Половая система и размножение
Все ранее рассмотренные системы органов являются органами индивидуальной жизни, тогда как биологическое назначение половой системы совсем иное - она выполняет функцию размножения и тем самым обеспечивает существование вида. Следовательно, половая система составляет систему органов видовой жизни.
Как правило, насекомые раздельнополы; самцов принято обозначать значком ♂, самок - значком ♀. Половой диморфизм нередко проявляется весьма ярко и заметно по ряду внешних, вторично-половых признаков - по форме и размерам усиков, величине тела, различных деталях строения и пр. Иногда он проявляется особенно резко; например, самец жука-носорога имеет на голове характерный рогообразный вырост, отсутствующий у самки, надкрылья у самца жука-плавунца гладкие, а у самки обычно ребристые (рис. 42) и пр. Существенные отличия между полами могут быть также и в образе жизни и поведении. Например, самцы большинства прямокрылых способны стрекотать, тогда как самки часто лишены этой способности. В целом самцы нередко отличаются большей подвижностью, нежели самки, последние же иногда ведут более скрытый образ жизни, чем самцы. Однако очень часто оба пола внешне неотличимы между собой и тогда их распознавание возможно только по гениталиям.
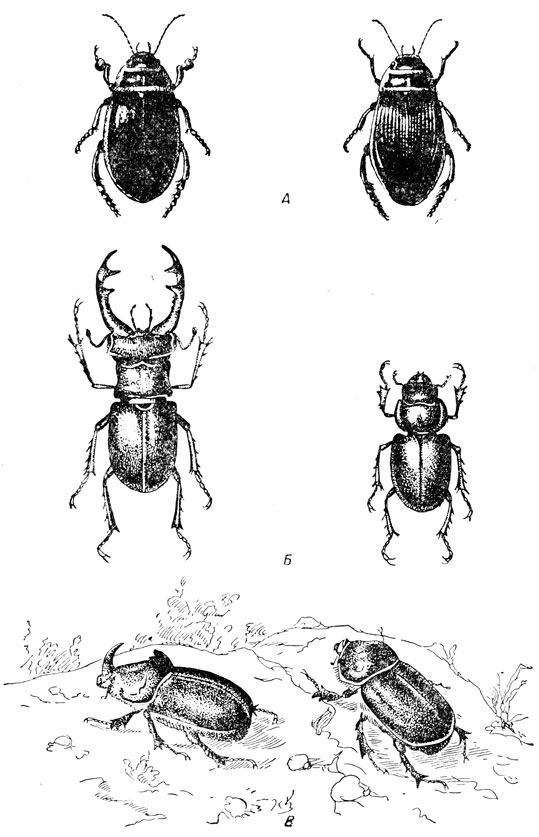
Рис. 42. Половой диморфизм (по Богданову-Катькову и Холодковскому): А - жук-плавунец; Б - жук-олень; В - жук-носорог (слева - самец, справа - самка)
Несмотря на существенные отличия половой системы самца и самки, она в своей основе имеет много общего, а в эмбриональном состоянии практически одинакова у обоих полов. Общий план строения органов размножения таков. Они состоят из парной половой железы, или гонад, пары выводящих протоков, сменяемых перед половым отверстием непарным выводным протоком, придаточных половых желез и половых придатков. Гонады относятся к типу трубчатых желез и состоят из серии трубок, называемых фолликулами.
В своем исходном состоянии половая железа насекомых, видимо, имела два половых отверстия, и образование непарного выводного пути произошло позднее. Остатком такого первичного состояния является половая система таких примитивных насекомых, как поденки (Ephemeroptera), у которых еще сохранились два половых отверстия.
Половое отверстие лежит перед анальным отверстием и в типичных случаях располагается позади IX стернита брюшка или у самки также позади VIII стернита.
Половая система самца. Половая система самца состоит из пары гонад - семенников, пары семяпроводов, непарного семяизвергательного канала, придаточных половых желез (рис. 43) и мужского полового придатка - эдеагуса (см. рис. 18, Б). Семенники обычно имеют вид округлых тел и состоят из фолликулов, число и форма которых неодинакова у разных групп насекомых. В вершинной части фолликула - гермарии - происходит образование живчиков, или сперматозоидов; последние возникают путем многократных делений из первичных семенных элементов - сперматогоний.
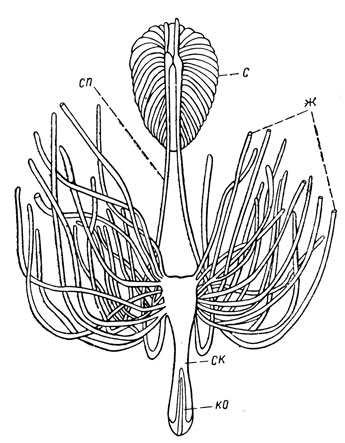
Рис. 43. Половая система самца перелетной саранчи (по Ивановой): с - семенники (слившаяся пара их), сп - семяпроводы, ж - придаточные половые железы, ск - семяизвергательный канал, ко - копулятивный орган
Образовавшиеся сперматозоиды из семенников поступают в семяпроводы, снабженные особым расширением - семенными пузырьками. Отсюда они поступают в семяизвергательный канал, через него достигают эдеагуса и при спаривании с самкой выводятся наружу. Эдеагус, или копулятивный орган, называемый иногда также пенисом, образует наружные гениталии самца. Они нередко имеют сложное строение, характерное не только для семейств, подотрядов и отрядов насекомых, но часто также и для родов и видов; поэтому строение гениталий самца широко используется в современной систематике насекомых.
Придаточные половые железы могут быть в числе от одной до трех пар и открываются своими отверстиями близ основания семяизвергательного канала. У ряда насекомых эти железы служат для образования сперматофоры - мягкой капсулы с порцией сперматозоидов (рис. 57). Такая сперматофора вводится самцом при спаривании в половое отверстие самки или прикрепляется к нему; сперматозоиды затем переходят из сперматофоры в половые пути самки. Сперматофорное оплодотворение свойственно многим насекомым. Его особенности всесторонне изучались В. Ф. Болдыревым у прямокрылых и других насекомых; при этом было установлено, что строение сперматофоры и поведение особей при спаривании разнообразно у разных насекомых и может служить их характерным признаком.
Половая система самки. Половая система самки состоит из пары гонад - яичников, пары яйцеводов, непарного яйцевода, придаточных половых желез, семяприемника и нередко яйцеклада (рис. 44). Яичники составляют основу и наиболее сильно развитую часть половой системы самки. Их фолликулы называются яйцевыми трубками, или овариолами; число яйцевых трубок различно у разных насекомых - от 1-4 пар до 100 и более, а у термитов даже свыше 2400 пар. У самок тлей один яичник может быть редуцированным, а оставшийся имеет всего лишь единственную яйцевую трубку.
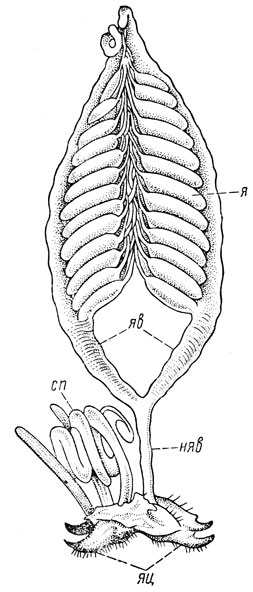
Рис. 44. Половая система самки перелетной саранчи (по Поспелову): я - яичник, яв - яйцеводы, сп - семеприемник, няв - непарный яйцевод, яц - яйцеклад
Каждая яйцевая трубка подразделяется на вершинную часть - гермарий - и основную часть - вителлярий (рис. 45). В гермарии происходит образование и размножение первичных половых клеток - оогоний; из них затем образуются ооциты, а также и питательные клетки. Созревший и выросший ооцит превращается в яйцо и поступает в вителлярий, стенки которого изнутри выстланы фолликулярным эпителием. К гермарию примыкает тонкий тяж - филамент; эти тяжи от разных яйцевых трубок соединены вместе и образуют концевую часть яичника.
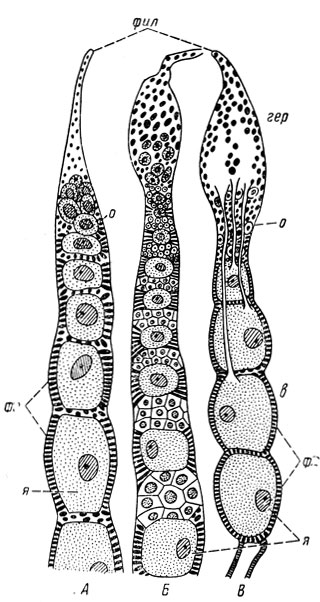
Рис. 45. Типы яйцевых трубочек. А - паноистический; Б - политрофический; В - телотрофический (из Поспелова): фил - филамент, гер - гермарий, о - ооциты, в - вителлярий, ф - фолликулярный эпителий, я - яйца
Находящиеся в вителлярий яйца по мере созревания увеличиваются в размерах и часто отделяются от соседнего яйца заметным сужением, или перехватом; в этих случаях вителлярий оказывается подразделенным на ряд последовательно утолщающихся яйцевых камер.
Рост и развитие ооцитов и яиц происходит за счет поступления питательных веществ. Последние образуются в питательных клетках, а также в фолликулярном эпителии путем поглощения из крови. По окончании развития яйца питание его прекращается, фолликулярный эпителий выделяет хитинообразное вещество и образует наружную оболочку яйца - хорион.
По присутствию или отсутствию питательных клеток и расположению их в яйцевых трубках различают три типа яйцевых трубок (рис. 45). Паноистический тип отличается отсутствием питательных камер, характеризует наиболее примитивное состояние и свойствен в большинстве низшим группам насекомых - таким, как стрекозы, надотряд ортоптероидных (Orthopteroidea), т. е. тараканам, богомоловым, термитам, прямокрылым и др.
Политрофический тип отличается присутствием питательных клеток, чередующихся с яйцевыми клетками; он характерен преимущественно для отрядов насекомых с полным превращением - сетчатокрылых, чешуекрылых, перепончатокрылых, двукрылых и плотоядным жесткокрылых, но свойствен также сеноедам и вшам. Телотрофический тип, называемый также акротрофическим, характеризуется расположением питательных клеток в вершинной части трубки, откуда питательный материал поступает к яйцевым клеткам по протоплазматическим тяжам; он свойствен равнокрылым, клопам и жукам из подотряда разноядных (Polyphaga). Нередко политрофический и телотрофический типы объединяются в один общий тип - мероистический, отличающийся от паноистического присутствием питательных клеток.
Созревшие яйца из яйцевых трубок поступают в парные яйцеводы, затем в непарный яйцевод и оттуда через половое отверстие выходят наружу. В непарный яйцевод впадает тонкий проток семяприемника; он имеет мешковидную форму и служит для хранения сперматозоидов, которые попадают в него при спаривании. Оплодотворение яйца происходит при прохождении его через непарный яйцевод во время яйцекладки; сперматозоиды в это время выходят из семеприемника и проникают в яйцо. Иногда у самок развита совокупительная сумка - мешковидный орган, впадающий также в непарный яйцевод; при копуляции сперматозоиды первоначально попадают в эту сумку, а уж затем - в семяприемник. Но у большинства чешуекрылых совокупительная сумка имеет свое независимое - копулятивное - отверстие; поэтому их половая система имеет два отверстия, из которых яйцекладное ведет в непарный яйцевод и служит только для вывода яйца наружу.
Придаточные железы открывают свой проток также в непарный яйцевод и выполняют разные функции - выделяют секрет для приклеивания яйца к субстрату, для обволакивания группы яиц и образования оотеки (тараканы, богомоловые) или кубышки (саранчовые) и пр. У полового отверстия нередко развит яйцеклад, который может быть наружным и хорошо видимым (прямокрылые, часть перепончатокрылых и др.), либо внутренним и скрытым, либо ложным.
Внешний вид половых желез самки сильно меняется в зависимости от степени созревания яиц. В самом начале развития яиц они еще очень бедны желтком, яйцевые трубки тонки и имеют беловатую окраску; в дальнейшем, по мере созревания происходит накопление желтка в яйцах, увеличивается толщина яйцевых трубок и в ряде случаев появляются перехваты между отдельными яйцевыми камерами. После откладки яиц клетки фолликулярного эпителия дегенерируют и в них появляется желтый пигмент.
Созревание и откладка яиц у кровососущих двукрылых (комаров, слепней и др.) невозможны без принятия порции крови; поэтому до кровососания яйца в яичниках самки остаются недоразвитыми. Но у насосавшейся крови самки яйца начинают быстро развиваться и увеличиваться в размерах; происходит это потому, что принятая кровь дает необходимые питательные вещества, которые быстро усваиваются и отлагаются в яйцевых клетках преимущественно в виде капель желтка. После полного использования принятой порции крови и освобождения от нее кишечника соответствующая порция яиц созревает и при наличии необходимых условий происходит яйцекладка. После нее новое кровососание обеспечит созревание и откладку новой порции яиц и так до 4-5 раз. Это явление получило название гонотрофического цикла и может быть использовано для определения возраста кровососов, что важно при борьбе с ними.
Итак, половая система насекомых имеет у обоих полов общий исходный план строения, но обнаруживает сильный половой диморфизм. Состояние развития половой системы может служить критерием для суждения о возрасте и половой зрелости особей; а особенности строения часто являются важнейшим признаком для различения видов, родов и других систематических групп и широко используются в современной систематике насекомых.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'