Брюшко
Брюшко, или абдомен (abdomen), является третьим отделом тела, состоит из серии более или менее сходных сегментов и у взрослых насекомых лишено ног (рис. 18). Сегменты брюшка, или уромеры (uromeres), устроены значительно проще грудных и состоят из верхнего полукольца - тергита и нижнего полукольца - стернита; верхнее и нижнее полукольца соединены по бокам тела мягкими плейрами. Обособленных твердых плейральных склеритов, как в груди, здесь нет.
Строение брюшка. В своем исходном состоянии брюшко состоит из 11 сегментов и хвостового компонента - тельсона (telson); но в таком виде брюшко сохранилось лишь в отряде безсяжковых (Protura) из первичнобескрылых насекомых (рис. 18, А) и у зародышей. У других насекомых происходит олиномеризация брюшка, число сегментов в нем сокращается сначала до 10, а в высших группах даже до 4-5 (некоторые перепончатокрылые и двукрылые). Сокращение числа сегментов происходит за счет утраты или сильного изменения вследствие смены функции вершинных сегментов, но также и за счет редукции I сегмента; иногда X сегмент остается, но сильно сокращаются или исчезают некоторые предшествующие ему сегменты. Вследствие указанных обстоятельств истинный порядковый номер сегментов далеко не всегда совпадает с фактическим.
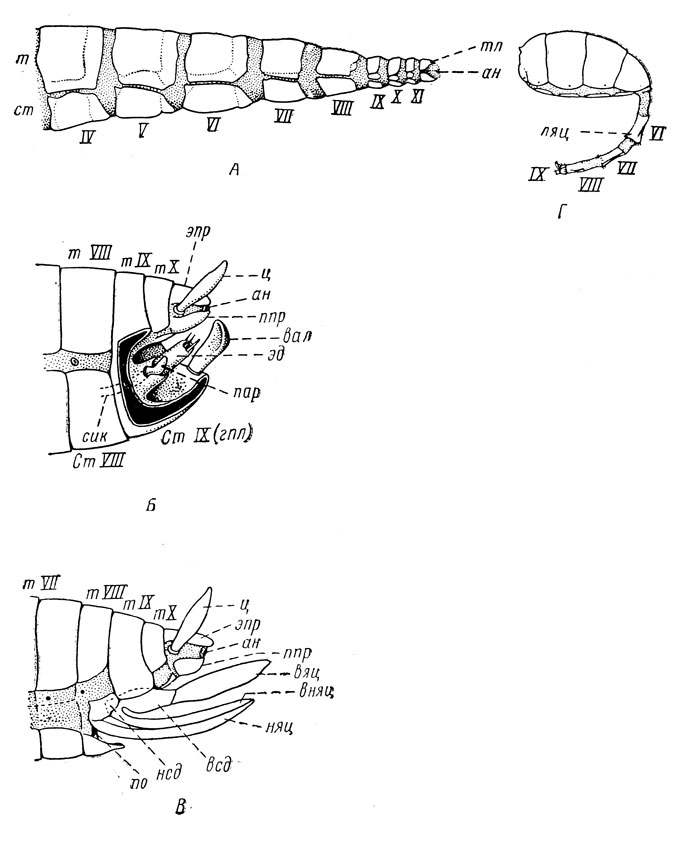
Рис. 18. Строение брюшка и его придатков (по Веберу и Снодграссу): А - часть брюшка безсяжкового Eosentomon, Б - вершина брюшка самца с гениталиями; В - то же у самки с яйцекладом; Г - брюшко комнатной мухи с ложным яйцекладом; т - тергит, ст - стернит, тл - тельсон, ан - анальное отверстие, ц - церк, эпр - эпипрокт, ппр - парапрокт, эд - эдеагус, пар - парамера, вал - вальва, гпл - генитальная пластинка, сик - семяизвергательный канал, ляц - ложный яйцеклад, вяц - верхняя створка яйцеклада, вняц - внутренняя створка яйцекладка, няц - нижняя створка яйцекладка, всд - верхний створкодержатель, нсд - нижний створкодержатель, по - половое отверстие, IV-XI - соответствующие сегменты брюшка
В разделе о строении груди уже указывалось, что I сегмент брюшка стебельчатых перепончатокрылых (пчелы, осы, муравьи и др.) вошел в состав груди, образуя промежуточный сегмент, или проподеум; следовательно, фактически первый сегмент брюшка у таких насекомых в действительности есть II сегмент.
Очень часто в брюшке утрачивается соответствие между числом тергитов и стернитов: первых на 1-2 может быть больше, чем вторых; например, у прямокрылых, тараканов и их родичей I стернит редуцирован, а X или даже VIII - X стерниты утрачены, поэтому число видимых стернитов у них равно 6-8, а тергитов - 8-10. В целом олигомеризация брюшка слабее выражена у низших отрядов и усиливается в высших группах насекомых.
На VIII и IX сегментах брюшка находятся наружные половые придатки, или гениталии; к их числу относится у самца копулятивный орган и у самки ряда насекомых яйцеклад (рис. 18, б, в). В соответствии с этим указанные сегменты называются половыми, или генитальными; предшествующие им I-VII сегменты получили название прегенитальных, а X и XI - постгенитальных.
Прегенитальные сегменты обычно не имеют каких-либо особенностей и не специализованы. Следует лишь отметить, что у стебельчатых перепончатокрылых брюшко отделено от груди тонким стебельком; если этот стебелек длинный (многие осы, муравьи, наездники), такое брюшко называют стебельчатым, если короткий (пчелы, некоторые осы) - висячим. Другие перепончатокрылые, у которых стебелька нет (пилильщики и рогохвосты), имеют сидячее брюшко, как и большинство остальных насекомых.
Постгенитальные сегменты представлены лишь в низших отрядах. Их XI сегмент сокращен в размерах, его тергит лежит над анальным отверстием и образует анальную пластинку, или эпипрокт, а остатки стернита образуют парапрокты - пару пластинок по бокам анального отверстия; X сегмент в этом случае представлен хорошо выраженным тергитом. У таракановых, однако, эпипрокт слит с X тергитом и это комплексное образование также называют анальной пластинкой. На генитальных сегментах особо сильное развитие приобретает VIII или IX стернит, которым нередко и заканчивается брюшко снизу; этот стернит обычно называется генитальной пластинкой. У самца генитальная пластинка образована IX стернитом и нередко обозначается как гипандрий (hypandrium); у самки генитальная пластинка образована VIII стернитом или замещена VII стернитом. В высших отрядах все эти структуры не выражены или сильно преобразованы.
Придатки брюшка. В зародышевом состоянии брюшные сегменты имеют зачатки конечностей, но в последующем развитии насекомого эти зачатки либо исчезают, либо некоторые из них преобразуются в органы с новой функцией. Такими видоизмененными остатками брюшных конечностей являются церки, грифельки, яйцеклад прямокрылых, жало перепончатокрылых и пр.
Следует отметить, что у зародыша хвостовой компонент, или тельсон, лишен зачатков конечностей и, следовательно, не может считаться истинным сегментом; однако иногда тельсон рассматривают как XII сегмент.
Церки представляют собой придатки XI сегмента и при исчезновении последнего перемещаются на X сегмент. Они свойственны многим низшим группам насекомых - таким, как часть первичнобескрылых, древнекрылые и отряды таракановые, прямокрылые и их родичи. У таракановых (Blattoptera), поденок (Ephemeroptera), щетинохвосток (Thysanura) церки имеют вид пары мягких длинных членистых придатков. У прямокрылых (Orthoptera) они обычно одночленисты и играют роль органов, позволяющих самцу удерживать самку при спаривании. У уховерток (Dermaptera) церки превращены в сильные нечленистые клещи - орган защиты и нападения.
Грифельки представляют собой пару небольших удлиненных нечленистых придатков на IX стерните брюшка самца у тараканов, кузнечиков и некоторых других прямокрылообразных (Orthopteroidea). Но в отрядах щетинохвосток (Thysanura) и двухвосток (Diplura) из первичнобескрылых грифельки часто развиты на многих сегментах: с I - II до VII или IX; грифельки здесь играют роль опорных выступов, которые поддерживают брюшко на некотором расстоянии от субстрата и, подобно полозьям салазок, облегчают скольжение тела по поверхности при движении насекомого. У щетинохвосток из семейства махилид (Machilidae) такие грифельки есть и на тазиках средних и задних, ног. В целом присутствие грифельков на брюшных стернитах свидетельствует об одном из ранних этапов утраты брюшком ходильных ног; как оказалось, тазики, или коксы, брюшных конечностей вошли в состав стернитов брюшка, а телоподит полностью исчез. У тараканов и кузнечиков грифельки еще сохранились на IX стерните самца и личинок обоих полов, но у громадного большинства насекомых они полностью исчезли. Из сказанного ясно, что брюшные стерниты являются комплексным образованием, т. е. коксостернитами, и, следовательно, не вполне гомологичны стернитам груди.
Яйцеклад (рис. 18, В) составляет у прямокрылых (Orthoptera), их родичей и некоторых других насекомых гениталии самки и служит для откладки яиц. Он имеет парную природу, хотя внешне часто выглядит непарным, как, например, у кузнечиков. У последних он состоит из пары нижних створок - выростов VIII стернита брюшка и пары разветвленных надвое выростов IX стернита; верхняя пара ветвей образует верхнюю пару створок яйцеклада, а нижняя пара ветвей - внутреннюю пару створок. Такой яйцеклад и его модификации получили название ортоптероидного - по имени отряда прямокрылых, или Orthoptera.
У паразитических перепончатокрылых (Hymenoptera), как наездники, он сохранил ортоптероидную природу и также состоит из трех пар створок, но у жалящих перепончатокрылых, как пчелы, осы и муравьи, он превратился в орган защиты и нападения - жало, снабженное протоком ядовитой железы. У большинства остальных насекомых ортоптероидный яйцеклад утрачен, но потребность в органе со сходной функцией может появиться вновь; однако вследствие необратимости эволюционного процесса (принцип Л. Долло) возникает совсем иной - вторичный, или ложный, яйцеклад, как у некоторых жуков (Coleoptera) и двукрылых (Diptera). Такой вторичный яйцеклад образуется из сильно уменьшенных в диаметре вершинных колец брюшка, вдвигающихся друг в друга, подобно частям телескопа (рис. 18, Г); поэтому он иногда называется телескопическим яйцекладом.
Гениталии самца (рис. 18, Б) представлены весьма разнообразными структурами, но вследствие недостаточной изученности и сложности строения происхождение их частей не всегда выяснено. В общем на конце брюшка самца располагается копулятивный аппарат, служащий для введения при спаривании в половые пути самки сперматозоидов, а также нередко и для удержания самки; входящие в этот аппарат части обычно сильно склеротизованы и нередко образуют сложные структуры. Почти всегда хорошо выражен копулятивный орган, или фаллус (phallus), называемый также эдеагусом (aedeagus); его срединная, обычно непарная склеротизованная часть называется также пенисом (penis), хотя этим термином иногда обозначается и эдеагус в целом. У поденок (Ephemeroptera) и у части уховерток (Dermaptera) пенис, однако, парный. На основании эдеагуса расположена пара боковых лопастевидных образований - парамеры (parameres).
Природа всех этих образований неясна и спорна; одни авторы считают их вторичными, не имеющими отношения к конечностям генитальных сегментов, другие рассматривают их как производное разветвленных парных выростов IX стернита, входящих у самки в состав яйцеклада (см. выше). Большое разнообразие структур копулятивного аппарата самца широко используется в современной систематике для различения видов, родов, семейств и даже отрядов. Нередко внешне одинаковые особи оказываются по структуре гениталий самца или самки хорошо различающимися видами. Очевидно, морфологическая дивергенция признаков при видообразовании у насекомых часто начинается с полового аппарата.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'