15.6. Высшие пчелы
Трибы Meliponini и Apini удобно рассматривать вместе. Trigona fuscipennis и Т. fulviventris - пчелы одинаковой величины; обе они питаются пыльцой на кустах Cassia, вскрывая пыльники (Johnson, Hubbell, 1974, 1975). На прибрежной равнине Коста-Рики эти растения цветут во время сухого сезона. Т. fuscipennis обычно фуражирует группами на кустах, растущих также группами на расстоянии в среднем 1,5 м от ближайшего соседнего куста, тогда как Т. fulviventris посещает либо одиночные растения, либо редко разбросанные цветки на кустах, растущих группами при среднем расстоянии 8,1 м от ближайшего соседа. Это различие частично обусловлено характером взаимоотношений между двумя видами: Т. fulviventris избегает Т. fuscipennis, которая демонстрирует свою агрессивность, "зависая" в воздухе на расстоянии 1 см от рабочих особей Т. fulviventris, пока те не улетят. Т. fulviventris вообще не подлетает даже к незанятым гроздьям цветков, хотя неизвестно, научились ли пчелы избегать этих цветков в результате прежнего опыта или их предупреждает пахучая метка, оставленная Т. fuscipennis. У Т. fuscipennis имеется видоспецифичный феромон, и она подлетает к цветкам группами, которые, возможно, ведет разведчица; других насекомых эти пчелы прогоняют, и их плотность достигает 22 особей на 1 м2. В отличие от этого Т. fulviventris не пытается вступать в конфликты с другими насекомыми, и ее плотность составляет всего лишь 3 особи на 1 м2. Короче говоря, Т. fuscipennis - агрессивный групповой фуражир, расходующий энергию на то, чтобы монополизировать и удерживать за собой хорошие грозди цветков, тогда как Т. fulviventris фуражирует в одиночку и затрачивает энергию на поиски новых цветков; обе тактики оказываются успешными.
Джонсон и Хаббелл (Hubbell, Johnson, 1978) ставили сахарные приманки, привлекавшие до 9 видов пчел. В этом эксперименте Trygona fulviventris из нескольких разных семей, встречаясь в центре приманки, вступали в схватку, но ограничивались тем, что прогоняли друг друга при помощи угроз и толчков головой. Позднее Т. fuscipennis возвращались большой группой под предводительством разведчицы и монополизировали несколько приманок. Крупная черная пчела Т. sylvestriana без труда прогоняла Т. fulviventris своим жужжанием, но не очень эффективно использовала завоеванные таким образом приманки. Т. testaceicornis применяла совершенно иную тактику: она пробиралась к занятым приманкам и кормилась вместе с другими пчелами, улетая только в том случае, если ее обнаруживали; однако ее численность могла достигать такой величины, что позволяла ей монополизировать приманки. Таким образом, и у Trigona мы находим различные способы, применяемые в ситуациях, связанных с конкуренцией: прямое нападение, пустые угрозы, настойчивость и мобилизация подкрепления или же попытки незаметно "втереться" - с уходом в случае неудачи. Джонсон и Хаббелл считают, что эти стратегии выработались в результате сосуществования и что они позволяют свести к минимуму потери энергии при конкурентных взаимодействиях.
Превосходные примеры межвидовых экологических различий можно найти и в роде Apis. Два вида - A. mellifera и А. сеrаnа - таксономически очень близки, и до того как человек стал заниматься пчеловодством, они распространялись соответственно на запад и на восток из южной части Азии, откуда они, вероятно, происходят. Это не просто географические подвиды, так как имеются бесспорные данные об отсутствии скрещиваний между ними (Ruttner et al, 1972, 1973). Дело в том, что их самцы собираются в одних и тех же местах, привлекаемые одним и тем же феромоном (9-ОДК), который образуется в мандибулярных железах репродуктивных самок всех видов Apis. Самцы A. mellifera, находясь на расстоянии в несколько километров, препятствуют осеменению самок А. сеrаnа их самцами. И это несмотря на то, что трутни А. сеrаnа летают быстрее и при более низких температурах. Возможно, что более крупный пакет спермы самцов A. mellifera, у которых сперма более жидкая, чем у А. сеrаnа, разрушает пакет последнего вида после копуляции, хотя это мало вероятно, так как в Пакистане присутствие в данном районе А. сеrаnа блокирует нормальную копуляцию A. mellifera (Koeniger, Wijayagunasekera, 1976)!
В Японии Apis сеrаnа - местный вид, но A. mellifera была интродуцирована сюда человеком, поскольку она запасает больше меда и реже роится. Взаимоотношения между этими видами описал Sakagami (1959, 1960; см. также Brian, 1965). У А. сеrаnа мощные жвалы, при помощи которых она может расширять разного рода полости, увеличивая тем самым диапазон полостей, пригодных для устройства гнезда; возможно, что это способствует более успешной зимовке. Однако A. mellifera быстрее обнаруживает врагов и жалит их, чем А. сеrаnа; она может отгонять А. сеrаnа, не давая ей возможности жалить в ответ; она быстрее мобилизует сборщиц и направляет их к блюдцам с сиропом, а также устраивает групповые набеги на семьи А. сеrаnа. Очевидно, A. mellifera способна к более быстрой передаче информации и к более эффективной групповой активности. У пчел А. сеrаnа существует, однако, некая уклончивая тактика: они улетают, когда гнездо подвергается нападению, а после ухода врагов возвращаются назад. Это спасает семью от полной гибели при нападении хищного шершня (Vespa mandarinia); в аналогичной ситуации A. mellifera оказывает сопротивление и нередко бывает уничтожена. Таким образом, экологическое равновесие между этими двумя видами Apis могло бы поддерживаться хищным шершнем, который поедает A. mellifera, и грабителем-человеком, который выращивает их. Во всяком случае, А. сеrаnа, по-видимому, лучше приспособлена к жизни в естественных условиях в горах.
В Пакистане Apis сеrаnа встречается с A. florea и A. dorsata в горах, которыми ограничено здесь ее распространение (Ruttner el al., 1972). В Шри Ланке она сосуществует с этими же видами, а также с Trigona iridipennis (Meliponini) (Koeniger, Vorwohl, 1979). Очевидно, на залежах, на которых после их кратковременного использования восстанавливается растительность, пища имеется в изобилии, особенно в связи с тем, что в засушливые летние месяцы и A. dorsata, и A. florea улетают в другие места. При конкуренции за искусственный корм взаимоотношения между особями складываются не всегда одинаковым образом, т. е. у них нет четко выраженной иерархической системы; однако в общем и целом Trigona iridipennis доминирует над А. сеrаnа и A. florea, но никогда не сталкивается с A. dorsata; A. florea доминирует над А. сегапа, а последняя - над A. dorsata. В среднем наблюдаются взаимоотношения Т. iridipennis > A. florea > A. cerana > A. dorsata, тогда как по размерам тела эти виды расположатся в обратном порядке. Авторы отмечают, что при таком положении вещей самый мелкий вид, не способный летать далеко, получает возможность фуражировать поблизости (примерно в 100 м) от гнезда без затруднений, тогда как виды, способные летать на большие расстояния, например A. dorsata (5 км), фуражируют в отдаленных местах, улучшая таким образом распределение видов в пространстве. Только у А. сеrаnа наблюдаются конфликты между семьями. Все эти виды строят гнезда в совершенно различных местах: полости, используемые маленькой пчелой Trigona iridipennis, меньше тех, в которых устраивается А. сеrаnа, а у двух других видов Apis гнезда открытые - A. florea строит гнезда вокруг маленьких веточек, а у A. dorsata они свисают с больших ветвей. Хотя у всех видов Apis репродуктивные самки используют в качестве полового аттрактанта 9-ОДК, запах которого привлекает всех трутней, гибридизация отсутствует, так как у разных видов брачный лёт происходит в разное время дня (рис. 15.4): у A. florea в 12-15, у А. сеrаnа в 16-18, а у A. dorsata в 18-19 часов.
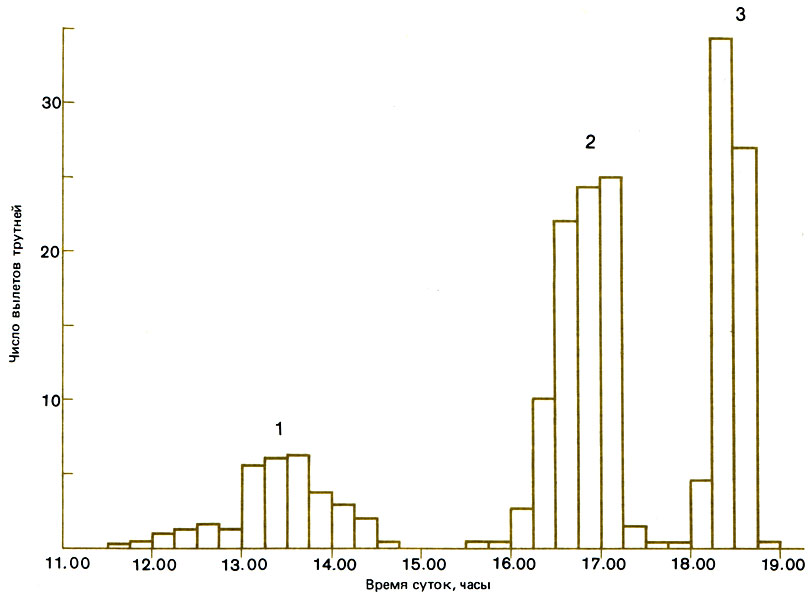
Рис. 15.4. Различия во времени лёта трутней у трех видов Apis: A. florea (7), A. cerana (2) и A. dorsata (3). (Koeniger, Wijayagunasekera, 1976.)
Медоносная пчела (Apis mellifera) - с помощью человека или независимо от него - распространилась по большей части земного шара и образовала много подвидов, которые в настоящее время скрещиваются между собой. Существует несколько африканских подвидов (Fletcher, 1978b). Например, в южной части пустыни Сахара в саванне, поросшей колючим кустарником и высокой травой, живет А. m. adansonii, а в Центральной Африке сходное местообитание, но с примесью вечнозеленых и листопадных тропических деревьев занимает другой подвид, A. m. scutellata. Первый из этих подвидов в 1956 г. был интродуцирован в Бразилию, где он вступил в конкуренцию с европейской A. mellifera, завезенной раньше, а к 1971 г. эта "африканизированная медоносная пчела" распространилась почти по всей стране (Michener, 1975).
Особенности этих подвидов состоят в том, что у них рабочие особи и ячейки мельче; гнезда они устраивают в любых местах, защищенных от непогоды, например в гнездах термитов, расщелинах скал или зданий, в ящиках, под деревьями и даже в почве, что делает их весьма гибкими в выборе местообитаний. Поведение, связанное с зимовкой, у них отсутствует; у них не наблюдается снижения активности, а образование клуба бывает кратковременным; когда корма становится мало, они легко перебираются в другие места. Обе эти черты указывают на приспособленность скорее к тропическому, чем к умеренному климату. Это активные трудолюбивые фуражиры, быстро перелетающие с цветка на цветок. Их система мобилизации, указывающая направление к корму даже при расстоянии всего лишь 50 м, а иногда и 10 м, отличается от аналогичной системы А. m. ligustica - главного европейского подвида - более низкой частотой виляний на единицу расстояния. Они тоже собирают и запасают большие количества меда. Их защитные действия хорошо координированы, что придает реакции массовость, и они успешно отгоняют врагов, хотя отдельные уколы жала у них не сильнее, чем у обычных пчел. Короче говоря, они представляют собой эффективный тропический вариант типичной Apis mellifera.
Замещая в Бразилии европейскую пчелу, описанные подвиды выработали ряд приспособлений. Одно из них состоит в том, что благодаря своей способности устраивать гнезда в несколько необычных местах они сумели создать обширную дикую популяцию. Это позволяет им непосредственно вытеснять семьи Apis mellifera путем грабежа и нападения, а их трутням дает больше шансов на успешную гибридизацию. Гибриды сочетают в себе признаки обеих родительских рас: они используют стандартные ульи и вощину, они активные фуражиры, хорошо воспринимающие указания о местонахождении корма, но вместе с тем они легко снимаются с места и очень чувствительны к нарушению их покоя и к агрессии.
Как показывает этот обзор экологии видов, относящихся к одному роду, существует множество способов, при помощи которых виды избегают взаимодействия в случае недостатка пищи или мест для гнезда. У них могут быть разные периоды сезонной и суточной активности, что обычно зависит от различия в критической температуре. Благодаря разнице в размерах тела отдельные виды могут либо собирать более крупные кусочки пищи, либо уходить дальше от гнезда на ее поиски. Они могут устраивать гнезда или кормиться на разных уровнях сильно структурированных биомов, в результате чего нередко потребляют различную пищу. Гибридизация между близкими видами может снижаться благодаря тому, что брачный лёт происходит у них в разное время суток или года. Некоторые виды, по-видимому, подразделяются на моно- и полигинные формы - по своей экологии соответственно первооткрыватели и завоеватели. Есть и такие виды, которые мигрируют в другие географические районы, чтобы избежать конкуренции в неблагоприятное время года. В главе 16 эти группы видов, относящихся к одному роду, рассматриваются в контексте тех сообществ, в состав которых они входят, чтобы можно было представить себе их взаимодействия в более широком плане.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'