадков. Поэтому изогиета весенних осадков в 100 мм проходит вдоль всего охарактеризованного выше ареала мароккской саранчи, повторяя все его причудливые очертания.
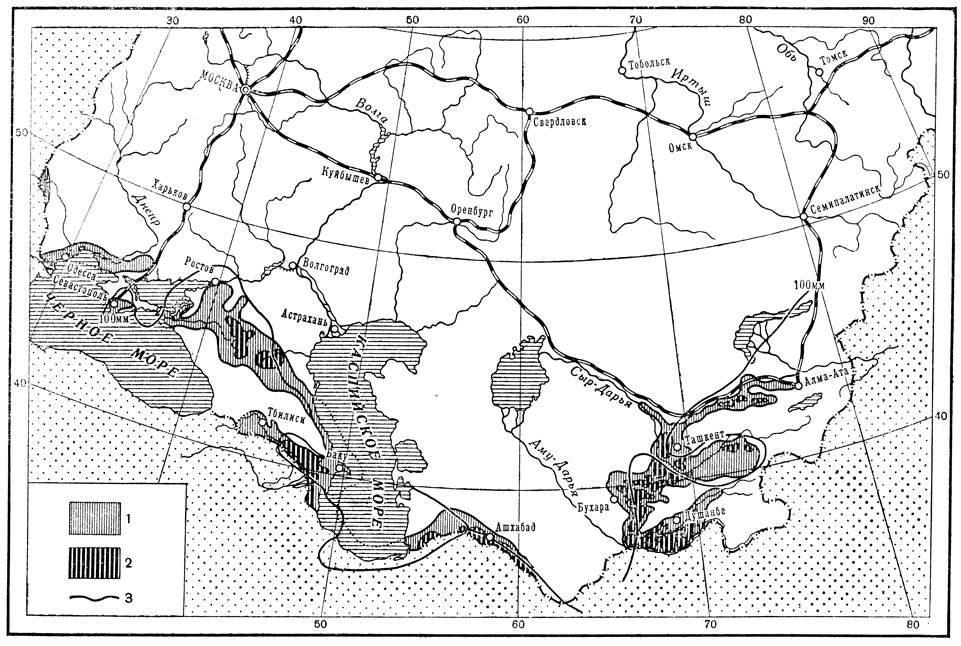
Рис. 187. Карта распространения мароккской саранчи в СССР (по Бей-Биенко): 1 - общий ареал, 2 - прежние и современные очаги массовых размножений, 3 - изогиета весенних осадков в 100 мм
Экологическое значение такой строгой избирательности мароккской саранчи к количеству весенних осадков расшифровывается так. Будучи представителем сухолюбивой средиземноморской фауны, мароккская саранча не терпит излишней весенней влажности, так как ее перезимовавшие в почве яйца при этом гибнут от грибных заболеваний; поэтому значительное увеличение весенних осадков против нормы в 100 мм вызывает резкое снижение численности саранчи. С другой стороны, слишком засушливая весна, когда осадков выпадает значительно меньше 100 мм, создает другую крайность - недостаток влаги; это способствует гибели части яиц от высыхания и гибели отродившейся части личинок от бескормицы вследствие преждевременного выгорания растительности.
В сущности сходный экологический механизм выявляется и при анализе условий существования мароккской саранчи методом биоклимограмм: избыток осадков в фазе развития яйца характеризует неблагоприятные условия существования мароккской саранчи (рис. 177). Однако этот избыток приходится в данной биоклимограмме на зимние месяцы, а не на весенние потому, что она была построена для Малой Азии, т. е. для южной окраины ареала с ее мягкой зимой; зимние осадки здесь в некоторой степени экологически эквивалентны весенним осадкам в более северных частях ареала.
Роль пищевого фактора в распространении насекомых наиболее резко сказывается на узко специализированных видах - олигофагах и особенно монофагах. Так, пшеничный трипс встречается практически всюду, где возделывается пшеница, а репная белянка - всюду, где возделывается капуста; то же самое можно сказать и о яблонной плодожорке - ее ареал совпадает с областью распространения культурной яблони. Несомненно, в широком распространении этих и аналогичных им видов решающую роль сыграло распространение их основного кормового растения.
Однако в большинстве случаев роль пищевого фактора в распространении видов перекрывается другими факторами, в особенности климатическими. Так, капустная белянка и озимая совка отсутствуют в Сибири, хотя здесь широко возделываются их основные кормовые растения: у белянки - капуста, у совки - озимые хлеба. Отсутствие этих видов в Сибири объясняется, как уже указывалось, пониженной их морозостойкостью.
Вместе с тем не возникает сомнений в том, что широкому распространению этих видов в Европе способствовало распространение возделываемых культур. В частности, развитие земледелия на Кольском полуострове и возделывание там капусты способствовало проникновению туда и капустной белянки; по особенностям зимнего периода Кольский полуостров более благоприятен для капустной совки, нежели Сибирь, так как располагается южнее январской изотермы -20°С и чуть севернее изотермы средних абсолютных минимумов температуры -25°С (ср. рис. 185). Очевидно, там, где климатические условия не препятствуют расселению вида, роль кормового фактора становится ведущей.
Своеобразна роль пищевого фактора в распространении завезенных иноземных видов из числа монофагов и олигофагов - виноградной филлоксеры, колорадского жука и др. Их ареал в значительной степени зависит от распространения кормовых растений, в данном случае - виноградной лозы и картофеля, но не охватывает всего ареала этих растений. Объясняется это тем, что дальнейшему расселению этих видов препятствуют в основном меры карантинного контроля, хотя не исключена и ограничивающая роль климатических факторов.
Таким образом, ареал видов определяется комплексным воздействием экологических факторов; при этом ведущая роль может принадлежать в одних случаях либо в той или иной части ареала климатическим факторам, а в других - пищевому фактору. Несомненно значение в распространении видов также и водно-почвенных факторов, но эта сторона вопроса здесь не рассматривается.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'