Строение крыльев и их типы
Крылья насекомых в типичных случаях представлены двумя парами и являются придатками птероторакса, т. е. средне- и заднегруди. В своей основе крылья представляют собой боковую складку тела и поэтому по происхождению двуслойны, т. е. возникли из верхнего и нижнего слоев. Между последними есть щель и сюда заходит из полости тела кровь, но в целом крыло имеет вид тонкой пластинки, по которой проходят жилки. Жилки представляют собою трубковидное утолщение пластинки крыла и образуют его опорный скелет; помимо крови, внутрь жилок заходят также трахеи и нервы.
По особенностям строения различается несколько типов крыльев (рис. 15). Для сетчатых крыльев характерно присутствие между продольными жилками большого числа поперечных жилок, что особенно свойственно стрекозам (Odonatoptera) и сетчатокрылым (Neuroptera). При небольшом числе поперечных жилок и негустом жилковании крылья называются перепончатыми; они характерны для многих отрядов, например для перепончатокрылых (Hymenoptera), бабочек (Lepidoptera), двукрылых (Diptera) и др. Иногда передняя пара крыльев становится плотной и даже роговой и в покое прикрывает задние крылья; такие крылья, следовательно, приобрели покровную функцию и называются надкрыльями, или элитрами (elytrae). Они свойственны тараканам (Blattoptera), прямокрылым (Orthoptera), жукам (Coleoptera) и др. У представителей отряда полужесткокрылых, или клопов (Hemiptera), передние крылья частью также несут покровную функцию, но участвуют и в полете; вершина крыльев нежная, перепончатая, а остальная часть более плотная, поэтому их называют полунадкрыльями, или полуэлитрами. Из сказанного ясно, сколь большое значение имеет строение крыльев в распознавании и классификации насекомых.
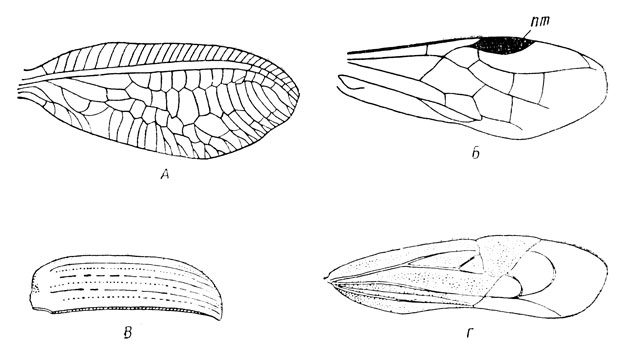
Рис. 15. Типы крыльев (по Веберу и Гуссаковскому): А - сетчатое (переднее крыло золотоглазки); Б - перепончатое (переднее крыло пилильщика, пт - птеростигма); В - надкрылье жука; Г - полунадкрылье клопа
Существенную особенность морфологии крыльев составляют особенности расположения на них жилок, или жилкование (рис. 16); оно складывается из продольных и поперечных жилок. У высших форм поперечные жилки сокращаются в числе или даже совсем исчезают; в таких случаях главную поддерживающую, т. е. механическую роль играют продольные жилки. Этим и объясняется то, что продольные жилки, хотя и претерпели у высших групп значительную эволюцию, все же сохранили свою основу. Продольные жилки являются поочередно выпуклыми и вогнутыми.
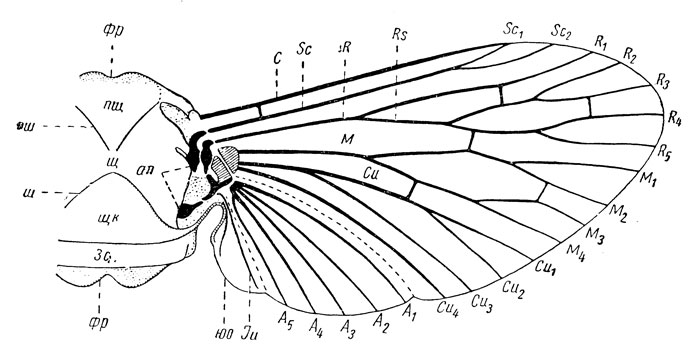
Рис. 16. Схема среднеспинки, прикрепления и жилкования крыла. Обозначение жилок в тексте (по Веберу): фр - передняя и задняя фрагмы, пщ - предщит, щ - щит, щк - щиток, зс - заспинка, пш - парапсидальный шов, ш - v-образный шов, ап - аксиллярные пластинки (показаны черным), юо - югальная область
В исходном состоянии на крыльях развиты следующие продольные жилки (рис. 16): костальная, или коста (costa, сокращенно С); субкостальная, или субкоста (subcosta, Sc); радиальная, или радиус (radius, R); срединная, или медиальная, также медиа (media, M); кубитальная, или кубитус (cubitus, Cu); анальная, или аналис (analis, A).
Однако эти жилки составляют лишь исходную, или основную, схему. В процессе эволюции жилкование подвергается существенным изменениям - упрощению или усложнению. В частности, могут появляться дополнительные ветви на субкосте, радиусе и других жилках; в этих случаях их обозначают порядковой цифрой, например, Sc1, R3 и т. д. Главная задняя ветвь радиуса, если она развита, часто называется радиальным сектором (RS). Кубитальных жилок может быть две - передний (CuA) и задний (CuP) кубитус, анальных - также несколько жилок (A1, A2 и др.). Помимо того, на задних крыльях позади анальных жилок могут быть также югальные жилки (Ju), входящие в состав складывающейся задней части крыльев - веера.
Промежутки между продольными жилками называются полями, которые обозначаются по имени проходящей спереди жилки, а именно: костальное, субкостальное, радиальное, срединное, кубитальное и анальное поля. Заключенные между поперечными жилками части обозначаются как ячейки и также имеют свою номенклатуру по прилегающим продольным жилкам. В некоторых случаях у костального края крыла близ его вершины развито плотное, темноокрашенное пятно - это крыловой глазок, или птеростигма.
Движение крыльев и полет. Насекомые являются первыми на земле животными, выработавшими активный полет; при этом, в отличие от летающих позвоночных, крылья не являются видоизменением конечностей. Следовательно, насекомые за приобретение крыльев не расплатились потерей передней пары ног.
Крыло прикреплено к мембране между спинкой и плейритом; при этом спинка, по выражению Б. Н. Шванвича, "может смещаться вверх и вниз по отношению к плейритам, наподобие того, как крышку сосуда можно приподнимать и опускать над ним". Благодаря этому место прикрепления крыльев к мембране оказывается подвижным. Чуть отступя от своего места прикрепления, крыло лежит на выросте плейрита - столбике. Следовательно, столбик играет роль точки опоры для крыла; раз эта точка опоры располагается слегка отступя от основания крыла, крыло становится двуплечным рычагом с неодинаковой длиной плеч (рис. 12, Б).
Длинное плечо - это гребная часть крыла, лежащая кнаружи от столбика; короткое плечо - это очень небольшой участок крыла у его основания: от места прикрепления к мембране сегмента до столбика. При опускании спинки вниз ее край начинает тянуть вниз основание крыла, т. е. его короткое основное плечо. И хотя это смещение спинки очень мало, его достаточно для поднятия крыла; причина здесь, в том, что плечевой рычаг имеет очень короткое основное плечо.
Здесь же у основания крыла располагается серия мелких склеритов - сочленовных пластинок: несколько аксиллярных и промежуточная (рис. 16). Эти пластинки играют существенную роль в работе крыльев, укрепляют основание крыла между спинкой груди и основанием жилок, обеспечивают гибкость этого сочленения и возможность складывания и расправления крыла. Из этих пластинок существенна роль двух аксиллярных: первая аксиллярная входит в состав короткого плеча крыла, а вторая аксиллярная - лежит на самом столбике. В целом создается совершенная система качания крыла на столбике с амплитудой до 180° между крайними положениями гребного рычага крыла.
Движение крыла при полете отличается большой сложностью, и роль крыльев сводится к двум задачам: удержанию тела в воздухе и движению вперед. В связи с этим крылья насекомых не могут быть уподоблены несущим плоскостям самолета; каждое крыло может рассматриваться как своеобразный пропеллер, создающий движение вперед. Следовательно, полет насекомых является пропеллирующим, что особенно характерно для высших групп - таких, как перепончатокрылые и двукрылые. Например, крыло неподвижно закрепленной пчелы или мухи описывает своим концом сложную траекторию, имеющую вид цифры 8, наклоненной верхним концом назад; при свободном движении вперед эта восьмерка растягивается и превращается в волнообразную кривую - синусоиду (рис. 17).
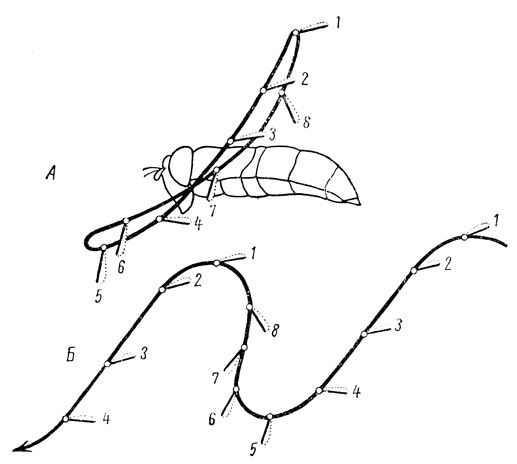
Рис. 17. Схема траектории крыла насекомого (по Залесскому с изменениями): А - при стоячем полете мухи-журчалки; Б - при полете ее вперед; 1-8 - восемь последовательных положений крыла (черная черта - его нижняя сторона, пунктир - верхняя сторона)
Достигнув высшей точки подъема, крыло опускается вниз в виде горизонтальной плоскости, поэтому создается подъемная сила, эффект работы крыла становится элеваторным. Но при опускании крыло одновременно отводится косо вперед и близ нижнего положения начинает поворачиваться вдоль своей продольной оси; в самой нижней точке оно ставится на ребро, т. е. его плоскость оказывается вертикальной. В таком состоянии оно отводится вверх и назад, чем создается движение вперед; эффект работы крыла становится пропеллирующим. Большая частота взмахов обеспечивает слияние элеваторного и пропеллирующего эффекта в единый аэродинамический эффект, что и позволяет насекомому лететь вперед и держаться в воздухе.
Одной из самых совершенных форм полета является так называемый стоячий полет, характерный для многих мух; насекомое "неподвижно" держится в воздухе в одной точке. Достигается это тем, что снимается пропеллирующий эффект - путем опускания крыльев отвесно вниз, т. е. без отведения их косо вперед.
Совершенный полет требует слияния элеваторного и пропеллирующего эффекта; это обеспечивается повышением частоты взмахов крыльями. Некоторые насекомые в этом отношении достигают поразительных показателей; например, крыло мухи совершает 300 колебаний в секунду, у настоящих комаров - 500-600 колебаний, а у мельчайших комаров-мокрецов - даже до 1000. Однако дневные бабочки совершают всего лишь 5-10 взмахов в секунду.
Скорость полета составляет у шмеля 18 км в час, у бабочки-бражника - 54 км, а у стрекозы - до 96 км; однако если отнести эту скорость к длине тела, то она будет более чем в 5-6 раз превышать относительную скорость современного самолета.
Особым приспособлением к нормальной работе крыльев при высоком ритме их работы или при большой их величине является образование на них птеростигмы; она есть у стрекоз, сетчатокрылых, перепончатокрылых и др. Оказалось, что птеростигма устраняет вредную вибрацию крыльев, которая в аэродинамике обозначается термином фляттер. Устранение фляттера у современных скоростных самолетов достигнуто аналогичным способом - установкой на переднем крае у конца крыла особого утяжеления. Совершенствование полета насекомых сопровождается также перемещением продольных жилок к переднему, или костальному, краю крыла; такая костализация жилкования способствует укреплению переднего края крыльев, улучшает их аэродинамические показатели и отличает высшие семейства и подотряды насекомых от низших.
Взлет насекомых происходит у разных представителей по-разному - в зависимости от величины тела, частоты взмахов крыльев, воздействия внешней среды. Многие используют для взлета прыжок; таковы саранчовые, кузнечики и другие прямокрылые, цикады и листоблошки из равнокрылых, земляные блошки из жуков. При большой площади крыла и низком ритме взмахов эффект взлета может быть достигнут от одного взмаха, например у дневных бабочек. Но при высоком ритме взмахов необходимо приспособление для подготовки к включению крыльев сразу на полную частоту, иначе подъемный эффект не будет достигнут. Таким включающим и стимулирующим аппаратом у двукрылых являются жужжальца - сильно измененные рудименты задних крыльев в виде тонкой, вздутой на конце палочки. Они способны колебаться с большой скоростью - свыше 300 качаний в секунду, что и обеспечивает соответствующую настройку нервной системы и включение в работу крыльев. Но, помимо того, жужжальца в полете играют и роль своеобразного жироскопа; их маятникообразные колебания на высокой частоте стабилизируют равновесие насекомого в полете.
Стимуляционное действие может оказывать и такой внешний фактор, как ветер; установлено, что у саранчи на голове есть особые группы волосков, которые стимулируют полет, если на них направлен ток воздуха.
Эволюция полета и крыльев. Как показал А. В. Мартынов, крылатые насекомые подразделяются на две группы: одни держат крылья в покое распростертыми в стороны или отгибают назад и не способны их компактно укладывать вдоль тела, другие приобрели способность плотно складывать крылья вдоль тела, и благодаря этому такие насекомые в покое имеют компактное, обтекаемое тело. Первые обозначены как древнекрылые насекомые (Palaeoptera), из них в настоящее время сохранились лишь два отряда - поденки (Ephemeroptera) и стрекозы (Odonatoptera), но в палеозойскую эпоху были и другие отряды, нацело вымершие; вторые названы новокрылыми (Neoptera), к ним относятся все остальные многочисленные отряды крылатых насекомых.
Способность складывать крылья вдоль тела возникла у новокрылых благодаря появлению в их крыльях новообразования - югальной области; это особая подгибающаяся складка, расположенная у основания обеих пар крыльев сзади, позади анальных жилок (рис. 16). Появление этой югальной области, а вместе с ней и способности складывать крылья вдоль тела, оказалось весьма крупным прогрессивным преобразованием крылатых насекомых, сделало их тело обтекаемым и способным использовать новые убежища для жизни - под растительными остатками, под камнями, в толще травостоя, в различных щелях и пр. Все это открыло перед насекомыми совершенно новые возможности для бурного расцвета и прогрессивной эволюции и поставило их на более высокую ступень развития; в целом возникновение новокрылых явилось таким коренным эволюционным преобразованием, которые акад. А. Н. Северцов обозначал в теории эволюции понятием ароморфоз.
Существенной стороной в эволюции полета и в изменениях крыльев оказалось и то обстоятельство, какую роль в полете играют та и другая пары крыльев. Как показал Б. Н. Шванвич, по участию в полете крыльев и их крыловой мускулатуры, или крылового мотора, различаются три группы насекомых: бимоторные, т. е. использующие при полете в одинаковой степени обе пары крыльев; переднемоторные - использующие преимущественно или исключительно переднюю пару крыльев, и заднемоторные - использующие в полете задние крылья.
При характеристике строения и типов крыльев уже указывалось, что у некоторых насекомых передние крылья приобретают покровную функцию и превращаются в надкрылья. В связи с этим летательная функция целиком или в значительной мере переходит к задним крыльям, усиливается их мускулатура и такие насекомые становятся заднемоторными. К их числу относятся отряды прямокрылых, жуков и их родичи. В сущности, у заднемоторных возникает функционально двукрылый полет.
Бимоторное состояние характерно для стрекоз и некоторых низших сетчатокрылых; в целом оно характеризует примитивные группы или примитивное состояние полета. Зато широко распространена переднемоторность. Она сопровождается объединением в работе передних и задних крыльев; между ними возникают различные оценочные приспособления, которые приводят в конце концов также к функциональной двукрылости: обе пары действуют уже как одна пара. Благодаря этому основная нагрузка при полете переходит к передней паре крыльев, возникает переднемоторное состояние. Все это ведет постепенно к ослаблению задней пары крыльев, и, наконец, к их исчезновению: функциональная двукрылость переходит в морфологическую, что свойственно отряду двукрылых (Diptera).
Таким образом, развитие и совершенствование крыльев насекомых шло в направлении выработки двукрылого полета; в одних случаях это достигалось путем выключения из полета передних крыльев, в других - путем ослабления или атрофии задних крыльев. На это явление впервые обратили внимание Б. Н. Шванвич и Б. Б. Родендорф; последний обозначил его как диптеризацию. Отсюда этот процесс развития полета был назван как принцип диптеризации, или принцип Родендорфа - Шванвича.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'