10.3. Взрыв численности
При достаточном количестве рабочих особей и, если это необходимо, солдат и при наличии найденных и охраняемых источников пищи семья может быстро наращивать число рабочих. Хотя, казалось бы, и так ясно, что чем больше рабочих, тем больше проделанная работа, это нетрудно проверить экспериментально и установить численное соотношение между этими двумя величинами. У Myrmica rubra одна самка, если предоставлять ей все возрастающее число рабочих особей, увеличивает число откладываемых яиц, но только до известного предела, который достигается всего лишь при 20 рабочих (Brian, 1969). Поскольку число рабочих, приходящихся на одну самку, у этого полигинного вида равно в среднем 76 (Elmes, 1973), остается еще много рабочих для ухода за личинками, и нет оснований думать, что самка не может продолжать откладывать максимальное число яиц, когда большинство рабочих особей (скажем, две трети) ухаживают за расплодом. Чем меньше рабочих приходится на одну самку, тем меньше яиц откладывает в среднем каждая из них (Brian, Rigby, 1978); некоторые самки, возможно, страдают от конкуренции за пищу и могут даже не выдержать этой конкуренции. Оптимальное число самок, при котором выход яиц достигает максимума, а неблагоприятные взаимодействия минимальны, с увеличением числа рабочих особей возрастает, но оптимальное число самок при данной группе рабочих, безусловно, не соответствует оптимальному для этой группы числу личинок, которых они должны кормить. Эти проблемы требуют экспериментального изучения и анализа путем численного моделирования.
Можно показать, что численность рабочих особей имеет также важное значение для выживания яиц. Эксперименты, в которых группам рабочих Myrmica ruginodis числом от 3 до 40 давали по 50 яиц, показали, что число вылупляющихся личинок сначала возрастает с увеличением числа рабочих, а затем, достигнув максимума (около 12), начинает снижаться (Brian, 1953); на самом деле для получения 11 личинок нужно всего 20 кормилиц. Учитывая эффективность группы в целом и индивидуальную эффективность рабочих особей, можно считать, что наиболее экономными были бы группы по 11 рабочих, способные вырастить 7-8 личинок. Такие же эксперименты можно провести для более поздних стадий развития. Так, группам рабочих особей разной численности - от 5 до 320-давали по 50 личинок третьего возраста сразу после зимовки; оказалось, что для получения максимального роста и максимального выхода куколок при очень малой смертности достаточно 80 рабочих особей. Поскольку с увеличением численности группы вклад каждой особи снижается, наиболее экономным вариантом будут группы по 42 рабочих, которые обеспечат примерно две трети максимального выхода. При этом оптимальное соотношение рабочих и личинок составит 0,84, а это практически равно тому, что было найдено в диких семьях, собранных только за один сезон, - 0,88. Это позволяет думать, что гнезда в природе функционируют в отношении выращивания расплода на уровне, близком к оптимальному. А из этого, конечно, следует, что экономичному функционированию благоприятствует естественный отбор, а не то, что рабочие особи всегда способны эффективно выполнять свои функции.
В гораздо большей выборке семей, относящихся к разным видам Myrmica и имеющих разную численность, отношение рабочие/личинки составляло в среднем 0,85 при 2000 рабочих особей (Elmes, Wardlaw, 1981). У всех семи изученных видов наблюдалась сходная зависимость между числом рабочих и числом личинок, но она не была линейной: при 1100 рабочих отношение рабочие/личинки было равно 1, при 1000 рабочих-1,14, а при 100 рабочих достигало 3. Таким образом, с увеличением числа рабочих это отношение, вместо того чтобы возрастать или хотя бы оставаться постоянным (как следовало бы ожидать, если бы с повышением плотности эффективность падала), на самом деле слегка уменьшается. Это может означать, что более крупные семьи эффективнее в отношении выращивания расплода, чем мелкие. Эффективность может снижаться из-за дублирования, избыточности, нарушений в передаче информации, столкновений или просто попыток избежать их. В условиях сильной скученности, когда, например, большое число рабочих особей пытается добраться до нескольких личинок, подаваемые личинками сигналы могут просто не дойти до рабочих, находящихся в отдалении, и те будут спокойно ожидать, оставаясь на краю группы. Это хорошо, так как при этом уменьшается группа суетящихся рабочих, которые стремятся обслужить личинок.
Поскольку рабочие особи выращивают рабочих, т. е. создается положительная обратная связь, приводящая к экспоненциальному росту, все еще трудно представить себе, сколь динамична популяция общественных насекомых на самом деле. Некоторые примеры могут помочь нам в этом. Матка пчелы Evylaeus marginatus откладывает в первый год только шесть яиц, а позднее, когда ей уже помогают рабочие, она за 5 лет своей жизни производит больше 1000 яиц; рабочие особи живут не более года (Plateaux-Quenu, 1962). У Bombus pascuorum откладка яиц сначала идет медленно, но затем ускоряется, достигая максимума - 12 яиц в день; у этих шмелей точное соответствие между приростом числа яиц и приростом взрослых особей достигается путем откладки яиц на коконы, что обеспечивает в среднем соотношение 3 яйца : 1 кокон (A. D. Brian, 1951, 1952). Данные об этой содержавшейся в помещении семье В. pascuorum (и одной семье В. humilis), обитатели которой могли свободно вылетать наружу, показывают, что личинки развиваются из 71% яиц, 73% этих личинок окукливаются и из 90% куколок выходят взрослые особи. Это дает вогнутую кривую выживания: оно описывается экспоненциальной функцией с отрицательным показателем при постоянной смертности 4% в день, средней продолжительности жизни 25 дней и периоде полужизни 17,5 дня. Таким образом, простая модель с приростом 12 яиц в день дает 4 новые особи в день и может обеспечить постоянную численность семьи в 100 шмелей, уравновешиваемую гибелью насекомых со скоростью 4% в день (Brian, 1965). Для живущего в тропиках В. morio, у которого фуражиры крупнее и старше, средняя продолжительность жизни кормилиц 73 дня, а фуражиров - только 36 дней (рис. 10.3) (Garofalo, 1978). Кривая выживания кормилиц имеет полунормальную форму, а кривая для фуражиров - вогнутая, но те и другие менее подвержены случайностям, чем у европейских видов шмелей. У Bombus atratus, обитающего в субтропиках Южной Америки, с расплодом дело обстоит примерно так же (Sakagami et al., 1967b), однако сведений о выживании взрослых особей не приводится.
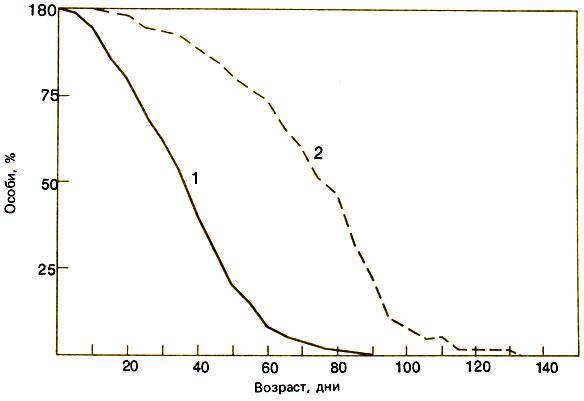
Рис. 10.3. Кривые выживания фуражиров (7) и гнездовых шмелей (2) Bombus morio. (Garofalo, 1978.)
У Polistes необходимость создания основы для ячеек ограничивает откладку яиц до двух в день - всяком случае, до тех пор, пока соотношение рабочие/личинки высокое, как это бывает после появления на свет первой партии рабочих особей. Первостепенное значение, несомненно, придается выращиванию личинок, так что при низких отношениях рабочие/личинки строительство ячеек и откладка яиц снижаются, несмотря на то что недавно освободившиеся ячейки используются повторно. Данные о диких семьях Dolichovespula sylvestris и Vespula vulgaris были использованы для построения модели, описывающей историю развития средней семьи (Archer, 1981а). Каждая маленькая ячейка довольно долго (около 45-50 дней) бывает заполнена расплодом. У D. sylvestris строительство ячеек продолжается непрерывно в течение 40 дней; число их достигает примерно 300, прежде чем освобождаются первые ячейки, которые можно использовать вновь. У V. vulgaris ячейки строятся на протяжении 80 дней, число их достигает 7000, а затем строительство прекращается. Эти цифры показывают, что у обоих видов одна рабочая особь строит чуть больше одной ячейки в день; просто у V. vulgaris это продолжается дольше, а поскольку число рабочих растет в геометрической прогрессии, вся система ячеек оказывается гораздо больше. У обоих видов матки способны обеспечивать все ячейки расплодом, хотя матка D. sylvestris откладывает только по 13 яиц в день, а у V. vulgaris она начинает с 34 яиц в день и доходит до 348. Это относится только к семьям, не достигшим полной зрелости; в зрелой семье D. sylvestris с большими маточниками матка может дойти до 65 яиц в день. Вероятно, даже в очень большом гнезде она не в состоянии достигнуть такого уровня, как матка V. vulgaris, хотя, конечно, мы этого не знаем.
Максимальная численность рабочих у Dolichovespula sylvestris равна 175, a у Vespula vulgaris - 2200, но в общей сложности у первого вида выводится 330 рабочих, а у второго - 10293. Фактически средний возраст рабочих особей в момент их гибели составляет всего 10,3 дня у V. vulgaris (исключая ювенильные стадии) и около 14 дней у D. sylvestris, но о последнем виде данных мало; у шершней (Vespa) рабочие особи живут в среднем около 5 недель. Кривая зависимости смертности от возраста у Vespula vulgaris асимметрична, с модой 9 дней и медианой между 8-м и 9-м днями, что позволяет объяснять значительную часть смертности разного рода случайностями при фуражировке. Период, когда насекомые находятся в гнезде и ухаживают за расплодом, очень короткий или отсутствует вовсе. Стационарная численность взрослых рабочих, возможная для модели Vespula vulgaris с 10%-ной смертностью в день, зависит от числа мелких ячеек, от продолжительности периода развития от яйца до взрослой особи и от выживания расплода при обилии корма. Если, предположим, имеется 5000 подходящих ячеек для рабочих особей, а период развития равен 25 дням, то ежедневный прирост рабочих составит 200 особей, и если они живут в среднем 10 дней, то стационарная численность их должна составлять 2000. В случае V. vulgaris такая численность существует только три недели - с 74-го по 95-й день. При размножении роением 10%-ная смертность - это очень много, но при очень большом числе ячеек и потреблении достаточного количества пищи число рабочих может поддерживаться на высоком уровне.
У Apis mellifera матка в зависимости от расы может откладывать до 2000 яиц в день; если в соте не хватает места, то она их просто бросает и их поедают рабочие особи. Число откладываемых яиц достигает пика в первый и второй годы жизни, а затем снижается; кроме того, наблюдается сезонный пик весной. Матка лишь в редких случаях располагает условиями, необходимыми для полного проявления своих возможностей: зимой, когда на сотах много свободного места, ее лимитирует то, что пчелы могут обеспечить теплом лишь небольшой участок сота, а летом, когда тепла достаточно, ее сначала ограничивает скорость построения ячеек, а позднее - пространство, занятое запечатанным расплодом, развивающимся из ранее отложенных яиц. Таким образом, так же как и у ос, пчелиную матку ограничивает в основном число рабочих особей, выступающих в роли источников тепла, воска или пищи, и скорость роста семьи зависит в первую очередь от этих факторов, до тех пор пока соотношение рабочие/личинки не падает слишком низко. Матка может отложить в "рабочие" ячейки некоторое количество неоплодотворенных яиц; они должны быть уничтожены вместе с оплодотворенными, но нежизнеспособными яйцами, и доля неразвивающихся яиц может возрасти до 50%, хотя обычно она составляет меньше трети. На стадиях личинки и куколки, так же как и у ос, смертность незначительна, если только нет заболеваний и хищников (например, клещей); в хорошем улье в слоях запечатанных ячеек с расплодом пустых мест бывает мало. Из 100 яиц могут вылупиться 94 личинки, 86 из них окукливаются и со временем дают 85 рабочих особей (Fukuda, Sakagami, 1968).
У медоносной пчелы рабочие особи живут значительно дольше, чем у ос, и кривая их выживания имеет "выпуклую" форму; смертность постепенно повышается с возрастом, достигая у фуражиров примерно 3%. Она зависит также от времени года: в марте продолжительность жизни фуражиров составляет 5 недель, в июне - 4 недели, а затем вновь увеличивается с появлением сонных зимующих пчел, которые могут прожить всю зиму, особенно если они поздно появились на свет (Free, Spencer-Booth, 1959). Осенью, как и у большинства других насекомых, интенсивность метаболизма у пчел понижена и они склонны к запасанию различных веществ в жировом теле (Maurizio, 1946). Если их удается побудить к выращиванию потомства, их выживаемость несколько снижается, но они обычно вовремя прекращают заботу о расплоде, так что зимой в улье остаются одни только имаго. По мнению Avitabile (1978), подготовка к зиме начинается вскоре после летнего солнцестояния вследствие чувствительности рабочих особей к укорочению дня, а удлинение дня после зимнего солнцестояния, возможно, побуждает к возобновлению выращивания расплода. Социальный и физиологический механизм этих реакций неизвестен (Kefuss, 1978). Одна из африканских рас медоносной пчелы, Apis mellifera adansonii, которую разводят в Бразилии, где летние температуры близки к 24°С, а зимние - к 18°С, не впадает в спячку; у этой расы нет особей, способных к зимовке, так что их средняя продолжительность жизни равна 26,3 дня летом и 24,3 дня зимой (Terada et al., 1975).
В стационарной семье, состоящей примерно из 60 000 пчел (сильная семья в июне) с общей смертностью 3% (1800 особей) в день матка должна откладывать (при выживаемости 70%) 2600 яиц в день, что близко к пределу возможного в хороших условиях. Модель такой системы, созданная McLellan et al. (1980), предусматривает график откладки яиц, при котором число их быстро возрастает, достигая пика в мае-июне, а затем постепенно снижается, доходя в октябре до нуля (кривая из семейства кривых, соответствующих уравнению у = хе-х). Авторы исходили из следующих данных: 1-1,2 тысячи яиц в день; продолжительность развития до имаго в среднем 20 дней; 10%-ная смертность среди молоди; время жизни имаго 30 дней летом и 200 дней зимой. Эта модель хорошо согласуется с тем, что известно об истории жизни семьи медоносной пчелы, и легко приложима к другим многолетним общественным насекомым, так как форма кривой биомассы достаточно универсальна (ср. кривую для Myrmica у Brian, 1965).
У Apis mellifera adansonii в Южной Америке цикл роения продолжается примерно два месяца (Winston, 1979). Через 20 дней после роения появляются новые рабочие особи и снижение численности семьи прекращается; спустя еще 20 дней число пчел начинает резко расти, и образуется рой. Как указывает Winston, семьи роятся в период быстрого роста населения в улье, а в своем резюме он отмечает: "...Рои возникают перед высокими пиками скоростей роста..." Шестьдесят процентов рабочих особей, особенно молодых, уходят со старой маткой; за 20 дней до роения новые рабочие особи составляют 40-60% имаго. После этого с молодыми самками может уйти еще до четырех роев, но их может и не быть вовсе; число дополнительных роев пропорционально объему запечатанного расплода. После того как рой покинул улей, выживание расплода снижается до 50%, но после появления молодых пчел оно вновь повышается, хотя семье приходится ждать еще 20 дней, прежде чем у нее появится новая плодущая матка. Этот двухмесячный цикл может быть причиной августовского вторичного пика частоты роения, наблюдаемого в областях с умеренным климатом (см. ниже).
Для Meliponini с их ясно выраженной внутригнездовой фазой выкармливания расплода характерно такое же увеличение смертности с возрастом, как и у медоносной пчелы. Garofalo (1978), изучавший Nannotrigona postica и Plebeia drorуana, нашел, что у последнего вида средняя продолжительность жизни на 42 дня больше, чем у Apis mellifera (рис. 10.4). Ни у одной безжалой пчелы, насколько известно, не бывает особых долгоживущих зимних пчел.
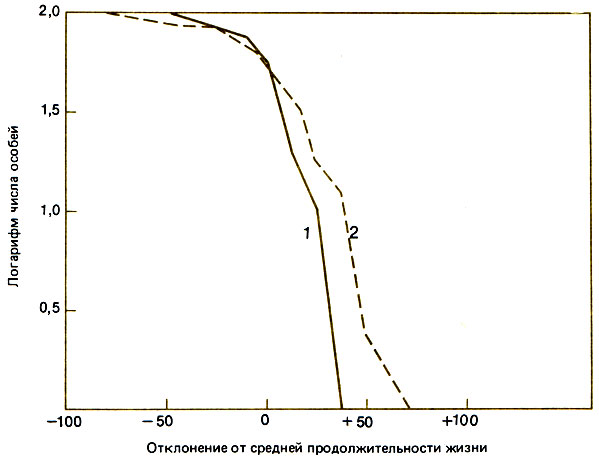
Рис. 10.4. Сравнение кривых выживания Nannotrigona postica (7) и Apis mellifera (2). (Garofalo, 1978.)
Для низших термитов характерна малая плодовитость, порядка 2-300 яиц в год у Calotermitidae и Termopsidae, но у Macrotermes число яиц, откладываемых царицей, достигает астрономических величин: 30-40 тысяч в день (табл. 10.1; другие данные см. Brian, 1965). О выживании молоди сведений мало; определить его трудно, так как молодые особи обычно не остаются на одном месте и часто линяют (Nielsen, Josens, 1978). В популяциях M. bellicosus доля личинок снижается с 60-64% общей численности до 28% в семьях старше 10 лет (Collins, 1981а). На протяжении 5 лет после достижения численности в 10 тысяч особей семьи растут в геометрической прогрессии, а затем в результате дальнейшего роста число особей может превысить миллион. Естественный годовой прирост составляет 78%, т. е. того же порядка, что и у Neotermes tectonae, у которого он равен 106% (Brian, 1965).
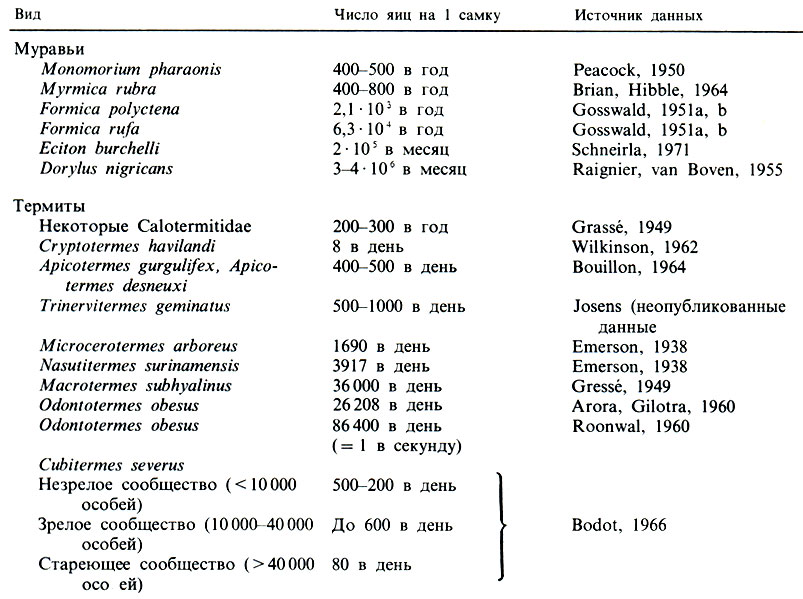
Таблица 10.1. Число яиц, откладываемых самками некоторых видов муравьев и термитов (Brian, 1978)
У муравьев диапазон столь же широк: самка Amblyopone pallipes откладывает всего несколько яиц, тогда как у Eciton burchelli самка, по имеющимся оценкам, дает каждые пять недель 1,2-105 яиц, т. е. в среднем по 3500 яиц в день. Число яиц у маток Myrmica rubra составляет от 410 до 826 в год (рис. 10.5). Рекорд, по-видимому, принадлежит обитающему в индо-малайской области термиту Odontotermes obesus: одно яйцо в секунду! К сожалению, данными о выживании и продолжительности жизни мы пока не располагаем.
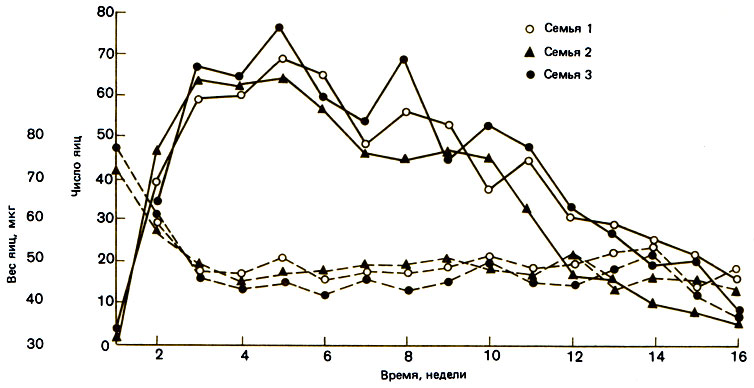
Рис. 10.5. Еженедельная продукция яиц и вес яиц (мкг) самок из трех семей iMyrmica/i rubra. Каждая кривая построена по данным о средней продуктивности четырех самок. Яйца, предрасположенные к развитию в рабочих особей, начинают появляться в период пика продуктивности, между 2-й и 3-й неделями, а затем их число медленно сходит на нет. (Brian, Hibble, 1964.)
Несколько простых моделей семей общественных насекомых, растущих по экспоненте, описал Brian (1965, р. 40). Во всех группах таких насекомых есть виды с большими семьями: из многих тысяч особей у ос и пчел, из миллионов особей у муравьев и термитов. Такие сообщества обладают большей мощностью и стабильностью, однако с увеличением возможностей сбора корма должна возрастать и их плодовитость (или число половых особей). Повышение плодовитости достигается за счет кормления самки-основательницы очищенной концентрированной пищей, за счет увеличения числа и удлинения яйцевых трубочек в каждом ее яичнике (и расширения брюшка, чтобы они могли в нем поместиться), а также развития ее эндокринной системы, в особенности corpora allata, которые снабжают яичники неотонином. У многих муравьев число самок может достигать нескольких сотен; как правило, наблюдается именно это, а не увеличение продолжительности жизни рабочих особей. Фактически время их жизни обычно сокращается; например, у Myrmecia оно составляет 2 года (Haskins, Haskins, 1980), а у Monomorium pharaonis - примерно 7 недель. У последнего вида может быть более 100 самок, откладывающих по 1,5 яйца в день каждая, и если хотя бы одна особь из трех доживает до взрослого состояния, как у Myrmica, то это уравновешивает потерю двух процентов особей в день, так что каждая самка способна поддерживать стационарную популяцию в 25 рабочих особей (Peacock, Baxter, 1950; цит. по Brian, 1965). При смертности 2% в день средняя продолжительность жизни - величина, обратная смертности, - составляет 50 дней. Это не связано с тем, что М. pharaonis - тропический вид; ведь Lasius alienus живет в среднем несколько недель (Nielsen, 1972). И все же по мере расширения внутригнездовых функций становится возможным увеличить также выживание молодых рабочих особей при сокращении жизни старых фуражиров. В чем преимущество высокой скорости обновления? Главным образом в том, что рабочая особь может специализироваться на кратковременном, эфемерном источнике пищи. Для пчел это цветки какого-то одного вида растений; для ос - определенный вид жертвы; для муравьев - возможно, семена какого-то одного растения, а для термитов - особый тип подстилки или вид злаков. Рабочих можно вырастить, получить от них то, что положено, когда они выработают нужный навык, а потом избавиться от них. Они могут не обладать способностью к последовательному освоению нескольких навыков, подобно своим более простым эволюционным прототипам, которые живут дольше и, обладая большей гибкостью, непрерывно учатся и переучиваются. Возможно, что упрощение функций позволяет стандартизировать и упрощать морфологию рабочих особей, но пока точных данных об этом очень мало.
В этой главе были рассмотрены стадии превращения корма в биомассу семьи. После того как рабочие особи берут себе пищу, доставляющую энергию, личинки и репродуктивные особи (у перепончатокрылых это самки) получают корм, необходимый для роста. Молодые личинки и взрослые особи, откладывающие яйца, получают наиболее очищенный и концентрированный пищевой материал, а личинки старшего возраста - смесь или просто непереработанный корм. Возможно, что крупные размеры помогают привлечь внимание кормилиц, так как корм распределяется неслучайным образом. В главе 11 будет описано, как под влиянием половых особей некоторые личинки получают ограниченное количество корма и поэтому из них развиваются мелкие рабочие особи. Подчеркивается важность конкуренции между самками и личинками и ее значение для стабильности сообщества, поскольку эта конкуренция обеспечивает соответствие между количеством эмбрионов и количеством корма; вырисовываются три альтернативных способа повышения общей плодовитости: 1) увеличение яичников у одной самки; 2) увеличение числа самок и 3) откладка рабочими особями яиц, дающих самцов, в период, когда продуктивность самки понижена. Семьи могут возникать в результате расщепления старой семьи, их могут основывать пары половых особей (у термитов) или осемененные самки - в одиночку или небольшими группами. В небольших новых семьях откладка яиц не составляет проблемы, но возможны разного рода помехи и нападения со стороны хищников при фуражировке, и в смысле защитных функций групповое существование дает преимущество. Одиночные самки хорошо заметны, и риск подвергнуться нападению до того, как они начинают создавать семью, для них больше, чем после этого. Несколько заточённых в гнезде самок могут откладывать яйца в один общий фонд и использовать их для кормления вылупляющихся личинок; небольшие группы от этого выигрывают, но если группа слишком велика, то она подвержена заболеваниям. После того как семья создана, численность ее возрастает, но точно так же возрастают и ее организационные проблемы.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'