Кружева ареалов
Итак, каждый вид на основе биотического потенциала и расселительных способностей, особенно в период возникновения волн жизни, оказывает непрерывное давление на окружающую среду и как бы стремится к наибольшему ее захвату. Среда оказывает сопротивление виду и ограничивает его распространение или делает это распространение прерывистым.
Есть виды, которые имеют почти планетарное распространение (конечно, по определенным типам местообитаний), но их немного. Чаще всего они связаны с хозяйственной деятельностью человека (их называют синантропными или, если связь с человеком не так строга, - антропофильными). Подавляющее большинство видов встречается лишь в определенной области, ограниченной контуром, своего рода линией равновесия между потенциалом расселения и силой сопротивления среды. Эта линия не постоянно фиксирована. В годы особенно высоких волн жизни вид может выплескиваться за ее пределы и даже существовать на новой территории год-другой, но затем исчезает, не сумев закрепить там свое потомство. В такой полосе "штормового наката" вид иногда наносит существенный, хотя и периодический, вред сельскохозяйственным культурам, тем более, что появление его часто бывает совершенно неожиданным. Только тщательное наблюдение за колебанием численности вида внутри ареала подскажет человеку сроки массовых размножений насекомых и выхода их за пределы обычных границ распространения.
Однако как ни штормит море, контур его можно нанести на карту, как ни изменчивы границы ареала вида, все-таки их можно изобразить графически и изучить рисунок. А это изучение не только интересно, но и полезло для практики сельского хозяйства.
Ареалы некоторых видов вытянуты вдоль границ природных зон - степей, тайги, пустыни и т. д. Виды, в пределах этих границ распространенные по всей зоне, называют трансзональными.
Представим себе, что севернее границы ареала степного вида вырубили леса и осушили площади, а в полупустыне к югу от южной границы провели орошение. Если вид распространен на большом протяжении степной зоны и мирится с разными условиями степной среды, то он может довольно легко продвинуться как на север, так и на юг, став там новым, а потому особенно опасным вредителем.
Множество видов обитает на относительно небольшом отрезке той или иной природной зоны или на небольших отрезках соседних зон. Такие виды очень разборчиво относятся к характеру грунтов, удаленности от влажного дыхания Мирового океана, составу трав и т. д., менее "легки на подъем", чем виды трансзональные, однако бывают случаи, когда смена растительности вызывает вспышку массовых размножений саранчовых именно с такими локальными ареалами.
Уничтожение пестрых растительных сообществ, замена их посевами часто вызывает активизацию потенциальных вредителей из мира саранчовых, которые до этого находились на своих исходных позициях, сдерживаемые сопротивлением среды. Человек невольно провоцирует активизацию биотического потенциала, словно опускает курок ружья, и помогает виду из потенциального стать реальным вредителем.
Из сказанного ясно, что знание границ ареалов позволит предугадать, где, когда и почему будет спущен этот курок. Иными словами, мы можем прогнозировать вспышку размножений вредителя при определенном изменении природной среды в результате хозяйственных мероприятий в конкретных природных зонах или сельскохозяйственных районах. Вот в чем состоит практическое значение зоогеографии, в частности географии саранчовых, для сельского хозяйства.
В природе встречаются виды с очень ограниченными, можно сказать, миниатюрными, ареалами, распространяющимися лишь на один хребет или долину, или даже одну вершину, котловину, пещеру и т. д. Такие виды животных и растений, характерные для горных стран с их особенной пестротой условий жизни, называются эндемичными. Строго эндемичные виды не угрожают сельскому хозяйству, зато оно само может нанести им непоправимый ущерб. Даже частичное сокращение ареала таит угрозу для вида, так как разреженность популяции, уменьшение соотношения самцов и самок вызывает снижение возможности перекомбинации генов, приводит к мутациям.
Заботясь о сохранении природы, человек всегда должен знать перед началом освоения какого-либо района, есть ли в нем эндемичные виды и как хозяйственная деятельность людей может на них отразиться.
Мы уже говорили о том, что границы географических ареалов насекомых представляют собой как бы линии равновесия между давлением жизни и сопротивлением ограничивающих факторов среды. Однако не везде это так. Граница может возникнуть как результат взаимодействия вида с факторами геологического прошлого. Нужно сказать, что эволюция вида и изменения его ареала идут столь неторопливо, что ледниковые щиты четвертичного периода или разливы морей третичного запечатлеваются видом как совсем недавнее прошлое. Но бывает, что вид, словно очнувшись и обнаружив смену геологических эпох, вдруг резко начинает расселяться.
Приведем пример, касающийся сибирских видов саранчовых. На юге средней и восточной части Сибири широко распространен хортиппус интермедиус. До 40-х годов текущего столетия исследователи обнаруживали его во многих точках к востоку от Енисея. Призвав на помощь палеогеографию, ученые установили, что хортиппусы обитают только на восточном берегу огромного пресноводного озера-моря, некогда занимавшего всю Западную Сибирь. Оно было образовано в результате подпруживания Уральско-Таймырским ледником вод, стекавших на север с Алтая.
Через четверть века после первых сообщений мы нашли этот вид в Новосибирской и в степях Омской областей. Заселение дна давным-давно исчезнувшего моря, видимо, продолжается на наших глазах, хотя началось миллионы лет тому назад.
Другой пример. Представьте себе, читатель, что вы - в экспедиции в Туве, вблизи географического центра ,Азии, на максимальном удалении от влажного летом и теплого зимой дыхания океана. Многое здесь напоминает пустыни Средней Азии, нет только ночного пения сверчков. Оно и понятно: днем жара, а к ночи участникам экспедиции придется надеть ватники и теплые шапки. Зимой же здесь нередки пятидесятиградусные морозы. Где уж выжить сверчкам - выходцам из тропических стран - в таком суровом климате. И вдруг у речки слышатся дружные ночные хоры. Тщательные поиски под камнями - ив руках несколько десятков крохотных удивительных существ из рода птеронемобиус (Pteronemobius). Откуда они здесь? Ведь виды этого рода характерны для Средиземноморья и Тихоокеанского побережья Дальнего востока, то есть сугубо приморских климатических районов. Правда, один вид был обнаружен на территории СССР, только на самом юге Дальнего Востока - у озера Ханка.
Для зоогеографа очевидно, что еще в доледниковое время по широкой полосе субтропических лесов, протянувшихся вдоль берегов Тетиса, этот вид был распространен от запада до востока Евразии. Но вот Тетис отступил. Резко возросла континентальность климата, наступило оледенение. Конечно, виды этого рода на большей части ареала исчезли, остались только в приморских областях. Исчезли, но кое-где, как бы на сохранившихся участках древнего моря, уцелели. А не поменяли ли они свой генофонд в такой изоляции, без обмена наследственными задатками с основной популяцией? Очень хотелось бы подметить момент видообразования, да еще измерить его скорость (мы ведь знаем, сколько времени прошло от начала исчезновения Тетиса). Но, увы, самые тщательные исследования, выполненные авторами вместе с Г. Я. Бей-Биенко, показали: "никаких отличий в строении". С точки зрения систематики вид не изменился. Вот как, оказывается, медленно течет, эволюционное время.
Примером могут служить и близкие родственники саранчовых - кузнечики. Насекомые из обширной группы крупных кузнечиков, связанных с кустарниками Средиземноморья и субтропиков, из Европы проникли в Азию вплоть до Тянь-Шаня. Здесь ареал прерывается, а дальше их обнаруживают только на Дальнем Востоке. И вот в 30-х годах Г. Я. Бей-Биенко находит такого кузнечика на Алтае, на Семинском перевале, покрытом горно-тундровой растительностью. Оказалось, что это новый, еще неизвестный вид, приспособившийся к необычным для него условиям Алтая. Справедливости ради надо заметить, что и этот кузнечик, и тот сверчок, о котором мы писали выше, позже были обнаружены еще в нескольких крошечных убежищах на территории Алтая.
Часто же эволюционные события развиваются быстрее. Оледенения, разливы морей, расширения пустынь разрывают ареал вида на две удаленные одна от другой части, и в этих осколках ареала вид начинает изменяться, превращаясь в два подвида или в еще близкие, но уже разные виды. Таковы, например, прусы и хортиппусы.
Пожалуй, наиболее интересна в этом плане история сибирской кобылки (Gomphocerus sibiricus). Сейчас она распространена по всей Евразии от 65-го градуса северной широты до 35-го градуса южной широты в полосе шириной 3 тысячи километров, захватывающей несколько природных зон. Кобылки постепенно исчезают в степях и пустынях и нигде не подходят по равнине вплотную к горам. Однако в ледниковый период, когда с севера подступали массивы движущегося льда, а тундровые и лесотундровые ландшафты отодвигались к югу, сибирская кобылка обитала в предгорьях. Но вот льды отступили, на месте тундр и лесов образовались степи, и кобылка ушла на север. Однако часть ее популяций предпочла дальним северным землям гораздо более близкие влажные участки по краям горных ледников Тянь-Шаня и Кавказа. Осталась там кобылка, как на островке ледникового периода, дав начало двум современным подвидам - кавказскому и тянь-шаньскому. "Возраст" их известен: примерно один миллион лет. Посмотрим, что же за это время успела сделать эволюция и насколько отличаются два "новых" подвида от основного.
При сравнении пяти главных признаков, в том числе формы переднеспинки, кубитального поля крыла и пропорций усиков у самцов и самок всех трех подвидов (сибирский - основной, кавказский и тянь-шаньский), оказалось, что признаки существенно различаются, особенно длина усиков и пропорции их срединных члеников. Математические методы помогли выявить степень родства этих подвидов: сибирский стоит ближе к тянь-шаньскому и дальше от кавказского. Если все три подвида расположить в ряд по степени сходства, то этот ряд начнет основной подвид - сибирский, среднее положение займет тянь-шаньский, а закончит ряд кавказский подвид.
Итак, изменение признаков дает некоторое представление о скорости эволюции.
Иногда для спасения живого населения при сокращении ареала достаточно даже крохотного островка с благоприятными условиями- среды. Таким островком для сибирской кобылки стала гора Богдо - соляной купол, одиноко возвышающийся, как сахарная голова, среди необозримых пустынных равнин Прикаспия. Когда-то она была островом среди разлившегося после таяния ледников Каспийского моря. И вот на вершине этой горы на площади не более 1,5-2 квадратных километров, куда попадает чуть больше осадков, чем на равнину, сохранилась совсем маленькая популяция сибирской кобылки. Интересно, к какому подвиду она принадлежит; к основному, широко распространенному севернее на равнинах, или к горному кавказскому? А может быть, это еще один подвид или особый вид? Неплохо бы звучало его латинское название - богдензис. Но не будем гадать, подождем, пока компетентный в решении таких вопросов специалист посетит гору Богдо.
Внутренняя гладь ареала, по образному выражению советского эколога К. В. Арнольди, подобна кружеву. Действительно, в природе не бывает так, чтобы в пределах ареала, например липы, это дерево в равной мере занимало бы и равнины, и сухие каменистые горные склоны, и болота, а, например, тростник рос бы не только у воды, но и на скалах, и в лесу. Значит, каждому виду внутри его географического ареала свойствен определенный и довольно узкий круг конкретных мест обитания в определенных ландшафтах. Они-то и образуют внутреннее кружево ареала. Свойство вида образовывать внутри ареала узор (такое же четкое, как, например, размеры или цвет насекомого) Г. Я. Бей-Биенко назвал ландшафтной, или стациальной, верностью вида. Он же доказал, что верность вида строго закономерным образом меняется от одной части ареала к другой, особенно в том случае, когда ареал включает в себя северные и южные природные зоны. Здесь проявляется зональная смена стаций саранчовых, которую Г. Я. Бей-Биенко хорошо изобразил графически. Что же это такое - смена стаций?
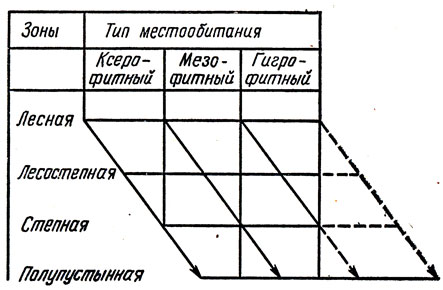
Схема смены стаций (по Бей-Биенко)
Понаблюдаем за коньком (Omocestus haemorrhoidalis). У северного предела географического ареала конек находится в условиях прохладного климата с избыточным увлажнением, поэтому он обитает только на сухих, хорошо прогреваемых песчаных холмах. У южного предела ареала, в жарком климате, приютом ему служат самые влажные места, например заболоченные приречные луга. По-видимому, и на севере в самых сухих местах, и на юге в самых влажных этот конек сохраняет для себя одну и ту же норму тепла и влаги.
Таким образом, смена местообитания саранчовыми выступает как средство сохранения постоянства условий среды.
Как же вообще плетется кружево поселений вида на всем северо-южном разрезе ареала? Для того чтобы выяснить это, нужно обследовать население вида (образно говоря, изучить кружево по отдельным его кусочкам) во многих местах ареала от севера до юга, в довольно длинном ряду ландшафтов от рек и болот до сухих междуречий. Причем во всех точках придется не просто отмечать наличие вида, но и определять степень густоты населенности, то есть проводить количественный учет насекомых на единице площади*.
*(Исследование ареалов многих десятков видов было выполнено авторами в различных географических зонах: по берегам Иртыша, на Алтае, в Туве, Южном и Западном Казахстане, Киргизии, на Кавказе, Урале и в Восточной Сибири.)
Изучение заселенности разных районов страны многими видами позволяет нам теперь рассказать о законах, по которым сплетено кружево ареала любого вида. Сразу же обращаем внимание читателя на практическое значение этих законов, в частности, для своевременного прогнозирования массовых размножений и для организации мероприятий по ограничению численности видов. Как уже было сказано, они временами могут становиться вредными для сельского хозяйства в. определенных резко ограниченных местах (там, где условия им "подыгрывают").
Оказывается, вблизи центра ареала каждого вида встречается некая оптимальная область. Она представляет собой относительно широкую полосу, идущую по одной из природных зон, входящих в ареал. В этой полосе климатические условия для вида наилучшие, поэтому он весьма многочислен. Нет здесь для него и непреодолимых преград (за исключением густого леса, если речь идет о саранчовых), поэтому особи вида в различных ландшафтах общаются между собой и используют один разнообразный генетический фонд.
Можно сказать, что в оптимальной области ареала существует одна крупная популяция вида. Естественно, что здесь по сравнению с другими частями ареала вид играет наиболее заметную роль в круговороте элементов и, конечно, может приносить ощутимый вред сельскохозяйственным культурам. Понятно, что за данным видом нужно вести непрерывные наблюдения. Только в этом случае появление волн жизни можно обнаружить своевременно, что очень важно, так как волны будут распространяться стремительно, охватывая все ландшафты и угодья. В оптимальной области географического ареала саранчовых в любой момент могут понадобиться ядохимикаты, но все же на первом плане должны стоять меры, предотвращающие размножение вредителя.
Из сказанного ясно, что в первую очередь здесь нужно позаботиться о том, чтобы большая общая популяция оказалась разбитой на множество малых, разделенных непреодолимыми и необычными для условий данного района преградами. Какими? Да хотя бы чересполосицей. Особенно нужна чересполосица поедаемых и непоедаемых сельскохозяйственных культур. Сама природа учит нас такому искусству землепользования, таящему в себе и много других выгод.
Полосная система земледелия применяется на целинных землях, о чем рассказал в своей работе о прусе в Павлодарской области агроном кандидат сельскохозяйственных наук Л. Д. Бунин. Материалы, приведенные исследователем, показывают, что чередование широких полос многолетних трав и зерновых дает возможность защитить почвы от ветровой эрозии и восстановить их плодородие. На полосах трав быстро нарастает численность пруса, и он может начать наступление на зерновые, но если и зерновые будут чередоваться (например, пшеница с просом), то прус не прорвется через сеяные барьеры, популяция окажется разобщенной и волны жизни не разгуляются по степным просторам.
Что же происходит с популяцией, удаленной от центра ареала? К северу и к югу от него и, вообще говоря, на окраинах ареала вид не всегда находит удобные для себя местообитания. Интересно, что принцип выбора условий жизни у саранчовых, распространенных к северу или к югу от центра ареала, независимо от вида и географического района страны один и тот же. Во всех случаях сплошная популяция как бы раздваивается, образуя разобщенные рукава: один из них тянется по долинам рек, другой - по водоразделам. Понятно, что в южной части население вида стремится к воде, на севере же прибежище виду в долинах рек дают хорошо прогреваемые песчаные гривы, намытые рекой поверх ее поймы.
Другой рукав популяции тянется по водоразделам, то есть по Таким ландшафтам, через которые вода стекает в долины. Но здесь насекомым хватает, видимо, и атмосферной влаги. Это объясняет расселение вида по таким рукавам (путям) к северу от центральной области его ареала. А как же "чувствуют себя" саранчовые в южных частях ареала, облюбовавшие водоразделы с явным недостатком влаги? Не так уж плохо. Большинство водоразделов плоские, вода с них стекает медленно, поэтому даже в маленьких понижениях на водоразделах образуются очаги застойного увлажнения. Более того, в степных районах в таких местах происходят просадки почвы и возникают степные блюдца с весенними озерками. Вот по этим-то блюдцам (от одного к другому) вид и продвигается к югу.
Оба рукава популяции надежно изолированы один от другого. На севере их разделяют густые леса на Склонах долин, а на юге - наиболее сухие степи. Таким образом, на окраинах ареала существуют две разобщенные части популяции, в которых возникновение общих волн жизни затруднено. Более того, рукава эти не сплошные. Чем дальше к северу от центральной области, тем больше заметно, что вид старается занять самые высокие гривки в долинах и выступающие места водоразделов. По мере же движения к югу вид "цепляется" только за самые влажные пятна в долинах рек и глубокие блюдца на водоразделах. Другими словами, здесь сама собой складывается та разобщенность микропопуляций, о которой в центральной области необходимо было позаботиться человеку. Значит, саранчовые нам не страшны? Да, но до тех пор, пока человек не нарушит естественных условий их обитания и сам себе не усложнит проблему защиты растений: ликвидировав в результате хозяйственной деятельности эту разобщенность популяции, он как бы облегчит создание новой оптимальной зоны ареала вида. В этом случае биотический потенциал может сыграть с нами злую шутку.
В северной части географических ареалов подобная картина складывается в результате вырубки лесов, слишком интенсивного осушения лесных земель, сильного вытаптывания пастбищ скотом. Все это увеличит приток солнечного тепла к земле, ликвидирует барьеры и откроет возможность для создания новой, сплошной и многочисленной популяции с характерными для нее массовыми размножениями. Но, пожалуй, наиболее существенные антропогенные (то есть вызванные деятельностью человека) изменения условий существования саранчовых можно ожидать в южных частях их ареалов в связи с развитием там орошения. Даже дождевание, при котором влага быстро испаряется с поверхности листьев, а влажность воздуха под ними поднимается лишь на короткий рок, способствует постоянному пребыванию на краях посевов и у дорог такого саранчового, как эпакромиус пульвирулентус.
В неорошаемой степи насекомые этого вида прячутся только в самых глубоких блюдцах с их солеными озерками. Естественно, что многие относительно влаголюбивые виды при орошении начинают появляться и в местах между своими прежними убежищами, в том числе и далеко отстоящими одно от другого. Особенно широко они распространяются, когда при строительстве оросительных каналов не предпринимают мер к предотвращению просачивания воды (инфильтрация), что вызывает образование заболоченных полос. По этим полосам вдоль каналов, подающих воду из долин рек в сторону водоразделов, могут легко продвинуться долинные популяции насекомых и сомкнуться с популяциями водоразделов. Вот и оказывается, что саранчовые не только "дети солнца", но и хорошие "гигрометры", способные реагировать на изменение влажности среды. Если геоботаники легко могут определить характер и направление происходящих в природе изменений (сукцессии) благодаря видам - индикаторам (показатели) из растительного царства, то ученые-энтомологи с не меньшим успехом предскажут такие изменения по видовому составу саранчовых, выявив более или менее влаголюбивые виды-индикаторы.
Итак, саранчовые - не только индикаторы ландшафтов, но и индикаторы сукцессии, то есть всех тех изменений, которые происходят в природе с участием или без участия человека.
Существенные перестройки внутреннего кружева ареала происходят в долинах рек, особенно тех, что текут с юга на север (типично для рек Сибири). Выше плотин подтопление надпойменных террас и переброска воды для орошения на водораздел создают предпосылки для продвижения влаголюбивых саранчовых (бесполосная кобылка) на водоразделы. Чтобы этого не случилось, рекомендуют перемещать сбросовые воды в долину при помощи дополнительных водосборных каналов, а ниже плотины сохранять увлажнение поймы при устройстве боковых водостоков. Тогда можно рассчитывать на регуляцию численности саранчовых в долинах рек.
Ниже плотин, напротив, происходит осушение пойм, так как плотина "гасит" высокие паводки, обычно заливающие поймы, и начинается остепнение пойм. Оно также может приводить к объединению ранее разобщенных популяций, но на этот раз теплолюбивых видов в северных частях ареалов.
Если в условиях, не измененных человеком, у северных и южных концов водораздельных и приречных рукавов популяции увеличивается разобщенность, то можно предположить, что обилие особей внутри этих популяций уменьшится. Однако происходит все наоборот. Популяции на самой периферии ареала, занимающие изолированные и малые по площади участки, как правило, оказываются даже более многочисленными, чем в оптимальной части ареала. Почему? Да потому, что они попадают в чрезвычайно благоприятные условия, сочетающие тепло и влагу. В то же время саранчовые находят здесь обилие пищи, а обычные для вида враги их не преследуют, отступая перед сыростью и густым травостоем. Заключенный в таком прекрасном, но маленьком убежище, вид не может распространяться в соседние ландшафты и создает на периферии ареала очень напряженные, небольшие по площади и жестко ограниченные очаги высокой численности вида. Их можно обнаружить в южных частях водораздельных рукавов популяции (возле озер) и на севере (на отдельных, хорошо прогреваемых песчаных холмах, гривах) как на водоразделах, так и в долинах.
Образованию таких очагов может способствовать хозяйственная деятельность человека. На юге очаги скорее всего будут складываться в местах скопления (застоя) сбросных оросительных вод на плоских водоразделах, на постоянно подтопляемых вследствие подпруды рек участках долин, а на севере - на участках, неумеренно осушенных и усиленно прогреваемых солнцем. Такие места появляются после вырубки леса и на пастбищах, вытоптанных скотом, вблизи скотоферм как на водоразделах, так и в особенности в долинах.
Внутреннее напряжение невольно создаваемых человеком очагов особенно велико. Непредвиденные последствия его хозяйственной деятельности могут быть очень серьезными. Именно в таких точках - у географических границ распространения вида, где среда, казалось бы, сдерживает размножение и расселение саранчовых, как раз и активизируется биотический потенциал. Отсюда вид может выплеснуться даже за пределы своего основного ареала.
В этом отношении особенно опасны влаголюбивые виды. Обычно они обитают в горах, где выпадает больше осадков, чем в предгорьях, и спускаются на -подгорные равнины лишь по прерывистым узким цепочкам, идущим по ущельям горных рек. При переходе с гор на равнину подобные реки дробятся на мелкие рукава, напоминающие дельты таких рек, как Нил, Волга. Выпадающие в них наносы образуют большие конусы, называемые в геоморфологии высшими, или промежуточными, дельтами. В промежуточных подгорных дельтах вода быстро просачивается в глубь наносов, поэтому ниже дельты река исчезает совсем или только на летние месяцы при ослаблении стока воды с гор.
Добравшись до промежуточных дельт, хорошо обводненных и прогреваемых лучами солнца, не заслоняемого облаками, как в горах, виды саранчовых образуют на них процветающие, но ограниченные краями дельты поселения. Это и есть подгорные очаги у самого предела распространения вида.
Здесь человек должен быть очень осторожен, поскольку сооружение даже небольших водохранилищ будет благоприятствовать саранчовым, а сельскохозяйственное освоение подгорий и прокладывание от гор на равнины каналов создаст предпосылки для бурного взрыва таких очагов при первой же волне жизни в них. Тогда насекомые начнут распространяться за пределы обычного географического ареала вида. Таким образом, мы опять наталкиваемся на необходимость включения биогеографических исследований в комплекс проектно-изыскательских работ, проводимых перед строительством оросительных систем. Кстати, данные, которыми располагают биологи, могут повлиять и на выбор конструкции оросительных систем. Известно, например, что многие виды, в том числе и итальянский прус, способны к массовому пассивному расселению с водой арыков, а его можно прервать простыми техническими средствами. Так что на будущей строительной площадке должны рука об руку работать и биолог, и техник.
Краевые (периферийные) очаги представляют большой интерес и с точки зрения эволюции. В этих очагах в погоне за исключительно благоприятным сочетанием тепла и влаги вид может оказаться в несколько необычном для него окружении. Скажем, растения будут иметь иные кормовые свойства, строение стеблей и листьев, чем у трав или кустарников, среди которых вид привык жить. Насекомым придется приспосабливаться к передвижению и питанию непривычным способом. Короче говоря, естественный отбор предъявит к виду новые требования.
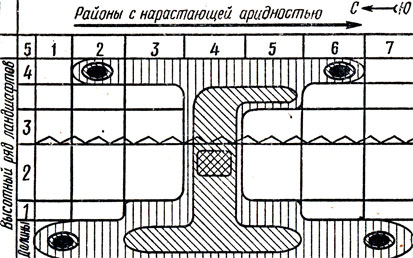
Схема ландшафтно-зонального ареала
Со временем в очаге должна образоваться особая популяция, а затем и внутривидовая группа, то есть произойти микроэволюция - превращение старого вида в новый. Если бы вид не имел таких очагов, он оказался бы куда менее пластичным, куда менее способным противостоять окружающей среде, смещающей границы ареала. Вид был бы эволюционно парализован, а возможно, и обречен. Краевые очаги -жизненно важные центры организации вида в пределах ареала. Вот почему эти очаги с точки зрения защиты растений нужно стремиться локализовать, а с точки зрения охраны природы - сохранять. Теперь мы видим, что иметь представление о внутреннем кружеве географического ареала вида очень важно и в теоретическом, и в практическом смысле, особенно для долгосрочного прогноза увеличения численности саранчовых.
Как же графически можно изобразить внутренний рисунок ареала? Пожалуй, в виде буквы "Н" с бусинками краевых очагов.
Прежде чем планировать системы ограничения численности какого-нибудь вида или меры его охраны в том или ином месте, нужно более или менее точно установить положение этого места на схеме. При помощи схемы можно определить, где и какие меры необходимы. В природе нам приходится иметь дело сразу со многими видами. Некоторые из них, пока не представляющие опасности для сельского хозяйства или не нуждающиеся в сохранении, могут по-своему реагировать на вмешательство человека.
Вот почему необходимы, накопление данных о локализации видов и составление схем их ареалов. Они послужат научной основой для долгосрочного не только пространственного, но и временного прогноза массовых размножений саранчовых.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'