Глава III. Экология лесных насекомых
Жизнь насекомых тесно связана с окружающей средой. В процессе эволюции под влиянием окружающей среды у насекомых вырабатывались приспособления к определенным условиям жизни и складывались разнообразные взаимосвязи с другими организмами и физическими условиями жизни. Изучением этих взаимосвязей на разных уровнях организации живых систем занимается экология насекомых.
В настоящее время экология получила бурное развитие и часто трактуется очень широко и противоречиво. Обсуждение этого вопроса не входит в задачи курса лесной энтомологии.
Общие сведения
Жизнь насекомых проходит под воздействием множества экологических факторов. Экологическим факторов называют любой элемент среды, способный оказать непосредственное влияние на живые организмы, а также на характер их отношений друг с другом. Число всевозможных экологических факторов потенциально является неограниченным. Однако по степени воздействия на организмы эти факторы не равнозначны. Поэтому при их анализе всегда выделяются наиболее существенные. По каждому фактору имеется диапазон выносливости, за пределами которого организм не способен существовать. Следовательно, любой фактор может выступать как лимитирующий, если он отсутствует, находится ниже критического уровня или превосходит максимально выносимый уровень. Так, например, распространение подкорного соснового клопа в посадках сосны лимитирует их возраст, с которым связано появление чешуи коры, защищающих клопа при питании и метаморфозе.
По аналогии с химией, в экологии существует понятие экологической валентности. Экологическая валентность (или пластичность) вида - его способность заселять разнообразные местообитания. Виды с низкой экологической валентностью способны выносить лишь ограниченные изменения экологических факторов и называются стенотопными (греч. стено - узкий). Виды, способные заселять очень различные местообитания и переносить широкую амплитуду колебаний экологических факторов, называются эвритопными (греч. эври - широкий). Такие виды насекомых легче расселяются по территории, выживают и размножаются в различных условиях, чаще всего имеют более широкую область распространения.
Область распространения вида называется ареалом. По ареалу вид обычно распространен неравномерно и всегда представлен более или менее прерывистыми поселениями. Между этими отдельными поселениями лежат пространства, непригодные для данного вида или занятые другими видами - более сильными конкурентами. В зависимости от особенностей организмов и характера территории размеры таких поселений могут быть самыми разнообразными как по численности особей, так и по величине занимаемой территории. Каждое такое поселение представляет собой популяцию. Большинство видов широко распространенных насекомых состоит из множества популяций. Следовательно, популяция (лат. populus - народ) - это группа особей одного вида, занимающая определенное пространство. Она является основной естественной единицей существования, приспособления и воспроизведения вида. Популяция обладает многими признаками, которые характеризуют группу как целое, а не отдельные особи в группе. Основные из них - плотность, рождаемость, смертность, возрастной состав, характер распределения в пределах территории и тип роста.
Популяции отдельных видов взаимодействуют между собой на определенной, более или менее однородной территории. Такая территория называется биотопом.
Живое население биотопа получило название биоценоза (греч. биос - жизнь, ценос - вместе). Следовательно, каждый вид, представленный местной популяцией, является членом определенного биоценоза.
Биоценоз представляет собой исторически сложившийся комплекс организмов и является частью общего природного комплекса - биогеоценоза или экосистемы, включающего кроме организмов почву, водный режим, геологическое строение и климатические условия, свойственные данной территории.
Действие экологических факторов на насекомых
Жизнь популяции любого вида насекомого проходит под контролем экосистемы, в которой она существует и с которой соединена множеством связей. В настоящее время еще невозможно сразу учесть весь комплекс факторов, влияющих на популяции и отдельные особи насекомых. Поэтому обычно последовательно рассматривают влияние отдельных главнейших факторов.
Существует ряд классификаций экологических факторов. Наиболее простое и распространенное деление их на биотические и абиотические факторы. Однако такое деление в значительной мере произвольно и недостаточно. Поэтому лучше все экологические факторы делить на климатические, эдафические (почвенные), пищевые и биотические (внутривидовые и межвидовые взаимодействия организмов) и антропогенные.
Климатические факторы
Основные климатические факторы - температура, влажность воздуха, осадки, свет, ветер, атмосферное давление. Кроме того, существенное влияние на насекомых оказывают климат в целом и непрерывно меняющиеся погодные условия.
Климат определяет распространение насекомых, границы их ареалов, а погодные условия влияют на сроки их развития и колебания численности в популяциях.
Температура
Насекомые - пойкилотермные животные, они не имеют постоянной температуры тела. Если насекомые находятся в состоянии покоя, то температура их собственного тела определяется температурой окружающей среды. В полете или под действием солнечной радиации температура тела насекомых значительно повышается. При этом дыхание становится более интенсивным и поглощение кислорода увеличивается. По наблюдениям И. Д. Стрельникова (1935), у крупных и сильных бабочек (бражников) температура тела под влиянием мышечной работы при летательных движениях крыльев в течение 30 мин поднимается на 18 - 20° С по сравнению с состоянием покоя. У азиатской саранчи после 2,5 мин полета температура тела увеличивается на 10° С.
При облучении солнечной радиацией температура тела насекомых быстро поднимается, вызывая их активность. Суточный ритм многих насекомых тесно связан с температурой. Перед восходом солнца хищные мухи ктыри безжизненно сидят на стеблях вейника и начинают пробуждаться по мере подъема температуры. При этом время появления отдельных видов тесно связано с их происхождением: раньше появляются виды северного происхождения и позднее, в самые жаркие часы дня, - тропического.
Регуляция теплообмена достигается насекомыми благодаря охлаждающему действию испарения с поверхности тела, через стигмы при дыхании и особенно за счет активных перемещений. При этом на интенсивность теплообмена оказывают влияние величина, форма и окраска тела. Например, у жуков златок с блестящей металлической окраской температура на солнечном свету ниже, чем у насекомых с другой окраской тела. Весьма обычно перемещение насекомых с освещенных мест в затененные, использование укрытий, зарывание в песок и лесную подстилку.
У каждого вида насекомого существует предпочитаемая им температура. Она может быть различной для разных фаз развития и меняется в различные периоды жизни в зависимости от внешних условий. Такая температура, привлекающая большинство особей в популяциях данного вида, получила название термического преферендума.
Знание термического преферендума вредных насекомых в разные периоды жизни облегчает надзор за ними, организацию мер борьбы - использование отравленных приманок, ловушек, назначение сроков химических обработок отдельных объектов и т. д.
Активная деятельность насекомых ограничена определенными температурными границами - нижним и верхним порогами развития. Нижний температурный порог равен примерно +5 - 8°С, изменяясь у отдельных видов от -1 до +10° С (у зимней пяденицы - - 1 С, а у посевного шелкуна +10° С). Он меняется на разных фазах развития насекомых и зависит также от их происхождения. У насекомых южного происхождения он выше, северного - ниже. При снижении температуры насекомого за пределы нижнего порога организм впадает в состояние холодового оцепенения, или депрессии.
Верхний термический порог также зависит от вида и фазы развития насекомого, но не превышает 40° С, чаще всего находясь в интервале 30-35° С. За этими пределами насекомые впадают в тепловое оцепенение.
Температурой определяется как самая возможность жизни насекомого, так и интенсивность ее проявления, благодаря изменению уровня обмена веществ, скорости развития, роста, интенсивности питания и размножения.
При изучении влияния температуры на организм насекомых выявлены некоторые общие закономерности, характеризующие его состояние при разных температурах. Схематически выделяется шесть градаций, или температурных зон, характеризуемых определенным поведением и физиологическими процессами, протекающими у насекомых в их пределах. Они нанесены на температурную шкалу, изображенную на рис. 12 в виде линейки АБ с интервалом от + 100 до -273° С. В пределах верхней смертельной зоны после короткого периода возбуждения наступает смерть вследствие инактивации ферментов и коагуляции белков протоплазмы.
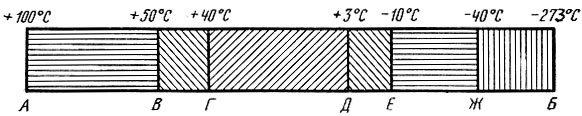
Рис. 12. Схематическое изображение отношения насекомых к температурным воздействиям: Зоны: АВ - верхняя летальная, ВГ - верхняя сублетальная, ГД - витальная, ДЕ - нижняя сублетальная, ЕЖ - нижняя летальная, ЖБ - витрификации и анабиоза
Верхняя сублетальная зона характеризуется тем, что в ее пределах при известном сочетании интенсивности температуры и продолжительности ее воздействия возможно выживание насекомых.
Зона активной жизнедеятельности (витальная) содержит оптимум развития, ниже и выше которого имеется область пониженной жизнедеятельности организма.
Оптимум характеризуется умеренной скоростью развития при минимальной затрате энергии и при наименьшем вымирании, а также наибольшей продолжительности существования взрослой фазы насекомого и высокой плодовитости. Следовательно, в области оптимума содержатся наилучшие условия для процветания популяции вида (рис. 13).
Зона температурного оптимума специфична для каждого вида и фазы развития. Она располагается в области 15 - 30° С. Наиболее высоких темпов развитие достигает при 28 - 33° С, а температурный интервал между оптимумом и этой температурой составляет 10 - 12° С. Нижняя сублетальная зона - это зона холодового оцепенения и переживания, а нижняя летальная зона характеризуется замерзанием и кристаллизацией жидкости тела и повреждением протоплазмы. В определенных пределах эти явления вызывают смерть.
В зоне витрификации или анабиоза возможно затвердевание живой протоплазмы. Здесь достигается полная приостановка процессов обмена с потенциальным сохранением жизни и возможности его последующего восстановления.
Каждому насекомому для его развития необходимо определенное количество тепловой энергии, называемой суммой эффективных температур. Она складывается из суммы среднесуточных температур, наблюдаемых в данной местности, за вычетом нижнего порога развития. Например, если наблюдаемая температура (Т) равна +25°С, а нижний порог развития (t) равен +10° С, то эффективная температура (Т - t) будет равна 25°С - 10°С = 15°С. Сумма эффективных температур равна эффективной температуре, умноженной на число дней развития (n), выражается формулой
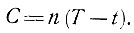
Для развития насекомого в данной местности требуется определенный минимум дней с температурой выше пороговой, так как именно порог определяет возможность развития.
Для каждого вида экспериментальным путем можно определить порог и длительность развития и на основе их вычислить сумму эффективных температур. Так, для полного цикла развития кольчатого коконопряда она равна 1470° С, для монашенки - 1239° С и т. д. По сумме эффективных температур и порогу развития можно узнать длительность развития. Подобного рода расчеты часто бывают полезны при краткосрочных прогнозах появления насекомых и для определения зависимости сроков развития от температуры. Графически эта зависимость выражается кривой, наиболее близкой к гиперболе (рис. 14). По оси ординат отмечаются данные температуры, по оси абсцисс - число дней развития. В точке порога развития гипербола почти параллельна горизонтальной оси, т. е. развитие при данной температуре стремится к бесконечности.
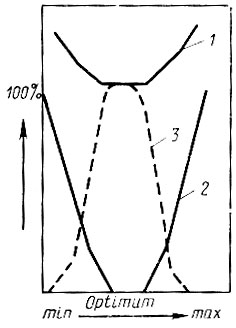
Рис. 13. Критерии оптимума развития (по Кожанчикову, 1961): 1 - расход энергии при развитии, 2 - смертность предимагинальных фаз, 3 - плодовитость самок
Опустив перпендикуляр из любой точки гиперболы на оси системы координат, можно установить продолжительность развития насекомого при заданной температуре и, наоборот, определить температуры при известной длительности развития.
Строить гиперболы и определять суммы эффективных температур можно для всего цикла развития насекомого и для отдельных его фаз.
Несмотря на широкое применение в практике, метод сумм эффективных температур не дает точных данных о длительности развития насекомых, которое происходит при переменных температурах, а не при постоянной. Колебания температуры в течение суток приводят к ускорению или замедлению развития по сравнению с таковыми при постоянной температуре. Большое влияние на длительность развития оказывают также другие сопряженные с температурой факторы, например влажность воздуха. Кроме того, насекомые приурочены в своем развитии к определенным местообитаниям, температура в которых сильно отличается от наблюдаемой в метеорологической будке.
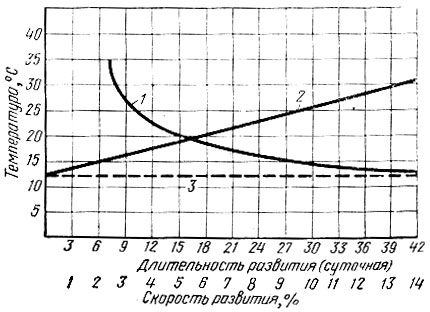
Рис. 14. Схема зависимости сроков развития насекомых от температуры (по Кожанчикову, (1961): 1 - длительность развития в сутках, 2 - скорость развития в процентах, 3 - порог развития
Температура влияет на все жизненные процессы насекомых, ускоряя одни и замедляя другие. Она влияет также на морфологические особенности (на величину тела, окраску, развитие крыльев, формирование крыльев и др.), поведение, географическое распространение и численность насекомых.
Повышение температуры до известных пределов стимулирует питание насекомых, линьку и активность спаривания, влияет на восприимчивость к пестицидам и болезням.
Выбор местообитаний насекомыми часто зависит от температуры. Теплолюбивые жуки златки, например, заселяют насаждения по хорошо прогреваемым южным опушкам леса и экспозициям. Перемещение насекомых из одного местообитания в другое также происходит под влиянием температур. Так, температура на вырубках быстро поднимается в дневные часы, и жуки большого соснового слоника мигрируют на неосвещенную часть вырубки, где температура на 15° С ниже. Непарный шелкопряд в южной части ареала откладывает яйца по всему стволу дерева, а в северной - только в нижней его части. Здесь яйца защищены от сильных морозов снеговым покровом, а выше его линии обычно вымерзают.
Распространение на север многих видов насекомых ограничено зимними низкими температурами, при которых происходит гибель популяции. В годы с сильными морозами наблюдается очень большая смертность личинок майского хруща в почве, яиц листоверток, зимней пяденицы и др.
Влажность
Влияние влажности воздуха на насекомых осуществляется несколькими путями. Насекомые испаряют много воды через покровы тела и трахейную систему и поглощают воду непосредственно в пище и при дыхании с водяными парами воздуха.
Вода в теле насекомого находится в свободном состоянии или адсорбирована различными веществами тела, при окислении которых освобождается и поступает в общий обмен веществ. При недостаточном поступлении извне организм использует воду, образующуюся при окислении жира. Соотношение воды и жира постоянно меняется в теле насекомых и может служить одним из критериев их холодостойкости. Чем больше жира и меньше воды в теле насекомого, тем лучше оно переносит низкие температуры. Так, гусеницы златогузки второго возраста, которые поздней осенью содержали 68 - 72% воды и 18,43% жира к сухой массе, выдерживали охлаждение при температуре - 15° С до 158 дней. Гусеницы той же бабочки, развивавшиеся весной до четвертого возраста, имели 84,08% воды и 4 - 6% жира. Они погибали при температуре - 15°С через 4 ч (Ушатинская, 1957).
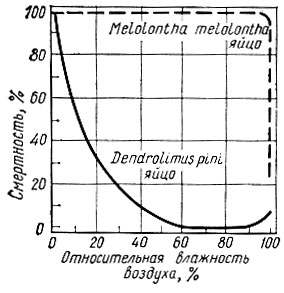
Рис. 15. Выносливость яиц насекомых с различной биологической пластичностью к влажности воздуха
Накопление жира у насекомых обычно происходит перед зимовкой. Однако есть и такие насекомые, которые отличаются высокой холодостойкостью, но зимуют с малым содержанием жира и большим количеством воды в теле. Так, зимующие гусеницы соснового шелкопряда, выдерживающие охлаждение до - 20° С, перед наступлением зимы имеют всего 1,2% жира и 77,9% воды. Такое явление свойственно насекомым, зимующим в фазах, на которых за весенним пробуждением сразу следует питание.
Недостаток влаги в атмосфере компенсируется также усиленным потреблением пищи, содержащей влагу. Через пищу влажность влияет и на скорость развития. В сухой древесине развитие домового усача и других ксилофагов замедляется и может растянуться на три-четыре года.
Потеря воды регулируется различными путями. Восковой слой кутикулы задерживает испарение через кожу; испарение через трахеи регулируется замыкательным аппаратом дыхалец, а мальпигиевы сосуды адсорбируют воду из содержимого кишки и задерживают ее в организме.
Большинство лесных насекомых может развиваться в довольно широких диапазонах влажности воздуха. Избыток влажности сказывается преимущественно на длительности развития, но не препятствует продолжению жизненных функций. Недостаток влажности переносится хуже и скорее вызывает гибель насекомого (рис. 15).
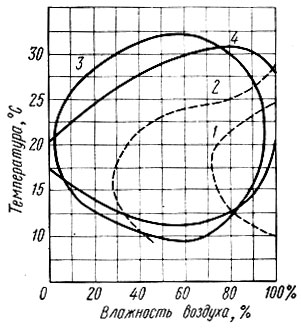
Рис. 16. Термогигрограмма оптимум для яиц волнянок (по Кожанчикову, 1950): 1 - для яиц монашенки весной, 2 - для яиц монашенки в осенне-летний период, 3 - для яиц непарного шелкопряда весной, 4 - для яиц непарного шелкопряда в осенне-летний период
По отношению к влажности воздуха среди насекомых различают влаголюбивые (гигрофильные) и сухолюбивые (ксерофильные)виды. Промежуточная группа носит название мезофилов. Лесные насекомые приспособлены к повышенной влажности воздуха и в подавляющем большинстве мезо- и гигрофилы. Тольке часть видов, обитающая на опушках и в наиболее сухих местах, может быть по сравнению с другими названа ксерофилами. Так, сосновая совка более ксерофильна, чем сосновая пяденица или монашенка, а вершинный короед сухолюбивее сосновых лубоедов и т. п.
Действие влажности всегда сопряжено с температурой. Одинаковая влажность может давать различный эффект при разных температурах. Для изучения эффектов различных комбинаций этих факторов проводятся опыты в термостатах с устойчивой влажностью. В результате таких опытов строятся термогигрограммы, позволяющие судить о требовательности данного вида насекомого на разных фазах его развития к гигротермическим условиям ( рис. 16) и выносливости по отношению к ним. Так, термогигрограмма, составленная для яиц волнянок, показывает, что зона оптимального развития связана с довольно узкими пределами влажности воздуха у одних видов (монашенки) и, наоборот, весьма широкими у других (непарный шелкопряд). Термогигрограмма для соснового пилильщика может быть еще названа диаграммой смертности. Она показывает размеры смертности личинок при разном сочетании влажности и температуры (рис. 17).
Влажность оказывает большое влияние на распространение насекомых в лесу. Очевидно, что требовательные к влажности виды будут держаться в более увлажненных местах, а ксерофилы будут занимать наиболее сухие местообитания. Так, короед дендроктон часто приурочен к сфагновым соснякам и развивается в прямом контакте с влагой. Осушение болота ведет к исчезновению этого короеда. В поймах рек ветла иногда стоит наголо объеденной ивовой паутинной молью. Паутина охватывает все кроны, местами образуя сплошной полог, висящий над зеркалом воды. Вредителям мертвой древесины также необходима для развития повышенная влажность. Личинки долгоносика-трухляка не боятся полного насыщения обрубков древесины влагой. В противоположность этим видам златки развиваются при очень незначительной влажности воздуха и субстрата, несмотря на окружающую высокую температуру.
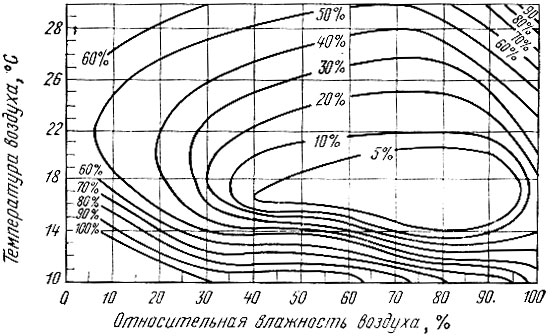
Рис. 17. Термогигрограмма для личинок обыкновенного соснового пилильщика (по Гёссвальду, 1935)
Осадки
Осадки - один из самых мощных факторов среды. Они влияют на насекомых прямо и косвенно, через изменение влажности, растительность и температуру почвы зимой.
Прямое влияние осадков не избирательно. При сильных наводнениях, ливнях и крупном граде погибает огромное количество насекомых, только очень немногие случайно избегают гибели. При очень сильной засухе также наблюдается депрессия организма и часто очень большая смертность особей данного вида.
Косвенное влияние осадков очень ярко проявляется через растительность. В годы сильных засух происходит нарушение водного обмена деревьев и их защитные функции (особенно осмотическое и смоляное давление) ослабевают. В результате происходит заселение деревьев обитателями стволов, и они получают возможность быстро размножаться. При засухе изменяется биохимический состав листьев в сторону, благоприятную для питания и успешного развития листогрызущих насекомых. Падение осмотического давления в листьях способствует их заселению тлями, смоляного давления в хвое - выживанию яиц одиночно кладущих пилильщиков и побеговьюнов в почках сосны.
Большое влияние на выживаемость насекомых оказывают зимние осадки в виде снега. Снежный покров вследствие малой теплопроводности обладает высокими теплоизоляционными свойствами. Усиленно отдавая тепло с поверхности, он предохраняет от охлаждения почву. Поэтому насекомые, зимующие в почве, лесной подстилке под корой поваленных деревьев и у основания стволов, надежно защищены снежным покровом от вымерзания. В этом случае для выживаемости насекомых под снежным покровом большое значение имеют сроки выпадания и таяния снежного покрова, его мощность и температура воздуха в период, когда почва оголена. Хорошо известны факты вымерзания в почве личинок майского хруща и куколок подгрызающих совок, а под корой бревен - короедов.
Годы с количеством осадков ниже нормы часто совпадают с температурой выше нормы, вследствие чего складывается засушливая погода. Очень многие исследователи связывают наступление засушливой погоды с последующим размножением ряда вредных насекомых По утверждению других, ряд видов повышается в численности после влажных лет. В связи с этим делаются попытки увязать изменение численности отдельных видов насекомых с количественными показателями засушливости. Так, по данным Б. В. Флерова (1958) нарастание численности сибирского шелкопряда начинается после двух-трех лет с большим дефицитом влажности. Обычно мерилом засушливости служит гидротермический коэффициент Селянинова. Формула этого коэффициента выведена эмпирически:
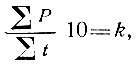
где k - гидротермический коэффициент;
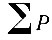
- сумма осадков за тепловое время года, когда температура выше нуля;
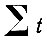
- сумма положительных температур для данного места.
Применительно к задачам энтомологических работ И. А. Рубцовым (1938) этот коэффициент был предложен в следующем виде:
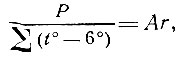
где P - годовая сумма осадков для данного места.
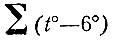
- сумма средних месячных температур, лежащих выше 6 С; Аr - гидротермический коэффициент.
Анализируя изменения этого коэффициента за период с 1860 по 1956 г. в Башкирии, М. Г. Ханисламов (1958) пришел к выводу, что начало вспышки массового размножения непарного шелкопряда является следствием сочетания низких гидротермических коэффициентов мая - июня с сильными зимними морозами.
Для оценки годичного цикла температуры и увлажнения можно пользоваться специальными графиками - климограммами. Они составляются на основании данных о средних месячных температурах и месячных суммах осадков. По оси ординат откладываются величины температуры, по оси абсцисс - количество осадков. Точки пересечения, найденные для каждого месяца по климатическим данным, соединяются в годичной последовательности. Климограммы могут составляться для отдельных лет и по многолетним средним данным. С помощью климограмм можно составить представление о типичном для данного вида насекомого климатическом режиме. На рис. 18 приводятся климограммы, составленные за ряд лет для определения условий, при которых повторяются вспышки массового размножения дубовой хохлатки. На рис. 19 даются сравнительные климограммы, составленные по многолетним данным для Москвы и Урды (Западный Казахстан).
Свет. Свет оказывает прямое и косвенное влияние на насекомых. При прямом влиянии света большое значение имеют его дозировки. По этому признаку можно различить насекомых с разным диапазоном восприятия световых условий. Ночным и почвенным насекомым нужны узко ограниченные световые условия, а виды с круглосуточной активностью имеют широкий диапазон реагирования на освещенность.
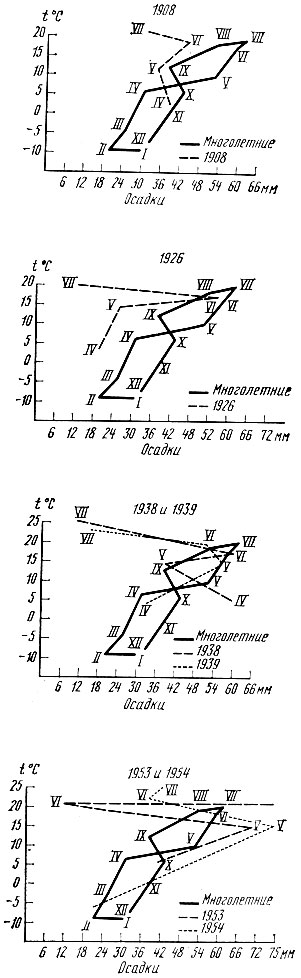
Рис. 18. Климограммы за 1908, 1926, 1938-1939 и 1953-1954 гг. по Воронежской обл. (по Егорову, 1960)
Освещенность измеряется в люксах и в значительной мере определяет общую активность насекомых и ряд важнейших жизненных процессов: оплодотворение, яйцекладку, выход имаго из куколки и и др. Так, вечерний полет майского хруща всегда начинается при освещенности, равной 1250 - 1400 лк. В зависимости от освещенности изменяется поведение насекомых. При этом одни виды светолюбивы, другие тенелюбивы. Так, например, жуки златки летают, спариваются и откладывают яйца только при ярком солнечном свете. Большое значение в жизни насекомых имеет тепловое действие солнечной радиации. Обогревание солнечным светом повышает температуру тела насекомого на 10° С и больше. Под влиянием солнечной радиации происходит нагревание частей дерева и почвы, которые выбирают для своего поселения или которых избегают насекомые.
Насекомые тонко реагируют на изменение длины дня. Длина дня - один из регуляторов сезонных циклов развития, особенно сказывается на появлении диапаузы. У насекомых, реагирующих на изменение длины дня, наблюдается три основных типа фотопериодической реакции, в известной мере аналогичных типам, установленным для растений,- длиннодневный, короткодневный и промежуточный (рис. 20). В условиях умеренного климата преобладает длиннодневный тип, к которому относится большинство листогрызущих насекомых. Противоположный тип фотопериодической реакции имеют виды короткодневные. У таких видов непрерывное развитие происходит лишь в условиях короткого дня. Под влиянием длинного дня происходит торможение роста и наступает диапауза, третий тип реакции - промежуточный - характерен для чешуекрылых, диапаузирующих на фазе гусениц.
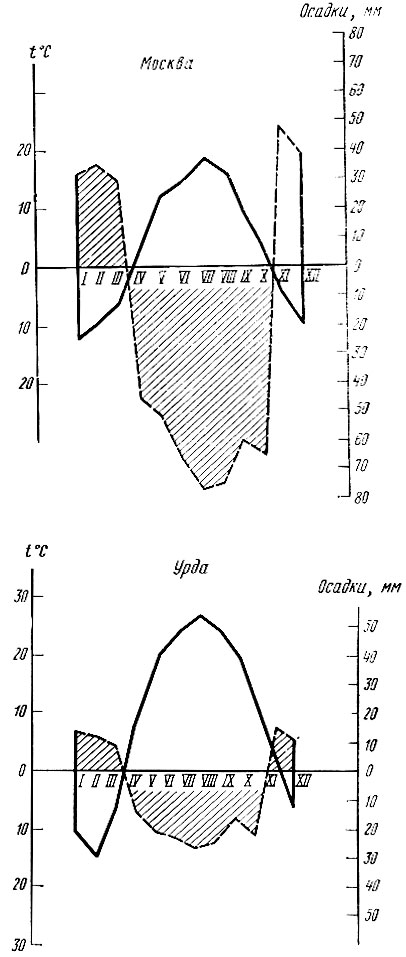
Рис. 19. Климограмма для Москвы и Урды (по многолет. данным)
Ветер
Роль ветра в распространении насекомых может быть довольно значительной. Так, например, формирование и перемещение очагов непарного шелкопряда в Крыму связано с меняющимся направлением ветра и встречными естественными преградами. Известно много случаев заноса насекомых ветром и воздушными течениями, например тлей с Кольского полуострова на снежные массивы Шпицбергена.
Ветер определяет характер погоды и тем самым оказывает косвенное влияние на поведение насекомых, их миграции, интенсивность питания и размножение в насаждениях.
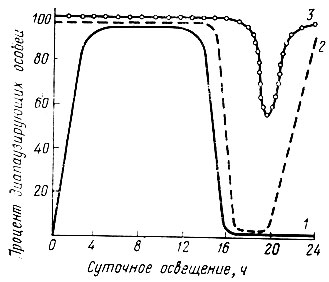
Рис.20. Типы фотопериодической реакции насекомых (по Гейспиц, 1953): 1 - капустница - длиннодневный тип, 2 - желтогузка - промежуточный тип, 3 - ивовая волнянка - промежуточный тип
В лесах степной и лесостепной зон ветер является постоянно действующим фактором. Он влияет на поведение насекомых не только тем, что разносит их, но и тем, что вызывает большое испарение влаги телом насекомых. Кроме того, насекомые вынуждены концентрироваться на участках, защищенных от ветра. Ветер раскачивает стволы деревьев, вызывая бурелом и ветровал в лесах. В этих местах потом образуются очаги короедов и других вредителей стволов деревьев.
Почвенные факторы
Почва служит средой для обитания многих насекомых, которые вместе с другими многочисленными представителями беспозвоночных (дождевые черви, многоножки, клещи и др.) составляют общий почвенный биоценотический комплекс организмов. Наряду с вредителями корневых систем растений здесь встречаются хищники, истребляющие вредителей, и сапрофаги, перерабатывающие растительный опад и рыхлящие почву. Многие насекомые используют почву как временное убежище (зимовка, укрытие от высоких температур, окукливание).
Исторически почва для многих насекомых явилась той промежуточной средой, в которой осуществился их переход от водного к наземному образу жизни (Гиляров, 1949).
Обитающие в почве личинки насекомых-фитофагов имеют целый ряд приспособлений для передвижения. Эти приспособления зависят от типа почв, в которых обитают насекомые, и служат видовыми признаками. Так, у личинок пластинчатоусых - С-образной формы, для закрепления тела от скольжения имеются шипики и крючковатые щетинки, расположенные на последнем стерните. При прокладке хода личинка приподнимает кверху и откидывает назад переднюю часть тела, придавая ей форму, близкую к латинской букве S, вонзает мощные и острые челюсти в почву и, сокращая продольные мускулы брюшной стороны тела, снова принимает С-образную форму. Благодаря последнему движению происходит откалывание кусочка почвы воткнутыми в нее челюстями, как киркой, и отгребание его нижней стороной головы к заднему концу туловища. В более уплотненной почве, откалывая ее кусочки, личинки прибегают к грызущему движению челюстей. В отгребании кусочков почвы, кроме головы участвуют также ноги.
Личинки щелкунов (проволочники) передвигаются при содействии уплощенной и сильно хитинизированной головы, снабженной верхними заостренными челюстями. Закрепляя задний конец тела в прокладываемом ходе при помощи расположенных на нем подталкивателя и хитинизированных отростков, личинка откалывает челюстями, как ломом, кусочки почвы и отгребает их назад ногами при помощи лопатообразной тергитной площадки, снабженной по бокам зубцами.
Насекомые живут в самых разнообразных почвах, однако большинство широко распространенных видов предпочитает легкие песчаные и супесчаные почвы, избегая тяжелые глинистые и переувлажненные почвы. Здесь, как и среди наземных фитофагов, имеются насекомые, обладающие различной степенью пластичности. Так, личинки европейского мраморного хруща и хищных мух ктырей приурочены только к песчаным сухим почвам. Личинки же июньского хруща встречаются в самых различных почвах: песчаных, супесчаных, глинистых.
Границы вертикального распространения насекомых в почве связаны с ее насыщением корневыми системами и гумусом. В почвах, богатых гумусом, фауна обильнее и разнообразнее. Наличие гумуса в свою очередь связано с развитием лесной подстилки, которая оказывает большое влияние на выживаемость насекомых в почве. Лишенные лесной подстилки почвы сильно промерзают зимой, а летом перегреваются и высыхают. В лесной подстилке зимуют и окукливаются многие вредители леса.
Большинство насекомых-фитофагов, живущих в почве, многоядны. Характер питания чаще всего оказывает большое влияние на их численность и распространение.
Выживаемость почвообитающих насекомых зависит также от почвенных условий (температура, влажность, кислотность почвы и др.). Многие виды очень тонко реагируют на изменение кислотности почвы, ее температуру и влажность. Подробнее экология почвообитающих насекомых, вредящих корням древесных растений, описана в гл. VI.
Пищевые факторы
Существование каждого вида насекомого прежде всего определяется обменом веществ, в процессе которого организм непрерывно расходует энергию и вынужден поэтому восполнить ее. Восполнение энергии связано с процессом питания. Пища оказывает влияние на все жизненные процессы насекомых и служит важнейшим экологическим фактором.
Характер питания насекомых
По характеру питания насекомые делятся на ряд экологических групп. Представители всех этих групп встречаются и в лесах, играя определенную роль в круговороте веществ. Основными из них являются: фитофаги (питаются только растительной пищей), зоофаги, или плотоядные (питаются только животной пищей), детритофаги (питаются мертвым органическим веществом), сапрофаги (питаются разлагающимися растительными веществами), некрофаги (питаются трупами животных) и копрофаги (питаются навозом). К зоофагам относятся кровососы и энтомофаги (рис. 21).
Фитофаги, живущие за счет древесных растений, часто еще называются дендрофильными насекомыми. С лесом связано очень много видов дендрофильных насекомых, населяющих все ярусы растительности, лесную подстилку и почву. Видовое разнообразие их определяется лесной средой и в первую очередь составом насаждений. Чем больше древесных пород входит в состав насаждения, тем богаче фауна вредных насекомых. С каждой древесной породой связан определенный комплекс видов лесных насекомых. В пределах древесной породы обычно различают вредителей плодов и семян, почек, листьев, ветвей, стволов и корней. При этом бывают такие случаи, когда при недостатке корма или в определенных географических условиях обитатели листьев начинают повреждать плоды или почки, вредители ветвей переходят на стволы и даже корни и т. д.
Выбор древесной породы и возможность питания определенным числом пород обусловлены пищевой специализацией насекомых, сложившейся в процессе их эволюции.
Специализация питания
Специализация питания насекомых развивается на основе химических и биологических особенностей растений, служащих им пищей. Количественно пищевая специализация может быть охарактеризована числом используемых для питания видов древесных пород.
По пищевой специализации, характеризующей степень требовательности к пище различают одноядных (монофаги), ограниченно-ядных (олигофаги) и многоядных (полифаги) насекомых. Среди вредителей древесных пород преобладают олигофаги.
Монофаги в чистом виде встречаются редко. В практике чаще всего относят к монофагам виды насекомых, повреждающих несколько очень близких древесных пород, относящихся к одному роду. Примером может служить березовый заболонник, повреждающий только березу. Однако он не делает различия между березой пушистой и бородавчатой. Алтайский усач повреждает только лиственницу, но также не отличает сибирскую от лиственницы Сукачева. Многие короеды, селящиеся на ели обыкновенной, повреждают также и ель сибирскую. Берестовый листоед отдает явное предпочтение бересту, но может питаться и на других ильмовых.
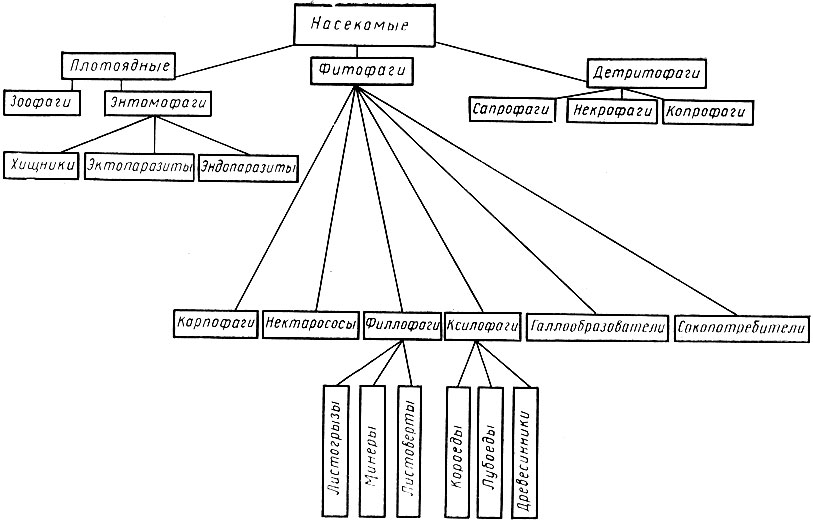
Рис. 21. Классификация насекомых по характеру питания
Полифагия, многоядность насекомых, представляет обратное явление их пищевой специализации. Она выражает возможность насекомого использовать для роста в развитии разнообразные и часто биохимически далекие вещества. Полифагия зависит не только от переваривающей способности организма, но главным образом от возможностей органического синтеза у данного вида. Однако многоядность не есть всеядность. Даже многоядные виды отдают явное предпочтение ограниченному кругу кормовых растений, на которых могут нормально завершить развитие и давать плодовитое потомство. Прекрасным примером служит непарный шелкопряд, признанный в учебной литературе широким полифагом в связи с тем, что зарегистрирован как вредитель более 300 видов растений, включая хвойные породы, культурные злаки, клюкву, полынь и огородные культуры.
В настоящее время установлено, что несмотря на многоядность гусениц непарного шелкопряда, они нормально развиваются и дают плодовитых бабочек на сравнительно ограниченном круге растений. Оказалось, что при поедании древесных пород у гусениц непарного шелкопряда нарушается обмен веществ и, как следствие этого, наблюдается низкая выживаемость популяции. В зоне степного лесоразведения к таким породам относятся липа и клены остролистный и ясенелистный. И. В. Кожачников (1941, 1951) указывает, что непарный шелкопряд специализирован на питании букоцветными растениями, в соке которых имеются прежде всего специфичные белки, а также таннины и некоторые другие вещества. С букоцветными сходны по химизму ивоцветные и розоцветные, но среди них есть уже много видов малопитательных, например кустарниковые ивы и травянистые розоцветные.
Пищевые связи большинства лесных насекомых сложились на основе биохимического родства кормовых растений, имеющих близкий состав белка. Белковое питание определяет процессы роста и размножения организма. Поэтому набор белков и их химическая специфика - решающее условие при выборе растения насекомыми.
Специфические растительные вещества (глюкозиды, алкалоиды, таннины, органические кислоты, фенолы и др.) чаще всего являются сигнализаторами, уточняющими биологическую связь. Они определяют поведение насекомого, но не играют решающей роли в его росте и развитии. Однако присутствие в растениях специфических веществ может порождать частные формы приспособления обмена веществ к их использованию. Так, например, олигофагия тополевого листоеда (Melasoma populi), вероятно, связана с использованием салициловой кислоты, ограничившей крут его кормовых растений.
Древесные породы могут отвергаться рядом насекомых и потому, что в их тканях содержатся пищевые вещества, совершенно непригодные для питания данного вида. Так, например, гусеницами непарного шелкопряда в опытах отвергались растения, в которых содержались в большом количестве молочная кислота, некоторые алкалоиды и эфирные масла. Избыточное содержание эфирных масел в хвое отдельных пихт защищает их от повреждения гусеницами монашенки.
Приспособление обмена веществ насекомых к определенному химизму служит основой для специализации питания на определенных тканях или частях растения. Каждый вид насекомого приспособлен к питанию на определенной части дерева: цветках, плодах и семенах, листьях, почках и побегах, на ветвях, стволе или корнях. Каждая часть растения построена из определенных тканей, содержащих различные пищевые вещества. Например, в меристеме много белка, в плодах и семенах также преобладают белки и запасные вещества, в листьях много углеводов, а в центральных слоях древесины - клетчатка и лигнин. Насекомые, живущие за счет листьев, не повреждают древесины, а живущие в древесине не переходят на плоды и семена. Однако при недостатке пищи или резком изменении физических условий можно наблюдать частичные переходы насекомых на питание другими тканями. Личинки усачей, живущие за счет луба под корой, часто углубляются в древесину, гусеницы сосновой совки кроме хвои поедают почки и обгладывают побеги, короеды меняют места поселения на дереве и т. п.
Не менее определенно приспособление питания лесных насекомых к фазам вегетации древесных пород. Химизм растений меняется в связи с их развитием в течение цикла вегетации. Эти изменения особенно отчетливо обнаруживаются в листьях. Весной листья древесных пород богаты водой и белками, но бедны углеводами, в частности растворимыми сахарами и клетчаткой. Листья, прекратившие рост, становятся более бедными водой и белками, но обогащаются углеводами, а позднее балластным материалом. В соответствии с этим имеются насекомые, приспособленные к питанию только весенним листом (непарный шелкопряд, зимняя пяденица, дубовая листовертка и др.) и питающиеся летним листом (лунка серебристая, краснохвост и др.).
Смена кормовой породы
В природе часто можно наблюдать закономерную и вынужденную смену лесными насекомыми их кормовых пород. Смена кормовых пород может быть временной, когда она обусловлена недостатком корма, регулярной, когда насекомое последовательно и закономерно питается двумя или несколькими породами, и постоянной, когда в силу отсутствия прежней кормовой породы, преобладания новой или под влиянием каких-либо других экологических условий насекомое приспосабливается к новой пище. Такое приспособление ведет к известному сдвигу в обмене веществ и образованию новых пищевых форм насекомых. Временная вынужденная смена кормовой породы наблюдается в природе очень часто при массовых размножениях, когда обычного корма не хватает. При этом гусеницы многих бабочек начинают питаться малопригодными растениями, в том числе и травянистыми, что приводит к резкому снижению выживаемости популяции. У монофагов такой смены не наблюдается, они в массе гибнут, но не переходят на питание несвойственными им растениями.
Явления регулярной смены кормовых пород вполне закономерны для некоторых насекомых, например для тлей семейства Phylloxeridae при чередовании половых и девственных поколений.
Смена кормовых пород может наблюдаться в случае попадания вида в новые области, где отсутствуют обычные для него древесные породы, или при введении новой древесной культуры в районе, где она раньше не произрастала. В этих случаях смена кормовой породы вынужденная; она может быть как временной, так и постоянной.
Влияние питания на рост, развитие, плодовитость и выживаемость лесных насекомых
Количество и качество съеденной пищи оказывает влияние на физиологическое состояние насекомых и находится в прямой связи с биологическими показателями их развития. Считается, что наиболее предпочитаемые насекомыми древесные породы являются для них самыми питательными. На этих породах насекомые быстрее заканчивают полный цикл развития, дают более плодовитое потомство и максимальную выживаемость. В пределах предпочитаемых пород показатели могут изменяться: одна древесная порода обеспечивает большую плодовитость и выживаемость, на другой достигается быстрейшее развитие всех фаз насекомого. Так, гусеницы сибирского шелкопряда питаются всеми хвойными породами сибирской тайги, но скорость их развития, средняя масса куколок, плодовитость и соотношение самцов и самок различны (табл.).
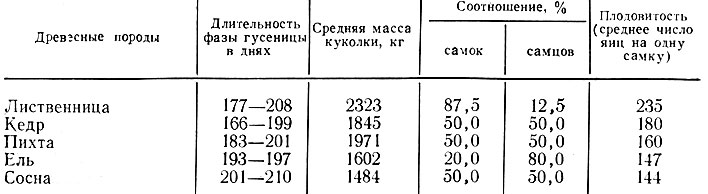
Таблица. Средние показатели развития сибирского шелкопряда на разных древесных породах (по Н. Г. Коломийцу и В. О. Болдаруеву)
Из таблицы видно, что лучше всего гусеницы растут и развиваются на хвое лиственницы, а хуже - на хвое ели и сосны. Условия питания гусениц отражаются и на массе куколок, которая связана с плодовитостью бабочек.
Другой иллюстрацией влияния пищи на рост и развитие насекомых являются многочисленные исследования по непарному шелкопряду. В лесостепной зоне развитие гусениц лучше всего протекает на дубе; на других породах оно отстает на 2-10 дней, особенно на липе и березе. Куколки имеют наибольшую массу в том случае, если гусеницы выкармливались на предпочитаемых в данном районе древесных породах. В условиях лесостепи самая большая масса куколок на дубе и яблоне (0,8 г), а минимальная - на липе (0,5 г). Плодовитость пропорциональна массе куколок и выражается прямолинейной зависимостью вида у = ах + b (рис. 22). Кроме того, гусеницы, выросшие на дубе и яблоне, дают больше самок, чем самцов, и обладают наибольшей выживаемостью.
В лесной зоне (около Москвы) гусеницы непарного шелкопряда предпочитают березу; соответственно меняются и все показатели развития.
Интересные связи с кормовыми породами обнаруживает майский хрущ, личинки которого питаются корнями древесных пород, а жукам необходимо дополнительное питание на листьях древесных пород. Развитие личинок интенсивнее проходит на корнях сосны, богатых углеводами и бедных азотистыми веществами. Отродившимся жукам, наоборот, требуется дополнительное белковое питание, им необходим азот, содержащийся больше всего в листьях дуба. Поэтому питание листьями дуба увеличивает плодовитость жуков. Очевидно, что сочетание питания личинок корнями сосны, а жуков - листьями дуба дает потомство, обладающее наибольшей жизненностью (рис. 23).
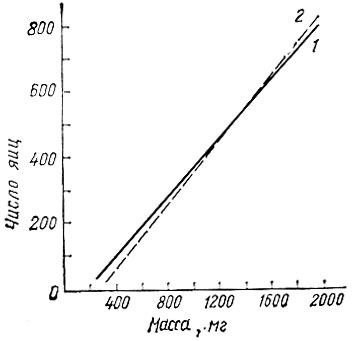
Рис. 22. Зависимость между массой куколок и числом яиц в кладке непарного шелкопряда (по Рудневу, 1951): 1 - теоретическая, 2 - экспериментальная
Влияние пищи сказывается и на вредителях, питающихся древесиной. Так, плодовитость желтопятнистого усача на тополе в среднем составляла 36,6 яйца, а на ольхе - 7,0; продолжительность развития фазы куколки на тополе 10,7 дня, а на козьей иве - 42,3 дня (Старк, 1955). Особенно тонко реагируют личинки стволовых вредителей на влажность тканей дерева. При ее недостатке личинки черного соснового усача впадают в диапаузу и развитие их затягивается на год (Кузнецова, 1957).
Лесные насекомые выносливы к голоданию, однако недостаток пищи вызывает резкое снижение плодовитости и выживаемости, сказывается на размерах тела насекомых и сроках развития. Так, например, гусеница сибирского шелкопряда для полного развития должна съедать пищи в 1500 раз больше ее первоначальной массы. При недостатке пищи рост и развитие гусениц задерживаются и сопровождаются большой смертностью. Голодающие гусеницы быстрее окукливаются, а вышедшие из куколок бабочки бывают мелкие и откладывают меньше яиц.
Опыты с голоданием гусениц пихтовой пяденицы показали, что они живут без пищи в первом возрасте в среднем четыре дня, а в старших возрастах восемь дней (Прозоров, 1955).
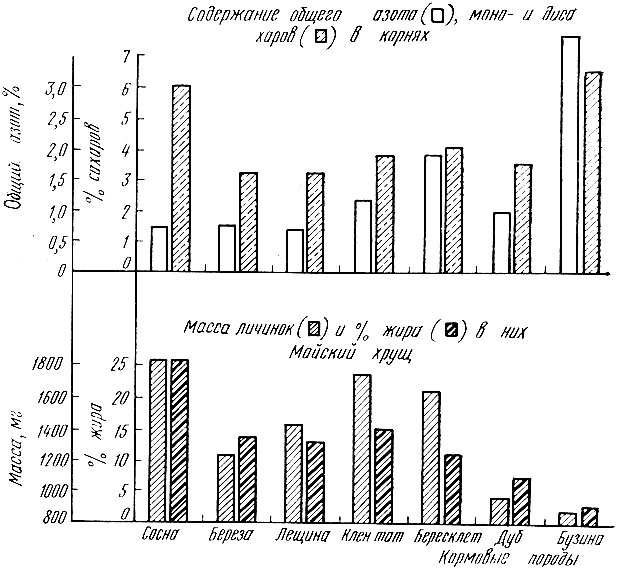
Рис. 23. Влияние вида древесных и кустарниковых пород на личинок майского хруща (по Березиной, 1957)
Бабочки кольчатого шелкопряда, полученные от голодавших четверо суток гусениц, дали на 36,5% меньше яиц, чем неголодавшие (Щербиновский, 1925).
Многие жуки, особенно хищные, могут существовать без пищи 7 - 10 дней, а в отдельных случаях целый месяц.
Повреждение древесных пород насекомыми
Различные древесные породы повреждаются не одинаково. Это зависит от их устойчивости и числа обитающих на них видов насекомых. Так, например, в СССР на дубе встречается около 850 видов насекомых, на тополях - около 700, на ильмовых - более 300 видов. Наиболее богата видами энтомофауна большинства букоцветных, ивоцветных и розоцветных. Меньше опасных вредителей встречается на кленах, ольхе саксауле и интродуцированных породах. Так, например, произрастающие в наших парках гледичия, софора, мыльное дерево, бархат черемуха Маака, катальпа почти не заселяются вредителями. На лиственных породах обитает много полифагов, связанных с рядом растущих совместно древесных пород и кустарников. В первые годы жизни они часто повреждаются насекомыми, обитающими на окружающей травянистой растительности.

Рис. 24. Главнейшие типы повреждений листьев древесных пород: А - Б - грубое объедание. В - скелетирование, Г - минирование, Д - скручивание, Е - галлы
Энтомофауна хвойных пород специализирована больше и представлена преимущественно олигофагами, не переходящими на лиственные породы. Наиболее богата видами фауна сосны, затем ели, пихты, кедра, лиственницы. Очень бедна фауна тисса, туи, кипариса, можжевельника обыкновенного.
Типы повреждений растений насекомыми
Насекомыми повреждаются все органы растений. Повреждения носят разнообразный характер. Они могут быть внутренними и внешними.
Примером внутренних повреждений растений служат ходы личинок насекомых внутри дерева, под корой, в лубе и древесине. Эти ходы могут иметь правильную форму геометрической фигуры (у короедов) а могут быть лишены такой формы и носить характер различных выгрызов, не представляющих определенной фигуры.
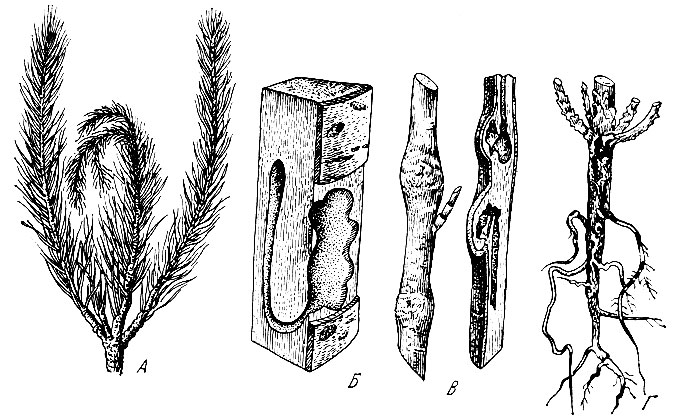
Рис. 25. Главнейшие типы повреждений стволов древесных пород: А - искривление побегов, Б - ходы в древесине, В - вздутие побегов, Г - ходы на корнях
Внешние повреждения растений очень разнообразны. Наиболее типичны повреждения листьев и коры. Личинки насекомых объедают листья с краев, выгрызают дырки внутри листьев, прокладывают в паренхиме ходы (минирование), выедают мякоть листа с оставлением сети жилок (скелетирование), свертывают листья с помощью паутины, выделяемой личинками (листоверток, молей и других насекомых), или скручивают в трубочки и "сигары" (жуки трубковерты) (рис. 24).
Под влиянием насекомых на стеблях (ветках и стволе) и листьях образуются наросты, опухоли и различные вздутия (рис. 25).
Разрастания тканей, возникшие в результате сосания насекомых и имеющие постоянную величину и форму, появляющуюся только при повреждении определенными видами насекомого, называются галлами. Галлы бывают сплошными и полыми, в последнем случае насекомые находятся внутри галлов и покидают их только после образования трещин. Галлы бывают очень разнообразны.
Многих насекомых очень легко определить по наносимым ими повреждениям1.
1 (Подробнее см.: Воронцов А. И., Мозолевская Е. Г. Практикум по лесной энтомологии, 2-е изд. М., 1978.)
Устойчивость древесных пород к вредным насекомым
является специфическим проявлением их взаимоотношений, сложившихся в процессе эволюции. Она зависит от многих причин. Обычно выделяют три основных фактора устойчивости: предпочтение и отсутствие продпочтения, анабиоз, выносливость (Пайнтер, 1953).
Предпочтение или отсутствие предпочтения той или иной древесной породе в качестве пищи, места для откладки яиц или укрытия определяется поведением самого насекомого и тесно связано с его пищевой специализацией.
Антибиоз - это отрицательное действие, оказываемое устойчивым деревом или породой дерева на определенные фазы жизненного цикла питающегося на нем насекомого. Такое действие может проявляться в снижении плодовитости или в увеличении смертности.
Под выносливостью понимается способность древесных пород восстанавливать поврежденные органы и только незначительно снижать текущий прирост по высоте. Например, черный тополь активно заселяется зеленой узкотелой златкой-, а бальзамический - темнокрылой стеклянницей. Однако эти древесные породы выносливы к повреждениям. Они быстро образуют каллюс и заращивают возникшие при повреждении раны. Текущий прирост этих деревьев изменяется мало. То же относится и к случаю повреждения листьев на деревьях. Наименее выносливы хвойные породы, гораздо больше - лиственные. Из хвойных пород самыми выносливыми являются сосна обыкновенная и лиственницы.
Биотические факторы
Жизнь насекомых в лесу обусловлена многообразными, часто очень сложными, а порой и противоречивыми связями с другими организмами и между собой. Их можно разделить на межвидовые и внутривидовые отношения.
Межвидовые отношения
У насекомых межвидовые отношения обычно проявляются в виде симбиоза, паразитизма, хищничества и конкуренции.
Симбиоз - это устойчивые, разного свойства тесные взаимосвязи организмов различных видов, выгодные одному или обоим видам. Выделяют два типа симбиоза: мутуализм, при котором оба организма извлекают пользу от совместного существования; комменсализм, при котором взаимоотношения для одного вида полезны, а для другого большей частью нейтральны. Оба типа симбиоза широко распространены в природе. Так, муравьи питаются сахаристыми выделениями тлей и кокцид (некоторые виды червецов), очищают при этом их колонии от клейких экскрементов и защищают от врагов (хищников и паразитов). Многие насекомые, питающиеся древесиной, заносят в ходы грибы, которые там развиваются и способствуют разложению клетчатки на составные химические соединения, которыми питаются насекомые - ксилофаги. Существуют мутуалистические отношения между термитами и обитающими в их кишечнике одноклеточными организмами.
Особенно многочисленны примеры комменсализма. Так, например, ряд насекомых прикрепляется друг к другу для передвижения (форрезия). Иногда насекомые одного вида живут за счет запасов другого, не причиняя ему существенного вреда (нахлебничество). Многие насекомые укрываются в гнездах муравьев, термитов, златогузки.
Паразитизм - это односторонне выгодное использование одним организмом другого организма в качестве источника пищи и среды обитания на протяжении большей или меньшей части своего жизненного цикла. Существуют первичные паразиты, которые в качестве хозяев используют фитофагов, и вторичные, или гиперпаразиты (сверхпаразиты); они используют в качестве хозяев других паразитов. В одном хозяине может развиваться одна или несколько особей паразита. Бывают случаи, когда одного хозяина заражает несколько видов паразитов (мультипаразитизм). Формы паразитизма очень разнообразны. Среди насекомых существует много видов самых разнообразных паразитов. Большинство из них относится к отрядам перепончатокрылых и двукрылых (см. гл. XI).
Хищничество отличается от паразитизма в основном тем, что жертва сразу погибает от нападающего на нее хищника. При этом один хищник может уничтожить много жертв. В контакте с каждой жертвой он проводит ограниченное время. Хищные насекомые охотятся за гусеницами бабочек, истребляют личинок жуков, различных куколок и даже взрослых насекомых. Они ловят свою жертву, убивают ее и затем поедают. Типичные хищники: жужелицы, рыжие лесные муравьи, мухи-ктыри, песчаные осы и другие насекомые. Хищниками являются также насекомоядные птицы, летучие мыши, бурозубки и другие представители млекопитающих из отряда насекомоядных.
Численность хищника (паразита) и численность его жертвы (хозяина) тесно связаны между собой. Некоторые исследователи доказали, что при неизменных условиях численность жертвы увеличивается постепенно в течение ряда поколений, а затем падает; то же самое происходит с численностью хищника, но с некоторым запаздыванием. Когда жертва имеется в изобилии, то численность хищника возрастает, что в конечном итоге приводит к сокращению численности жертвы и в свою очередь вызывает уменьшение хищника, после чего численность жертвы может снова возрастать и цикл повторяется. На рис. 26 графически изображены колебания численности одного и того же вида хозяина (фасолевая зерновка), преследуемого медленно (вверху) и быстро (внизу) воспроизводящимся паразитом. При быстром размножении паразита он стабилизирует популяцию жертвы.
Конкуренция между разными видами насекомых возникает в том случае, когда сходные потребности к условиям жизни полностью не удовлетворяются. Так, если на дубе одновременно развивается большое число гусениц нескольких видов чешуекрылых, питающихся листьями, и этих листьев всем не хватает, между гусеницами складываются конкурентные отношения. Очень часто популяция вида, чьи гусеницы отрождаются в момент распускания листьев, вытесняют гусениц других видов, появляющихся позднее, так как им уже не остается пищи.
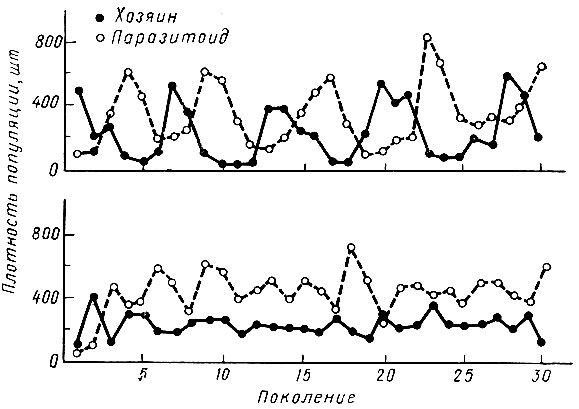
Рис. 26. Колебание плотности популяции паразита и хозяина в результате их взаимодействия (по Утида, 1957)
Очень ярко проявляется межвидовая конкуренция у хищных насекомых. Например, муравей черный лазиус (Lasius niger) всегда вытесняет с занятой им территории желтого лазиуса (Lasius flavus).
Конкурентные отношения между видами оказывают большое влияние на характер размещения и перемещения насекомых по территории.
Внутривидовые отношения
Особи в популяциях одного вида насекомого связаны различными по характеру и значению взаимоотношениями.
Отдельные особи и группы особей одного и того же вида могут воздействовать друг на друга прямо и активно или же косвенно, через изменение окружающей среды. Результаты таких отношений для отдельно взятой особи или группы особей могут быть весьма различны. Они могут стимулировать протекание жизненных процессов (рост, размножение, развитие) или ослаблять их. На большинство форм внутривидовых отношений определенное влияние оказывает численность участвующих в них особей. Регулирование численности достигается рядом внутривидовых механизмов. Типичным внутривидовым приспособлением является переход саранчи от одиночного к стадному образу жизни и последующие миграции стай. Гусеницы многих бабочек, питающиеся листвой древесных пород, живут в первых возрастах обществами, а затем расходятся и переходят к одиночному образу жизни, избегая таким образом конкуренции за пищу. В период же коллективного питания гусениц интенсивность последнего стимулировалась специфической средой их совместного обитания.
Характерным внутривидовым приспособлением служат гнезда у пушистого и кольчатого коконопрядов. В этих гнездах живет по нескольку сотен гусениц. Ночью температура гнезда на 8-12° С выше, чем окружающего воздуха. Это позволяет гусеницам легко перенести низкие ночные температуры.
Существует связь между размерами листа тополя и количеством откладываемых на него яиц тополевой молью. С увеличением размера листа возрастает и число яиц. Автоматическое регулирование численности происходит также у щитовок. При очень густом поселении одни особи вытесняют механически другие. Вытесненные особи не получают питания и погибают или голодают и становятся бесплодными.
При очень большой плотности популяции обычно возникает острая конкуренция между особями за пищу. В результате резко возрастает смертность, развиваются эпизоотии и численность автоматически сокращается, вновь увеличиваясь через определенный промежуток времени.
Насекомые как элемент экосистемы
Выше говорилось, что первичной элементарной структурной единицей биосферы является экосистема. Обычно ставят знак равенства между экосистемой и биогеоценозом. Однако есть некоторые расхождения между пониманием экосистемы в зарубежных работах и трудах В. Н. Сукачева (автора учения о биогеоценозах) и его школы.
В отличие от понятия экосистемы у зарубежных авторов, согласно которому экосистема может охватывать пространство любой протяженности - от капли прудовой воды и до Вселенной, - биогеоценоз имеет строго определенный объем. Биогеоценозы (экосистемы) являются частями земной или водной поверхности, однородной по топографическим, микроклиматическим, почвенным, гидрологическим и биотическим условиям.
В современной экологической литературе применяется термин экосистема. Он краткий и международный. Этот термин и будет использован в дальнейшем.
Экосистема относительно устойчива во времени и термодинамически открыта в отношении притока и оттока веществ в атмосферу. Экосистемы сложились в процессе длительной эволюции, они являются слаженными, устойчивыми механизмами, способными путем саморегулирования противостоять как изменениям в среде, так и изменению в численности организмов.
Все взаимодействия компонентов экосистемы основаны в конечном счете на обмене веществом и энергией между ними. Основным источником энергии в экосистемах Земли служит Солнце. На Земле энергия солнечного излучения включается в биологический круговорот только через фотосинтез, осуществляемый автотрофными организмами. Кванты видимого света, проходя через экосистему, поглощаются хлорофиллом зеленых клеток растений. В процессе фотосинтеза автотрофные растения поглощают из атмосферы углекислый газ, из почвы - воду и минеральные вещества и с помощью энергии Солнца создают из них сложные органические соединения (углеводы, белки, жиры, органические кислоты и др.). Эти вещества богаты энергией и способны к различным химическим превращениям как в самих растениях, так и в телах различных гетеротрофов, использующих энергию растительных материалов в процессе питания на свои жизненные функции.
Количество солнечной энергии, поступающей в экосистемы, ничтожно по сравнению с общим энергетическим бюджетом Земли. Всего около 0,1% энергии, получаемой от Солнца связывается в процессе фотосинтеза. Однако за счет этой энергии может синтезироваться несколько тысяч граммов сухого органического вещества на 1 м2 в год. Более половины энергии, связанной при фотосинтезе, тут же расходуется в процессе дыхания самих растений. Часть же ее переносится через посредство ряда организмов по пищевым или трофическим цепям экосистемы.
Пищевые цепи возникают как результат сложных пищевых взаимоотношений между растительными и животными организмами. Они объединяют прямо или косвенно большую группу организмов в единый комплекс.
Цепь питания обычно состоит из нескольких звеньев. Первое звено образуют продуценты, или производители. Это автотрофные растения. Они создают первичную биологическую продукцию и аккумулируют солнечную энергию.
Второе звено представлено консументами (консумцио - потребляю), или потребителями, - гетеротрофными организмами. Различают консументы первого порядка - фитофаги, второго порядка - плотоядные животные, третьего порядка - хищники, питающиеся другими животными, и т. д.
Наряду с консументами существуют еще редуценты (редуцирую - уменьшаю), или деструкторы, которые разрушают и поедают мертвое органическое вещество и минерализуют его до простых неорганических соединений. К ним относится большая группа сапрофагов, в которую входят грибы, микроорганизмы, многие членистоногие и др.
В каждой цепи формируются определенные трофические уровни, характеризующиеся различной интенсивностью протекания потока вещества и энергии. Следовательно, зеленые растения (продуценты) занимают первый трофический уровень, фитофаги размещаются на втором уровне.
Необходимо подчеркнуть, что эта трофическая классификация основывается на функционировании, а не на видовом составе животных и растений как таковом.
Начиная со звена продуцентов можно наметить два основных пути использования энергии. Во-первых, она может использоваться травоядными животными (фитофагами), поедающими непосредственно живые ткани растений; во-вторых, ее могут потреблять сапрофаги в виде уже отмерших тканей (например, при разложении лесной подстилки). В соответствии с этим существует два вида пищевых цепей: цепи выедания и цепи разложения. Энергия может запасаться на длительное время в цепях обоих видов, но соотношение ее потоков, поступающих в эти цепи для разных экосистем, неодинаково. В наземных экосистемах в цепи выедания (к фитофагам) поступает только очень небольшая часть (не более 10%) энергии, связанной с растениями, основная же часть (90%) используется в пищевых цепях разложения (рис. 27).
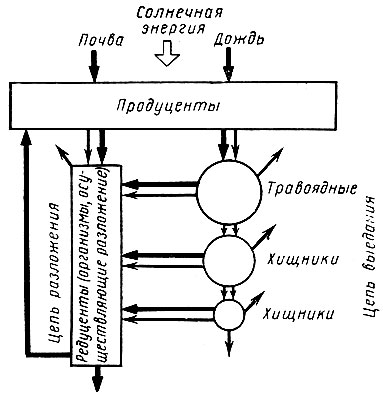
Рис. 27. Поток энергии (тонкие стрелки) и питательных веществ через экосистему (по Вудвеллу, 1972)
Следует подчеркнуть, что пищевые цепи разложения не менее важны, чем цепи выедания. На суше эти цепи начинаются с мертвого органического вещества (листьев, коры, ветвей), в воде - с отмерших водорослей, фекальных масс и других органических остатков. Органические остатки могут полностью потребляться бактериями, грибами и мелкими животными (в том числе насекомыми) - сапрофагами; при этом выделяются углекислый газ и тепло. Цепи выедания и разложения тесно переплетаются между собой, так как мертвое органическое вещество может поступать в ложные пищевые цепи, потенциально включающие более крупных животных, потребляющих это вещество. Эти животные в свою очередь потребляются еще более крупными хищниками. При переносе энергии с одного трофического уровня на другой значительная ее часть деградирует в тепло, как этого требует второй закон термодинамики. Чем короче пищевая цепь или чем ближе данный организм к началу пищевой цепи, тем пищевая энергия более доступна.
Экологическая эффективность экосистемы определяется отношением величины ассимиляции на данном уровне трофической цепи к величине ассимиляции на предыдущем уровне, выраженном в процентах. Эта эффективность, как говорилось выше, всегда очень низкая. Если расположить один под другим прямоугольники, длина которых пропорциональна потоку энергии каждого уровня, то получится экологическая пирамида. Высота пирамиды будет соответствовать длине пищевой цепи (рис. 28).
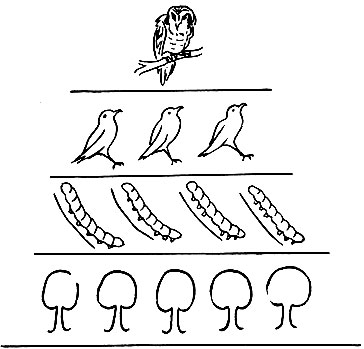
Рис. 28. Экологическая пирамида, представляющая очень простую трофическую цепь: дуб - гусеницы непарного шелкопряда - насекомоядные птицы - хищные птицы
В каждой лесной экосистеме насекомые представлены большим числом видов и особей. В процессе обмена веществом и энергией они играют различную роль в трофических цепях. В цепях выедания преобладают фитофаги, потребляющие различные ткани растений, и их энтомофаги, а в цепях разложения - детритофаги, перерабатывающие растительный опад, трупы животных и их экскременты. Каждый вид в экосистеме занимает определенное местообитание и выполняет определенную работу, говоря образно, имеет свой адрес и профессию. В соответствии с этим существуют понятия стации и экологической ниши.
Стация, или местообитание вида (особи или целой популяции вида), - это место, где он живет, или место, где его обычно можно найти. В пределах стации популяция вида (или ее отдельные особи) может проходить весь жизненный цикл или только часть его. Так, например, излюбленная стация обитания восточного майского хруща - свежие песчаные почвы, заросшие вейником, или закультивированные сосной. В то же время взрослые жуки, выходя из почвы, улетают в березовые или дубовые насаждения, где проходят их дополнительное питание и спаривание.
Экологическая ниша - понятие более емкое, включающее в себя не только физическое пространство, занимаемое организмом, но и функциональную роль организма в экосистеме. Следовательно, ниша - это термин, употребляемый для обозначения специализации популяции вида внутри экосистемы. Современные экологи придают очень большое значение изучению экологических ниш и описывают их с помощью количественных характеристик, оперируют с ними математически (Федоров, 1980). Автор широко известной монографии по экологии Ю. Одум указывает, что знание местообитания - это только начало в изучении организмов. Чтобы определить положение организма в экосистеме, необходимо располагать сведениями о его активности, в частности о питании и источниках энергии, о скоростях метаболизма и роста, о его влиянии на другие организмы, с которыми он вступает в контакт, и о тех пределах, в которых он влияет или способен влиять на важные процессы в экосистеме. В лесной энтомологии учение об экологической нише почти не используется. Лесоводы пока довольствуются изучением приуроченности популяции видов лесных насекомых к определенным, местообитаниям (стациям) и типам леса (рис. 29), которые в широком понимании являются экосистемами.
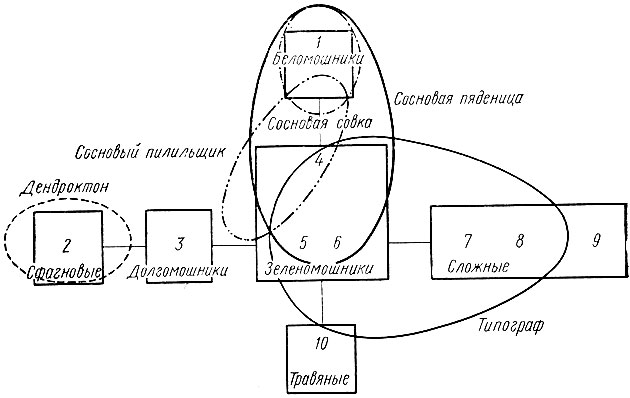
Рис. 29. Распространение некоторых видов вредных насекомых по типам леса: 1 - сосняк лишайниковый, 2 - сосняк сфагновый, 3 - сосняк долгомошник, 4 - сосняк черничник, 5 -сосняк брусничник, 6 -сосняк кисличник, 7 - сосняк липовый, 8 - сосняк лещиновый, 9 - сосняк дубняковый, 10 - сосняк травяной
Стация всегда характеризуется определенным режимом подпологовой среды, который отвечает всей сумме факторов, необходимых для жизни популяции данного вида. При этом на разных фазах развития насекомого требования к условиям подпологовой среды могут меняться, что обусловливает динамичность насекомых в лесу, их постоянные перемещения и мозаичность распределения в насаждениях. Знание стациального распределения отдельных видов лесных насекомых дает возможность их быстро обнаружить, вести постоянные наблюдения за их численностью и управлять ею путем лесохозяйственных мероприятий, изменяющих среду обитания.
Особенно четко выявляется стациональное размещение почвообитающих насекомых. Механический состав почвы, ее влажность и температура, состав и толщина лесной подстилки и напочвенный травяной покров определяют стации обитания майского хруща, проволочников, ложнопроволочников и других вредителей корней.
Свойство видов избирательно заселять те или иные стации - очень важная экологическая закономерность. Это свойство получило название принципа стациальной верности. Однако этот принцип оказывается действительным лишь для определенного географического района, где экологические условия более или менее однородны.
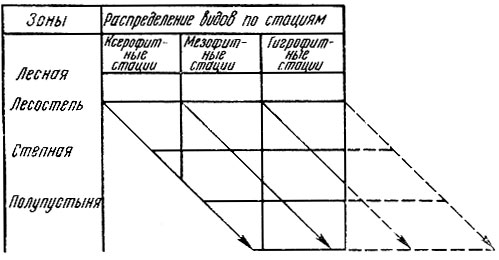
Рис. 30. Диаграмма зональной смены стаций (по Бей-Биенко)
При изменении условий применительно к другим ландшафтным зонам виды меняют свои местообитания. Так, например, восточный майский хрущ в зоне тайги поселяется только на открытых местах, а в степной зоне живет под пологом сомкнувшихся насаждений. Такое явление получило название зональной смены стаций и впервые было сформулировано крупным советским энтомологом Г. Я. Бей-Биенко (1904 - 1971). Проявляется зональная смена стаций как закономерно направленное изменение местообитаний при переходе вида из одной природной зоны в другую: при продвижении к северу избираются более сухие, хорошо прогреваемые открытые стации с разреженным растительным покровом, часто располагающиеся на легких песчаных или каменистых почвах. При продвижении к югу тот же вид заселяет более увлажненные и тенистые стации с густым растительным покровом и с глинистыми почвами (Бей-Биенко, 1971). Графически принцип зональной смены стаций показан на рис. 30.
Ранее указывалось, что видовой состав вредных лесных насекомых изменяется по типам леса и стациям, а равно и с возрастом насаждений. Каждая лесная экосистема подвержена возрастным изменениям, получившим название сукцессий. Если первый этап охватывает период жизни деревьев от появления их из семян до смыкания кронами и образования древостоя, второй наступает после смыкания деревьев кронами и длится до окончания интенсивного изреживания, третий этап - это период средневозрастных и спелых насаждений, а четвертый характеризуется старением древостоя. Каждому периоду свойственны определенные энтомокомплексы дендрофильных насекомых (Воронцов, 1963).
По мере разрушения древостоя под влиянием самых различных причин (пожары, рекреационное воздействие, ветровалы и т. д.) происходит заселение деревьев насекомыми и грибами, комплексы которых последовательно сменяются под влиянием изменения физических свойств древесины. Наблюдается типичная сукцессия, хорошо изученная на многих древесных породах Б. М. Мамаевым и сотрудниками (рис.31).
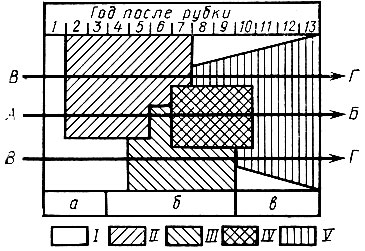
Рис. 31. Сукцессия разрушающих древесину беспозвоночных животных в различных местообитаниях в сравнении с фазами ее разрушения грибами (по Мамаеву, 1960): Стрелки - общее направление сукцессии: ВГ - укороченная сукцессия, I - бактериально-грибное разложение, II - церамбицидная стадия, III - луканидная стадия, IV - форми-цидная стадия, V - лумбрицидная стадия; a - фаза деревоокрашивающих грибов, б - фаза дереворазрушающих грибов, в - фаза шляпных грибов
Динамика численности популяции
В экологии насекомых центральное место занимает динамика численности популяций. Плотность определяется числом особей, приходящихся на единицу площади или объема. Методы определения плотности популяции разнообразны и зависят от экологии особей данного вида и тех биотопов, которые она населяет. Характер распределения особей, составляющих популяцию, в пространстве может быть равномерным, случайным и скученным. Знание типа распределения особей в популяции важно при оценке ее плотности методом выборки.
Возрастной состав популяции имеет очень большое значение для ее существования и процветания. При благоприятных условиях в популяции присутствуют все возрастные группы и поддерживается более или менее стабильный состав. В быстро растущих популяциях доминируют интенсивно размножающиеся молодые особи, а в сокращающихся - старые, уже неспособные интенсивно размножаться. Такие популяции малопродуктивны, недостаточно устойчивы. На возрастной состав популяции большое влияние оказывает продолжительность жизни особей, период достижения ими половой зрелости, число генераций. Очевидно, что у многих насекомых, дающих по нескольку генераций в год, возрастная структура популяции очень простая, а у долгоживущих - очень сложная.
Рождаемость в популяции определяется прежде всего эволюционным положением вида, его биологией. Низкая плодовитость характерна для тех видов, которые проявляют большую заботу о потомстве. Кроме того, рождаемость зависит от скорости полового созревания, числа генераций в году, соотношения в популяции самцов и самок, обеспеченности кормом, влияния погодных условий.
Смертность в популяции - это количество особей, погибших за определенный период. Она бывает очень высокой и изменяется в зависимости от условий среды, возраста и состояния популяции. У большинства видов смертность в раннем возрасте всегда бывает выше, чем у взрослых особей. Однако встречаются и такие виды, у которых смертность приблизительно одинакова во всех возрастах или преобладает у особей старших возрастов. Факторы смертности очень разнообразны. Она может быть вызвана влиянием физических условий (низкие и высокие температуры, ливневые осадки и град, избыточная и недостаточная влажность и др.), биотическими факторами (отсутствие благоприятного корма, инфекционные заболевания, враги и т. д.) и антропогенными (загрязнение окружающей среды, уничтожение насекомых, вырубка деревьев и др.).
Рост популяций определяется двумя противоположными явлениями - рождаемостью и смертностью, также миграциями и иммиграциями. Большинство исследователей полагает, что рождаемость - более постоянная, хотя и очень изменчивая величина, чем смертность. Поэтому и уровень численности зависит главным образом от факторов смертности.
Баланс популяции изучают, пользуясь таблицами выживания, которые дают возможность оценить сравнительное значение каждого фактора смертности в отдельности и выделить критические периоды в течение генерации, определяющие дальнейший ход динамики численности. В таблицах описана серия последовательных измерений, с помощью которых можно обнаружить изменение численности популяции в естественных условиях1.
1 (Техника построения таблиц выживания описана в книге А. И. Воронцова)
Каждому виду присуща определенная оптимальная плотность популяции, отклонения от которой в обе стороны отрицательно сказываются на темпах воспроизводства и жизнедеятельности особей. Колебания численности вокруг оптимального уровня носят разный характер. Они могут быть плавными циклическими и резкими нерегулярными. Механизм этих колебаний очень сложен и до сих пор трудно прогнозируем.
Для объяснения причин колебания численности насекомых был предложен ряд теорий, не оправдавшихся в своем первоначальном виде. Одни исследователи признавали ведущую роль в регуляции численности фитофагов за их врагами - хищниками и паразитами (паразитарная теория), другие считали, что колебания численности обусловлены воздействием на насекомых климатических факторов (климатическая теория). Ряд исследователей отдавал преимущество пищевому фактору. По мере дальнейшего развития исследований стали считать, что численность популяции каждого вида насекомого в экосистеме регулируется комплексом факторов (биоценологическая теория). В настоящее время наибольшее признание получила синтетическая теория. Она рассматривает колебания численности популяций как автоматически регулируемый процесс, слагающийся из отклонений под влиянием случайных воздействий абиотической среды и стабилизирующего действия биотических факторов. На рис. 32 показана общая схема динамики численности популяций насекомых. Существует два принципиально различных процесса, определяющих движение численности популяций во времени: модификация и регуляция. Модификация выражает подъемы и спады численности, обусловленные случайными по отношению к популяциям изменениями факторов, функционально не связанных с ее плотностью. Это в первую очередь погодные условия. Они имеют решающее значение при выяснении причин колебания численности и прогнозировании массовых размножений насекомых.
Аномальное развитие отдельных метеорологических элементов может почти мгновенно и очень резко изменить уровень численности популяции. Погодные условия влияют с равной интенсивностью при любой плотности популяции. Так, при очень низкой или очень высокой температуре, ливне или наводнении смертность будет неизбирательной и не будет зависеть от числа особей, она, как правило, будет массовой, и сохранятся лишь отдельные особи в надежных укрытиях. Подъем численности популяции часто бывает обусловлен устойчивым типом погоды, например длительной засухой.
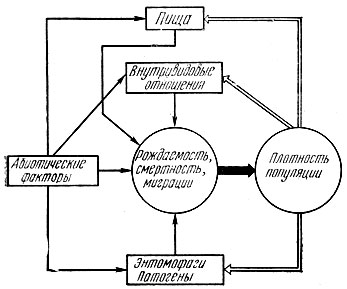
Рис. 32. Общая схема динамики численности насекомых
Еще чаще метеорологические условия оказывают косвенное влияние на численность популяции, усиливая смертность через другие факторы. Под влиянием погодных условий сезонные изменения развития кормовых пород могут совпадать с появлением определенных фаз данного вида насекомого или не совпадать. Так, например, в один год выход гусениц дубовой зеленой листовертки совпадает с распусканием почек дуба, а в другие годы наступает раньше, когда почки еще не раскрылись. В этом случае происходит массовая гибель гусениц. Под влиянием выпадающих осадков колеблется интенсивность смоловыделения у сосны. В годы засухи смоляное давление в хвое и почках падает, что ведет к резкому увеличению выживаемости гусениц ряда хвоегрызущих насекомых.
Регуляция численности насекомых заключается в сглаживании возникающих колебаний в результате действия регулирующих механизмов, которые зависят от плотности популяции и действуют по принципу обратной связи. Регуляторные механизмы объясняют частоту и интенсивность колебаний численности насекомых. На рис. 33 показаны основные механизмы регуляции численности популяций насекомых. Различают межвидовые и внутривидовые механизмы регуляции численности. К первым относятся паразиты, хищники и патогены. Многоядные энтомофаги (муравьи, птицы и др.) стабилизируют численность популяции на самых низких уровнях, специализированные паразиты регулируют ее на более высоких уровнях. Резкое снижение численности насекомых около ее высшего предела осуществляют болезни насекомых, вызывающие эпизоотии. Если они отсутствуют или действуют недостаточно, включаются внутривидовые регулирующие механизмы. Они являются результатом взаимодействия особей внутри самой популяции. В простейшем виде - это разнообразные формы внутривидовой конкуренции, связанные с прямым и косвенным отрицательным воздействием особей друг на друга.
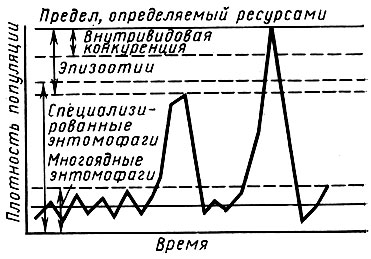
Рис. 33. Пороги и зоны активности основных механизмов регуляции численности насекомых (по Г. А. Викторову, 1974)
Большую роль в регуляции численности популяции играют миграции. С помощью миграций идет убыль и пополнение популяций, может резко изменяться их численность, происходить перемещение в пространстве и накопление особей в новых местах. Миграции позволяют популяции расселяться и избежать неблагоприятных условий, которые создаются в результате ее же жизнедеятельности, сохраняться в период депрессии в резервациях и затем быстро накапливаться в соседних с ними насаждениях.
Численность популяции может изменяться также и под влиянием физиологической разнокачественности особей. Например, вследствие такой разнокачественности ежегодно часть особей рыжего пилильщика, дубовой хохлатки и других вредителей леса уходит в диапаузу на фазе пронимфы и куколки, а остальные особи заканчивают нормальное развитие по одногодичному циклу. За счет этого в одних генерациях происходит быстрое сокращение, а в других - увеличение численности популяции.
Литература
Беклемишев В. Н. Биоценологические основы сравнительной паразитологии. М., 1970.
Викторов Г. А. Проблемы динамики численности насекомых на примере вредной черепашки. М., 1967.
Воронцов А. И. Биологические основы защиты леса. М., 1963.
Дажо Р. Основы экологии. М., 1975.
Дарвин Ч. Происхождение видов (1859). М.- Л., 1935.
Дрё Ф. Экология. М., 1976.
Кашкаров Д. Н. Основы экологии животных. Л., 1944.
Кожанчиков И. В. Методы исследования экологии насекомых. М., 1961.
Макфедьен Э. Экология животных (цели и методы). М., 1965.
Новиков Г. А. Очерк истории экологии животных. Л., 1980.
Одум Ю. Основы экологии. М., 1975.
Радкевич В. А. Экология. Минск, 1977.
Риклефс Р. Основы общей экологии. М., 1979.
Смит Дж. М. Модели экологии. М., 1979.
Сукачев В. Н. и др. Основы лесной биоценологии. М., 1964.
Тимофеев-Ресовский Н. В., Яблоков А. В., Глотов Н. В. Очерк учения о популяции. М., 1973.
Уатт К. Экология и управление природными ресурсами. М., 1971.
Ульямсон М. Анализ биологических популяций. М., 1975.
Федоров В. Д., Гильманов Т. Г. Экология. М., 1980.
Фридерикс К. Экологические основы прикладной зоологии и энтомологии. М - Л., 1932.
Элтон Ч. Экология животных. М., 1934.
Яхонтов В. В. Экология насекомых, 2-е изд. М., 1969.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'