Глава I. Вопросы общей энтомологии
Место насекомых в системе животного мира
Насекомые составляют один из семи классов типа членистоногих животных. К этому типу относятся также ракообразные, многоножки и паукообразные. Представители этих классов широко распространены в природе. В лесах наряду с насекомыми всегда встречаются многоножки и паукообразные. Ракообразные живут преимущественно в воде. Тип членистоногих характеризуется делением всего тела на отдельные членики, или сегменты, наличием твердого наружного покрова, выполняющего роль скелета, парными сочлененными конечностями и нервной системой в виде цепочки.
Насекомые характеризуются разделением тела на три отдела - голову, грудь и брюшко, наличием одной пары усиков, трех пар ног и в большинстве случаев одной-двух пар крыльев. На земном шаре насчитывается более 1 млн. видов насекомых - больше, чем видов; всех остальных животных, взятых вместе. На территории СССР встречается около 80 тыс. видов, из которых более половины связаны в своей жизни с лесом.
Внешнее строение
Тело насекомых состоит из головы, груди и брюшка (рис. 1).
Голова
Голова образована из шести плотно слитых передних сегментов тела. Она одета общей хитиновой капсулой и, как правило, подвижно сочленена с грудью. Голова может иметь различную форму. По ее обеим сторонам между глазами прикреплены усики, или антенны. Они служат для распознавания запахов, иногда для осязания и даже захвата пищи. Антенны состоят из отдельных члеников и имеют разнообразную величину и форму. Типы антенн и число члеников на них - важный признак при определении насекомых. На голове кроме антенн и глаз расположен ротовой аппарат, служащий насекомым для добывания пищи.
Ротовой аппарат
У насекомых ротовой аппарат имеет различное строение в зависимости от способа питания. Основным и более примитивным типом ротового аппарата являются грызущие ротовые органы, свойственные большинству лесных насекомых.
Грызущий ротовой аппарат служит для питания твердой пищей. Он состоит из следующих частей: верхней губы, пары верхних челюстей, пары нижних челюстей и нижней губы. Верхняя губа представляет складку кожи над челюстями. Под ней по бокам ротового отверстия находятся верхние челюсти. Это твердые придатки с зубцами на внутренней стороне; правая и левая челюсти двигаются навстречу друг другу как две половины щипцов и откусывают частицы пищи. Под верхними челюстями находятся нижние челюсти. Они состоят из нескольких члеников и несут по нижнечелюстному щупику. Снизу ротовое отверстие прикрыто нижней губой. Она состоит из двух основных члеников, жевательных Пластинок и пары нижнегубных щупиков.
Видоизменением грызущего является грызуще-лижущий ротовой аппарат пчелиных. Главная его особенность заключается в сильном удлинении нижних челюстей и нижней губы, причем концевые отделы последней образуют очень длинный и узкий язычок. Верхняя губа и верхние челюсти почти неизменны. Этот тип ротового аппарата приспособлен для высасывания из цветков нектара и часто называется лакающим.
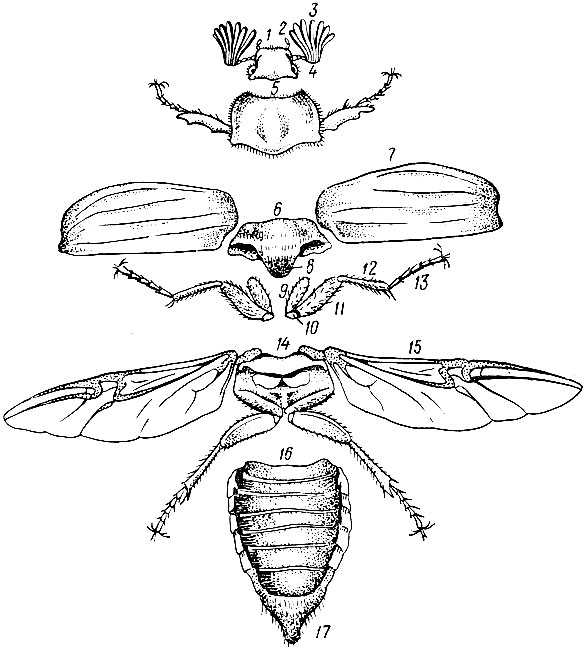
Рис. 1. Расчлененный майский жук: 1 - голова, 2 - нижнечелюстной щупик, 3 - антенны, 4 - глаза, 5 - переднеспинка, 6 - среднеспинка, 7 - надкрылья, 8 - щиток, 9 - вертлуг, 10 - тазик, 11 - бедро, 12 - голень, 13 - лапка, 14 - заднеспинка, 15 - крыло, 16 - брюшко, 17 - пигидий
Сосущий ротовой аппарат служит для питания жидкой пищей. Существует большое разнообразие ротовых органов сосущего типа. Их можно объединить в две группы: сосущие и колюще-сосущие. В первом случае пища принимается без прокола субстрата, во втором - производится прокол пищевого субстрата.
Сосущий ротовой аппарат бабочек представляет собой хоботок, закрученный в спираль. Во время питания насекомые всасывают нектар из цветков, опуская в них закручивающийся хоботок. Нектар поднимается по хоботку вследствие капиллярности и всасывающих движений клеточных мышц. Хоботок является видоизменением нижних челюстей, которые имеют вид пары желобовидных придатков, подвижно соединенных друг с другом. Верхние челюсти в процессе эволюции у бабочек исчезли полностью.
Колюще-сосущий ротовой аппарат имеется у клопов, тлей, цикад и других равнокрылых, трипсов, вшей и кровососущих двукрылых. Он приспособлен для прокалывания кожи жертвы или тканей растений и питания кровью или клеточным соком. У клопов и тлей ротовой аппарат имеет вид хоботка, в который втянута нижняя губа, состоящая из ряда члеников. В хоботке помещаются четыре колющие щетинки - видоизмененные верхние и нижние челюсти.
У кровососущих комаров ротовые органы устроены таким же образом, но нижняя губа не члениста, а верхняя губа сильно вытянута, сохранились хорошо развитые челюстные щупики.
Самое сложное видоизменение ротовых органов - лижущий хоботок многих мух. Он образован главным образом мясистой нижней губой, которая заканчивается двумя большими сосательными лопастями; челюсти обычно отсутствуют. Верхняя губа вместе с выпячиванием дна ротовой полости (языком) образует трубочку, принимающую жидкую пищу, слизанную пластинчатыми выростами нижней губы.
Грудь
Грудь насекомых состоит из трех члеников: переднегруди, среднегруди и заднегруди. В каждом членике различают четыре части, или склерита: сверху - спинку (тергит), снизу - грудь (стернит) и сбоку каждой стороны - боковые пластинки (плейриты). От члеников груди отходят придатки - ноги и крылья.
Ноги
У насекомых ноги находятся с нижней стороны груди в количестве трех пар. Каждый членик груди несет по паре ног. Нога состоит из тазика, сочленяющегося с грудью, вертлуга, бедра, голени и лапки. Лапка имеет 1 - 5 члеников, последний из них имеет два коготка.
В зависимости от условий жизни у насекомых встречаются разные типы ног: ходильные, бегательные, прыгательные, копательные и хватательные.
Крылья
Крылья представляют парные выросты или складки кожи, расположенные по бокам груди. Как правило, крыльев бывает две пары: передние крылья расположены на среднегруди и задние крылья - на заднегруди. Иногда крылья совсем отсутствуют (вши, блохи, пухоеды) или укорочены и поэтому непригодны для полета (постельный клоп, черный таракан). Имеются насекомые с одной парой крыльев, а вторая пара видоизменена в придатки (мухи).
Каждое крыло представляет складку кожи и состоит из двух тонких прилегающих друг к другу пластинок, сложенных между собой. Между ними проходят трахеи и нервы. В местах прохождения трахей хитин крыла утолщен и образует жилки, составляющие опорный скелет крыла. Жилкование крыльев разнообразно и служит одним из основных признаков при определении насекомых.
Крылья могут быть перепончатыми, кожистыми или роговыми. У насекомых, имеющих две пары крыльев, они бывают однородными или разнородными. Бабочки и перепончатокрылые насекомые имеют однородные крылья. Крылья бабочек покрыты чешуйками. Жуки, прямокрылые и полужесткокрылые имеют две пары разнородных крыльев. У жуков и прямокрылых передние крылья утолщены и превратились в надкрылья, служащие для защиты нежных задних крыльев и брюшка. У полужесткокрылых (клопы) передние крылья у основания кожистые, а к вершине перепончатые; задние крылья перепончатые.
Брюшко
В противоположность груди, которая у взрослых насекомых обычно снабжена твердым скелетом и заполнена мускулатурой, брюшко содержит главным образом внутренности и обладает мягкими покровами и слабой мускулатурой. Брюшко имеет различное число члеников - от .11 до 5, скелет каждого членика состоит из двух полуколец - тергита (верхнего) и стернита (нижнего). По форме брюшко бывает сидячее, широкостебельчатое, длинностебельчатое и висячее.
На VIII и IX сегментах брюшка находятся наружные половые придатки, или гениталии; к их числу относится у самца копулятивный орган, а у самки - яйцеклад.
Брюшко у ряда видов насекомых имеет придатки, которые являются видоизменениями брюшных конечностей; к ним относятся церки (на XI - X сегментах) и грифельки (на IX сегменте).
Внутреннее строение
Кожные покровы
Кожные покровы насекомых состоят из кожи и ее производных.
Кожа состоит из трех слоев: кутикулы, гиподермы и базальной перепонки.
Кутикула является наружным скелетом и служит опорой для прикрепления мышц. Она защищает тело насекомого от внешних воздействий, препятствует испарению воды из организма и проникновению в него ядов. Кутикула имеет тонкое гистологическое строение и состоит из наружного и внутреннего слоев.
Наружный слой, или эпикутикула, хорошо развита у наземных насекомых, непроницаема для воды и обладает гигрофобностью.
Внутренний слой, или прокутикула, во много раз толще наружного. Она содержит высокомолекулярный полимер из числа азотистых полисахаридов - хитин. Он нерастворим в щелочах, химических растворителях и слабых кислотах и придает кутикуле твердость.
Гиподерма состоит из одного слоя клеток и образует кожный эпителий, подстилающий кутикулу. Она производит кутикулу.
Базальная перепонка подстилает гиподерму. Она очень тонкая и не имеет клеточной структуры.
В целом толщина кожного покрова у насекомых различна: у одних он очень толстый (жуки), у других более тонкий и мягкий (тли).
Кожные покровы образуют ряд производных - различные скульптурные придатки (шипики, или хетоиды, бугорки, бороздки, вдавленные точки на кутикуле), структурные образования (волоски и щетинки, называемые хетами, чешуйки на крыльях бабочек), а также эндоскелет (внутренние выросты кутикулы, служащие для прикрепления мышц) и кожные железы.
Кожные железы очень разнообразны. Они могут быть одно- и двух- или многоклеточные. Известны восковые (у тлей), пахучие (у клопов), лаковые (у червецов), прядильные (у бабочек) и другие железы.
Окраска тела насекомых очень разнообразна и зависит от окраски и строения кожных покровов. Основные красящие пигменты - меланины - связаны с кутикулой и отличаются многообразием оттенков - от желтых и светло-бурых до черных.
Структурная окраска возникает вследствие особенностей строения кутикулы и расположения на ней чешуек. Она обусловлена дифракцией и интерференцией.
Мышечная система
У насекомых мышечная система состоит из соматических, или скелетных, и внутренностных, или висцелярных, мышц; они прикрепляются к скелету и внутренним органам. Соединение мышцы с соответствующей частью скелета происходит при помощи тонофибрилл, которые от конца мышечной клетки отходят к кожным покровам. Благодаря сильно развитой мускулатуре насекомые обладают большой мускульной силой. Например, у прыгающих насекомых высота прыжков превышает высоту их тела в десятки и сотни раз (блохи). Особенность мышц насекомых - способность к многократным и очень быстрым сокращениям. Летательные мышцы комаров и мух, например, сокращаются по 300, даже по 500 раз в секунду. Бабочки из семейства бражников пролетают в минуту расстояние в 22 - 25 тыс. раз больше длины своего тела.
Полость тела
У насекомого полость тела разделена двумя тонкостенными перегородками - диафрагмами - на три отдела, расположенные друг под другом. В каждом отделе помещаются определенные внутренние органы насекомых: в верхнем - кровообращения, в среднем - пищеварения, выделения, размножения и жировое тело, в нижнем находится брюшная нервная цепочка. Дыхательная система пронизывает стенки всех внутренних органов и не ограничена каким-либо одним отделом полости тела.
Жировое тело
Жировое тело представляет собой рыхлую ткань. Оно состоит из отдельных долек, или лопастей, которые заполняют всю полость тела. Клетки его наполнены каплями жира и белковыми включениями и очень близки к гемоцитам - клеткам крови насекомых. Физиологическая роль жирового тела разнообразна, но в основном сводится к накоплению питательных веществ и поглощению продуктов обмена. В течение индивидуальной жизни насекомого жировое тело претерпевает большие изменения. Накопление питательных веществ в жировом теле способствует выживанию насекомых при низких температурах, дает возможность переживать голодовку и существовать насекомым, не питающимся во взрослой фазе.
Органы пищеварения
Пищеварительный тракт состоит из трех отделов: передней, средней и задней кишок (рис. 2).
Передняя кишка начинается ротовой полостью, где пища смачивается слюной и измельчается. За ротовой полостью следует глотка, за глоткой - пищевод. Пищевод переходит в зоб, который служит для временного сохранения пищи и бывает не у всех насекомых; за зобом помещается мышечный желудок, имеющий развитую мышечную ткань. Его стенки покрыты твердыми хитиновыми зубцами, содействующими механическому перетиранию пищи. Передняя кишка заканчивается кольцевой складкой, носящей название кардиального клапана.
Передняя кишка, как и задняя, образуется из впячивания наружного зародышевого пласта (эктодермы) и в соответствии со своим происхождением выстлана хитиновой кутикулой, а средняя лишена ее.
Средняя кишка образуется путем сложного процесса из внутреннего зародышевого пласта (эндодермы). В ней выделяются все пищеварительные соки, кроме слюны, и происходит переваривание пищи и всасывание продуктов пищеварения. Средняя кишка представляет собой равномерно утолщенную трубку. Она выстлана железистым эпителием, клетки которого выделяют капли секрета, окруженные слоем плазмы. Секретирующие клетки постепенно уменьшаются и перестают существовать, а вместо них из специальных участков (крипт) возникают новые.
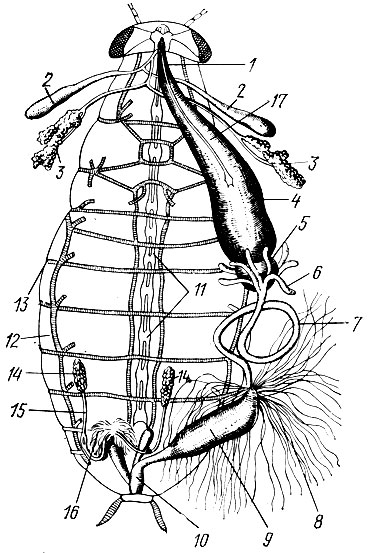
Рис. 2. Внутреннее строение черного таракана-самца (с брюшной стороны): 1 - пищевод, 2 - резервуары слюнных желез, 3 - слюнные железы, 4 - зоб, 5 - мышечный желудок, 6 - слепые отростки желудка, 7 - средняя кишка, 8 - мальпигиевы сосуды, 9 - задняя кишка, 10 - прямая кишка, 11 - узлы нервной цепочки (ганглии), 12 - трахейные стволы, 13 - дыхальца, 14- семенники, 15 - семяпровод, 16 - семяизвергательный канал, 17 - симпатическая нервная система
В средней кишке образуется также тонкая оболочка - перитрофическая мембрана, окружающая пищевой комок и предохраняющая стенку кишки от соприкосновения с плохо размельченными частицами пищи.
Задняя кишка начинается коротким пилорическим отделом, в который впадают мальпигиевы сосуды. Он регулирует с помощью кольцевой мускулатуры и специального клапана поступление пищевого комка из средней кишки в тонкую кишку, где происходит отсасывание воды из пищевой массы. Тонкая кишка переходит в прямую. Здесь происходит окончательное формирование экскрементов (кала), которые выделяются наружу через анальное отверстие. Кал имеет определенную структуру и форму, по которым можно определить, какому виду насекомого он принадлежит.
Пищеварение заключается в механической и химической переработке пищи. Характер этой переработки зависит от режимов питания насекомых. Механическая переработка пищи, ее измельчение ротовым аппаратом и перетирание в мышечном желудке имеют место только у грызущих насекомых. Сосущие насекомые употребляют жидкую пищу, всасывая ее из тканей растения в виде коллоидных растворов.
У грызущих насекомых пищеварение в свою очередь очень сильно изменяется в зависимости от состава пищи, так как химическая переработка поглощаемых листьев, древесины или животных тканей различна. В типичном случае пища в ротовой полости смачивается выделениями слюнных желез, раздробляется и попадает в среднюю кишку, где окружается перитрофической мембраной и подвергается химической переработке.
Химическая переработка пищи слагается из процессов гидролиза трех ее основных компонентов - белков, жиров и углеводов. При помощи гидролиза сложные по составу химические вещества превращаются в более простые, идущие на построение тканей и органов, а молекулы коллоидных растворов этих веществ распадаются на более мелкие, способные проникать через стенки кишечника в организм насекомого.
Гидролиз происходит с помощью ферментов, содержащихся в пищеварительных соках. Действие ферментов в значительной мере зависит от наличия оптимальной для них рН. Она может меняться в отделах кишечника и у разных насекомых. Так, у колорадского жука рН средней кишки равняется 6,6 - 7,4, у кузнечика - 5,8 - 6,9, а у ряда гусениц достигает 9,0 - 9,4. В соответствии с тремя основными группами пищевых веществ (белки, жиры и углеводы) у насекомых имеются протеазы, служащие для переваривания белков, карбогидразы, переваривающие углеводы, и липазы, расщепляющие жиры. Каждая из этих групп содержит несколько ферментов, различающихся по своему действию. Так, среди карбогидраз амилазы превращают крахмал в сахар, а целлюлоза действует на клетчатку. Белки серией ферментов расщепляются до аминокислот, а жиры под действием липаз превращаются в глицерин и жирные кислоты. Последние со щелочами образуют соли, т. е. мыла. Глицерин и мыла хорошо растворимы в воде и легко усваиваются стенками кишечника.
Набор ферментов меняется в зависимости от того, чем питается насекомое из поколения в поколение. Меняется он и на разных фазах развития. Так, листогрызущие гусеницы бабочек имеют все три группы ферментов, а взрослая бабочка из этого набора сохраняет только инвертазу, позволяющую питаться сахаром нектара.
Особенно трудно перевариваются высокомолекулярные вещества, в частности древесина. Древесина в основном состоит из клетчатки, лигнина и гемицеллюлоз. Кроме того, в ней содержатся в небольшом количестве крахмал, жиры и масла, органические кислоты, дубильные вещества, пигменты и другие химические вещества. Лигнин никогда не переваривается, а клетчатка и гемицеллюлоза - только некоторыми видами насекомых. В связи с этим различают следующие группы насекомых: личинки одних усваивают только содержимое клеток (древогрызы, некоторые усачи, ложно-короеды), личинки других усваивают также и углеводы клеточных стенок, включая гемицеллюлозы, но не усваивают клетчатки (короеды), третьи усваивают, кроме того, и клетчатку (точильщики, часть усачей). В связи с тем, что состав живой и мертвой древесины различен и быстро изменяется, насекомые, питающиеся живой древесиной, чаще всего не питаются мертвой. В питании насекомых древесиной большую роль играют простейшие организмы - симбионты. Они содержатся в кишечнике насекомых и способствуют разложению поступающей туда клетчатки (у термитов, древогрызов и др.). У короедов-древесинников самки вносят в ходы, которые делают в древесине, споры грибов. Споры прорастают и мицелий гриба подготавливает пищу для отрождающихся в ходах личинок.
В средней кишке происходит также всасывание пищи, которая попадает через стенки кишечника в кровь. Кровь, омывая внутренние органы, отдает тканям и отдельным клеткам продукты переваренной пищи, из которой создаются вещества, специфичные для тела насекомых.
Непереваренная пища поступает в заднюю кишку, где происходит всасывание воды и частично еще питательных веществ. Скорость пищеварения и всасывания пищи зависит от температуры тела. Наиболее быстро они идут при оптимальной температуре.
У ряда хищных насекомых, например жужелиц, происходит внекишечное пищеварение. Оно состоит в том, что сок средней кишки вливается через челюстные каналы в тело жертвы и переваривание начинается раньше проглатывания.
Кровеносная система
Все пространство между стенкой тела и отдельными органами представляет полость тела насекомого. Она заполнена кровяной жидкостью, кровь приводится в движение работой сердца. Сердце лежит в брюшке, на спинной его стороне, и представляет длинную трубку, состоящую из ряда камер. На заднем конце трубка обычно замкнута. С боков каждая камера имеет пару боковых отверстий с клапанами (остий), впереди сердце переходит в аорту, которая не имеет отверстий и не замкнута в голове. Сердце прикрепляется к спинной стенке тела тяжами, а от боковых сторон сердца к стенке тела отходят пучки крыловидных мышц, приводящих в движение камеры сердца (рис. 3).
Камеры сердца последовательно сокращаются одна за другой и перегоняют кровь вперед к головному концу, где она, через аорту изливается в полость головы, а оттуда в полость тела. Отсюда кровяной поток при помощи спинной и брюшной диафрагмы поступает в околосердечную полость и кровь втягивается в сердце через остии; таким образом, у насекомых при открытой кровеносной системе возникает циркуляция крови: по спинному сосуду - вперед, а в полости тела назад.
Кровь насекомых, или гемолимфа, состоит из жидкой плазмы и клеточных элементов - гемоцитов. Она представляет раствор множества неорганических и органических веществ (органические соли, питательные вещества, мочевая кислота, ферменты, гормоны, пигменты), бесцветна или окрашена. Содержание воды в крови колеблется от 75 до 90%.
Гемоциты имеют микроскопические размеры, форму пузырьков или звездочек с неясно различимыми контурами и структурой. Это амебовидные клетки, свободно плавающие в плазме (рис. 4). Они различны по форме, величине и функциям. Имеются молодые делящиеся клетки - пролейкоциты; клетки, способные заглатывать твердые тела и бактерии,- фагоциты; клетки, извлекающие из крови посторонние вещества и мочевую кислоту,- нефроциты. Встречаются еще макронуклеоциты, микронуклеоциты, эозинофилы и эпоцитоиды. Содержание их в крови различно. Оно зависит от вида насекомого, меняется по фазам развития. Состав гемоцитов меняется при разном состоянии насекомых. Поэтому часто прибегают к анализу клеточных включений крови, по изменению которых судят об их жизнеспособности.
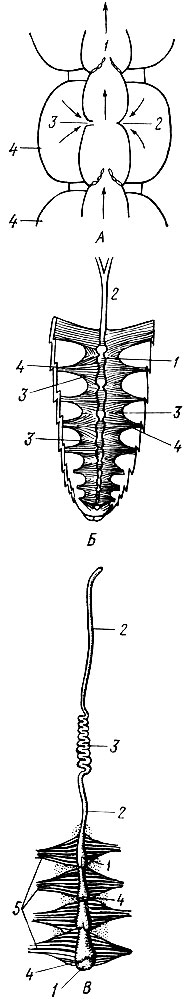
Рис. 3. Схема органов кровообращения; А - средняя часть спинного сосуда (сердце): 1 - межкамерные клапаны, 2 - закрытые, 3 - открытое устье, 4 - диафрагма; стрелки показывают направление крови;
Функции крови у насекомого разнообразны. Кровь разносит питательные вещества к тканям и переносит вредные продукты обмена к органам выделения. Кровь является носителем гормонов, содержит ядовитые и защитные вещества. Частично кровь служит переносчиком кислорода и участвует в удалении углекислоты из тканей.
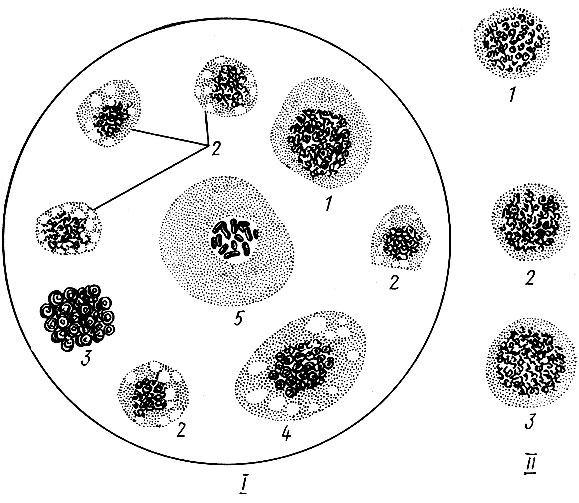
Рис. 4. Гемолимфа здоровых насекомых (по Сиротиной, 1965): / - типы гемоцитов: 1 - макронуклеоцит, 2 - микронуклеоцит, 3 - эозинофилл, 4 - фагоцит пассивный, 5 - эноцитоид; II- пролейкоциты: 1 - кольчатого шелкопряда, 2 - соснового шелкопряда, 3 - соснового пилильщика
Кровь выполняет также механическую функцию. Она создает необходимое внутреннее давление, благодаря ему у насекомых с мягкими покровами поддерживается форма тела. Кроме того, путем сокращения мышц иногда возникает повышенное давление крови, под действием которого происходит расправление крыльев у только что отродившихся взрослых насекомых, развертывание хоботка и т. д.
Органы дыхания
Органы дыхания насекомых представляют систему трахей, пронизывающих тело насекомого. Они обычно состоят из двух продольных стволов и ветвей, отходящих к различным органам и мышцам. Трахея - это трубка округлого сечения, многократно ветвящаяся и заканчивающаяся тончайшими трахейными капиллярами - трахеолами. Наружу трахейная система открывается особыми отверстиями - дыхальцами (стигмами), расположенными по бокам брюшка (до восьми пар) и на груди (две пары). У дыхалец есть замыкательные клапаны (рис.. 5), регулирующие поступление воздуха в трахейную систему.
Воздух поступает через дыхальца в трахеи у одних насекомых благодаря ослаблению мышц брюшка и увеличению объема тела, у других - путем диффузии. Дальнейшее распространение воздуха по мелким трахеям и трахеолам, а оттуда в клетки тканей происходит только с помощью диффузии.
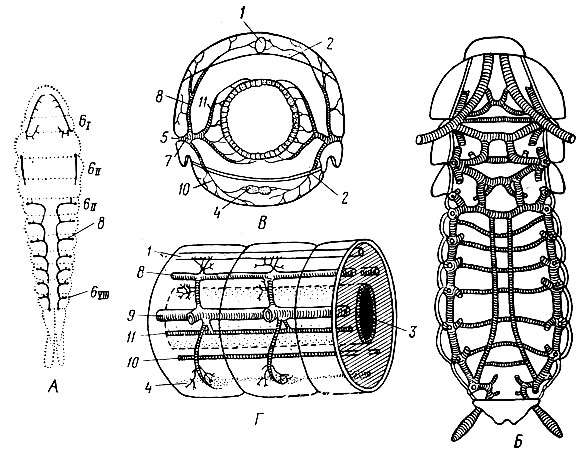
Рис. 5. Трахейная система. А - простейший тип; Б - специализированный (вентральные стволы); В - схема поперечного ветвления трахей в сегменте; Г - схема продольных стволов трахей: 1 - сердце, 2 - диафрагма, 3 - кишечник, 4 - нервная система, 5 - дыхальца, 6subI/sub - грудные сегменты, sub5II-YIII /sub- брюшные сегменты, 7 - стигмальная трахея, 8-11 - дорзальный, митральный, вентральный, висцеральный продольные стволы
При дыхании кислород поглощается из воздуха и служит для окисления молекул белков, жиров и углеводов. Процесс окисления идет при участии окислительных ферментов - оксидаз и сопровождается образованием углекислого газа и выделением тепловой и механической энергии, необходимой для жизнедеятельности организма. Удаление углекислого газа происходит через кожу и частично через трахеи.
Интенсивность дыхания или газообмена определяется количеством поглощенного кислорода и выделенного углекислого газа. Отношение между объемом выделенного углекислого газа и поглощенного кислорода называется дыхательным коэффициентом. При окислении углеводов дыхательный коэффициент равен единице. Если газообмен идет за счет жиров и белков, которые окисляются труднее, дыхательный коэффициент снижается до 0,7-0,8. Каждый вид насекомого на определенной фазе развития характеризуется определенными значениями дыхательного коэффициента, отвечающими химическому составу его основной пищи. При изменении пищевого рациона изменяется дыхательный коэффициент насекомого. Он изменяется также при метаморфозе насекомых, диапаузе, в течение вегатационного периода, при голодании.
Основной резервный питательный материал насекомых составляют жиры, поэтому при голодании и диапаузе дыхательный коэффициент заметно снижается.
У хорошо летающих насекомых в груди и брюшке имеются связанные с трахеями воздушные мешки. Перед полетом они наполняются воздухом и облегчают полет. Так, у майского хруща общая емкость трахейной системы составляет 630 мм3, что позволяет сравнительно крупному насекомому делать большие перелеты.
Органы выделения
С помощью органов выделения из организма насекомого удаляются ненужные, преимущественно азотсодержащие вещества, образующиеся в результате жизненных процессов в тканях и органах, т. е. вне пищеварительной системы насекомых. Основной орган выделения - мальпигиевы сосуды. Они представляют собой длинные трубочки, которые одним концом открываются около места соединения средней кишки с задней, другим - слепо оканчиваются в полости тела. Стенки мальпигиевых сосудов состоят изнутри из одного слоя эпителиальных клеток и нередко имеют собственную мускулатуру, обеспечивающую им подвижность. Число их различно: от четырех - шести до нескольких десятков.
Мальпигиевы сосуды поглощают из гемолимфы главным образом соли мочевой кислоты, которые превращаются в мочевую кислоту в виде кристаллов. Кристаллы удаляются через заднюю кишку вместе с калом.
Выделительные функции частично выполняют также нефроциты и жировое тело. Нефроциты поглощают из полости тела коллоидальные вещества, а жировое тело производит внутриклеточное накопление вредных веществ (экскретов). Они остаются в жировом теле пожизненно или передаются мальпигиевым сосудам и выводятся из организма.
Железы и секреция
Выше уже упоминались железы, связанные с кожными покровами. Они выделяют вещества, называемые секретами, которые поступают в различные органы, полости или наружу. Такие железы называются экзокринными. Они разнообразны по функциям и строению. Одни из них участвуют в пищеварении насекомых, другие осуществляют механическую защиту, третьи образуют биологически активные вещества, называемые феромонами. Среди феромонов наиболее изучены половые аттрактанты, позволяющие самкам привлекать самцов с больших расстояний.
Эндокринные железы выделяют секреты непосредственно в кровь, их выделения называются гормонами или инкретами. Попадая в кровь, гормоны передаются ею во все части тела. Эндокринные железы регулируют многие важнейшие процессы и развитие насекомых, как-то: личиночный рост, линьки, торможение развития, половое созревание и др. Эти железы находятся над передней кишкой позади головного мозга, в брюшной части переднегруди. Эндокринные железы действуют взаимосвязанно как единая эндокринная система, находящаяся в теснейшей связи с нервной системой, и играют большую роль в жизни насекомых. Многие стороны деятельности эндокринной системы остаются еще недостаточно исследованными.
Нервная система
Нервная система насекомых регулирует все функции организма и объединяет его в единое целое. Ее основу составляют нервные клетки - нейроны, которые в соответствии со" своим назначением делятся на чувствительные, двигательные и ассоциативные.
Нервная система насекомых сильно дифференцирована и обычно разделяется на центральную, периферическую и симпатическую.
Центральная нервная система состоит из совокупности узлов? (ганглиев), от которых отходят нервы. Нервные узлы соединяются продольными и поперечными перемычками. Вся система ганглиев разделена на два отдела - головной и брюшной. В голове над пищеводом расположен надглоточный, а под пищеводом - подглоточный ганглий. Они соединены между собой тяжами, которые образуют окологлоточное кольцо. Брюшной отдел состоит из серии ганглиев и связывающих их нервных тяжей, образующих брюшную нервную цепочку.
Надглоточный ганглий развит очень сильно и нередко называется также головным мозгом. Особенно сложно строение головного мозга у насекомых с высоко развитой нервной деятельностью-(муравьи, пчелы). Он служит главным центром, подчиняющим себе деятельность прочих нервных узлов. От него отходят нервы к глазам и усикам, а от подглоточного узла - к ротовым органам и в передний отдел кишечника.
Симпатическая нервная система регулирует работу внутренних органов' и мышечной системы насекомых и имеет весьма сложное строение. Рото-желудочный отдел этой системы связан с эндокринной системой.
Периферическая нервная система образована из нервов, отходящих от ганглиев центральной и симпатической нервных систем. Они связывают эти системы с различными органами.
Физиологические функции нервной системы основаны на общем принципе рефлекса, которым организм отвечает на внешние сигналы. Нервное раздражение с периферии тела по чувствительному нерву достигает нервного узла, а отсюда по двигательному нерву возвращается к той или иной мышце, вызывая соответствующее движение. Образуется рефлекторная дуга.
Любая рефлекторная дуга начинается с рецептора, который трансформирует энергию раздражающего стимула в нервные импульсы. Каждый рецептор обнаруживает очень высокую чувствительность к определенному кругу адекватных раздражителей и относительно низкую чувствительность к остальным, неадекватным раздражениям. Насекомые имеют множество всевозможных рецепторов. Так, число чувствительных нейронов, связанных с рецепторами антенн медоносной пчелы, приближается к 500 тыс. Совокупность рецепторов, приспособленных к восприятию одинаковых раздражителей, называется органом чувств.
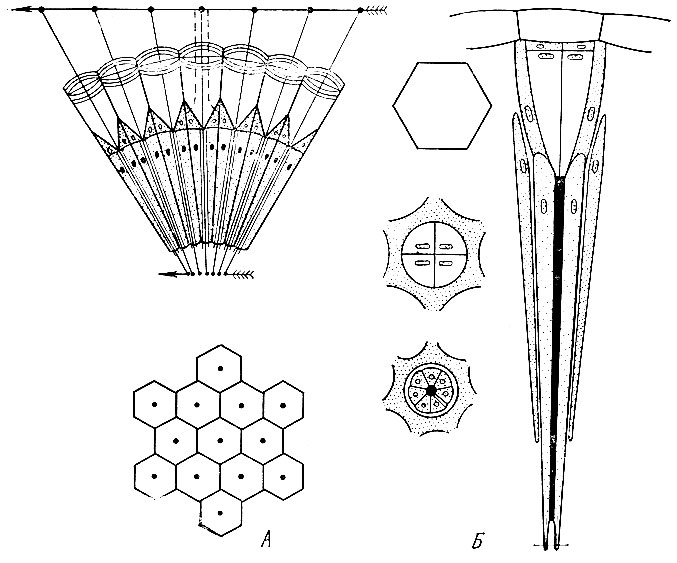
Рис. 6. Схема строения омматидия аппозиционного глаза на продольном разрезе (А) и фасетки на поверхности сложного глаза (Б)
Многочисленные сенсиллы (простейший рецептор, состоящий из воспринимающей структуры кожи и прилегающих к ней нервных чувствительных клеток), разбросанные по телу насекомого, реагируют на прикосновение к ним. Каждая такая сенсилла является одиночным осязательным рецептором, но все вместе они образуют орган осязания. Рецепторы антенн, реагирующие на запаховые раздражители, формируют орган обоняния.
Органы чувств
Выделяют следующие чувства насекомых: зрение, обоняние, осязание, слух, вкус, температурная и осмотическая чувствительность, гравитационное и двигательное чувство. Для большинства насекомых зрение и обоняние являются важнейшими источниками информации о событиях внешнего мира. На уровне органов чувств достигается только первичная обработка информации, воспринимаемой рецепторами. Высшие этапы анализа раздражений осуществляются в ганглиях центральной нервной системы, с которыми связаны органы чувств.
Обычно все рецепторы делятся на четыре группы: механорецепторы (приспособлены к восприятию механической энергии раздражающего стимула), терморецепторы (воспринимают температурные раздражения), хеморецепторы (раздражаются химическими веществами) и фоторецепторы (воспринимают световую энергию).
Особенно хорошо у насекомых развиты органы зрения.
Органы зрения - сложные, или фасеточные, глаза и простые глазки. Фасеточные глаза состоят из множества круглых или шестигранных фасеток и дают прямое мозаичное изображение.
В продольном разрезе фасетка состоит снаружи из прозрачной двояко- или плосковыпуклой хитиновой линзы - роговицы, под которой располагается коническая светопреломляющая часть - хрустальный конус. Под конусом находятся клетки сетчатки, воспринимающие световые лучи. К клеткам сетчатки подходят нервные окончания, идущие от надглоточного узла и служащие для передачи в узел световых раздражений (рис. 6).
У разных насекомых степень развития сложного глаза и зрительных восприятий различна. Одни насекомые различают только общие очертания предмета, другие обладают способностью очень четких восприятий. Так, бражники узнают знакомые им цветки на рисунках и подлетают к ним сосать нектар.
У насекомых хорошо развито цветовое зрение, однако большинство из них слепо к красному цвету, но воспринимает ультрафиолетовые лучи. Медоносная пчела различает поляризованный свет, излучаемый голубым небом, что позволяет ей ориентироваться при полете. Ряд насекомых ориентируется в зависимости от направления солнечных лучей. Прилет ночных насекомых на свет связан со светокомпасным движением. Это движение все время идет по логарифмической спирали и приводит насекомое к источнику света.
Простые глаза, или глазки, располагаются у взрослых насекомых между сложными глазами на лбу и темени. У личинок они помещены на боковых частях головы и дают возможность ориентации: ими воспринимается не форма предмета, а только его движение и свет.
Обоняние у насекомых развито очень сильно. По своей тонкости и точности оно превосходит острое чутье любого млекопитающего. Органы обоняния расположены на усиках в виде особых клеток, связанных с нервными окончаниями. Так, наездник эфиальт, ощупывая усиками кору, находит личинку рогохвоста в древесине, жуки-короеды отыскивают в лесу по запаху ослабленные деревья, самцы иногда за несколько километров находят самку и т. д.
Органы вкуса расположены на ротовом аппарате в виде чувствительных волосков. Физиологическое различие между обонятельными и вкусовыми рецепторами заключается в том, что первые анализируют газообразную среду с низкой концентрацией вещества, а вторые - жидкую среду с высокой его концентрацией.
Вкус более специфичен, так как служит только для распознавания пищи. Насекомые различают сладкий, горький, кислый и соленый вкус.
Обоняние и вкус часто объединяют, называя химическим чувством (Бей-Биенко, 1980). Оно имеет очень большое значение в жизни насекомых. На знании химического чувства построены многие меры борьбы с насекомыми.
Органы осязания представляют собой волоски, щетинки, шипики, находящиеся в усиках, щупиках и на поверхности тела. Ими воспринимаются сотрясения, Положение тела, его равновесие, соприкосновение с твердыми предметами, водой и т. п.
Органы слуха имеются преимущественно у насекомых, издающих звуки. Это чувствительные клетки, расположенные внутри отверстия, затянутого перепонкой. К ним подходят нервные окончания. Расположены органы слуха в разных местах тела. Так, у саранчовых они расположены по бокам первого сегмента брюшка, у кузнечиков - на передних голенях, а у певчих цикад - у основания брюшка.
Насекомые воспринимают широкий диапазон частот - от инфразвука до ультразвука.
Нервная деятельность
Нервная деятельность насекомых очень разнообразна и сложна. В совокупности она составляет их поведение. Поведение представляет комплексную реакцию насекомого, обусловленную воздействиями внешней среды и физиологическими особенностями организма.
Наиболее просто нервная деятельность проявляется в таксисах. Это простейшая положительная или отрицательная двигательная реакция (рефлекс) на то или иное внешнее раздражение, однако усложненная общим возбуждением центральной нервной системы. Таксисы разнообразны. Особенно развиты у насекомых термотаксис, фототаксис, гидротаксис, хемо- и трофотаксис. Соответственно в этих случаях насекомые реагируют на тепло, свет, влагу, химические раздражители и пищу. Таксисы носят приспособительный характер и потому в зависимости от условий и состояния организма могут быть положительными и отрицательными. В одном случае насекомое может стремиться в зону более высоких температур или яркого света, а в другом - в зону более низких температур или темноты.
Знание таксисов широко используется для борьбы с насекомыми с помощью приманок, ловушек и изменения среды их обитания.
Более сложным видом нервной деятельности являются инстинкты. Они представляют сложную цепь рефлексов, причем конец одного из звеньев цепи служит толчком для начала последующего звена. Проявление определенного инстинкта связано с состоянием организма насекомого. Так, созревание оплодотворенных яиц вызывает у самки появление заботы о потомстве.
Инстинкты врожденные. Никто не учил пчелу строить ячейку, короеда - прокладывать под корой определенной формы ход, жука-слоника - скручивать лист в виде трубки или сигары, гусеницу - плести кокон и т. п. Под влиянием условий жизни могут возникать новые инстинкты и могут ослабевать и даже исчезать старые, однако для этого нужен определенный, часто довольно значительный период времени.
Инстинкты могут быть направлены на сохранение вида (забота о потомстве) и особи. Особенно сложны инстинкты у пчел, муравьев и термитов, "умеющих" строить очень сложные сооружения и ухаживать за потомством, регулируя его состав.
У насекомых может в отдельных случаях наблюдаться и высшая нервная деятельность, проявляющаяся в выработке условных рефлексов. Одним из примеров служит быстрое установление у пчелы связи запаха цветка с его медоносностью.
Половая система и размножение
Насекомые раздельнополы. У многих насекомых развит половой диморфизм и самцы отличаются от самок по различным внешним признакам (по величине тела, усикам, скульптурным образованиям, окраске и др.). Однако очень часто оба пола внешне неотличимы и распознать их можно только по гениталиям.
Половая система
Половой аппарат насекомых расположен в брюшке и открывается наружу половым отверстием. Женские половые органы состоят из двух яичников, двух яйцеводов, непарного влагалища, придаточных половых желез, семяприемника и нередко яйцеклада. Половые органы самцов состоят из пары семенников, пары семяпроводов, семяизвергательного канала, совокупительного органа и придаточных желез (рис. 7).
Семенники самца представляют парную железу разнообразного строения. Семенники образуют мужские половые клетки - сперматозоиды, оплодотворяющие яйца самки.
Наиболее существенной частью полового аппарата самки являются яичники. Каждый яичник состоит из яйцевых трубок, которых бывает от 2 до 2500.
Внутри яйцевые трубочки выложены фолликулярным эпителием, из которого образуются яйцевые клетки. Из них в результате усиленного поглощения питательных веществ и сложных процессов развития и роста образуются зрелые яйца. Каждая яйцевая трубка делится на ряд яйцевых камер, в которых развиваются яйца. Созревшие яйца поступают из яичников в яйцеводы и далее через влагалище и половое отверстие выводятся наружу.
Количество откладываемых самкой яиц зависит от строения и величины яичников и специфично для каждого вида насекомого. Так, самка непарного шелкопряда может отложить 600 - 800, реже 1000 яиц, сосновая пяденица - 120 - 180, а самки многих хищных жуков - всего 10 - 20 шт. В отдельных случаях плодовитость достигает огромных размеров. Например, самка (матка) медоносной пчелы откладывает в день до 3 тыс. яиц, а у термитов - даже до 30 тыс.
Плодовитость самок резко колеблется в зависимости от погодных условий и питания (см. гл. III).
Размножение
Способность к размножению появляется у одних насекомых вскоре после окрыления, у других - через некоторый период, в течение которого они проходят дополнительное питание. Оно необходимо для созревания половых продуктов.
Существует два способа полового размножения - обоеполый, когда при спаривании яйца самки оплодотворяются сперматозоидами самца, и девственный, когда яйцо развивается без оплодотворения (партеногенез). Девственное размножение произошло из обоеполого как его упрощение и наблюдается у тлей и других насекомых. Оно представляет очень выгодное приспособление, дающее возможность насекомым быстро увеличить численность и расселяться по территории.
Партеногенез разнообразен в своих проявлениях. Так, у медоносной пчелы из неоплодотворенных яиц развиваются самцы (трутни), а из оплодотворенных - самки (рабочие пчелы, матки). У ряда насекомых наблюдается правильное чередование обоеполых и многочисленных партеногенетических поколений. Так, у многих орехотворок чередуются два поколения: летнее - крылатое обоеполое и осеннее - бескрылое, состоящее только из самок. Еще сложнее чередование поколений (гетерогония) проявляется у тлей. Оно описано в гл. VII.
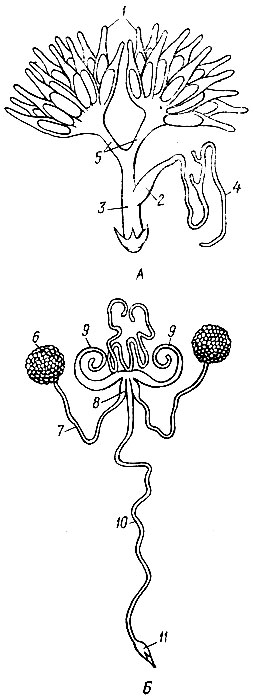
Рис. 7. Половые органы насекомых. А - самки; Б - самца чернотелки: 1 - яичники, 2 - семяприемник, 3 - влагалище, 4 - железа семяприемника, 5 - яйцепроводы, 6 - семенник, 7 - семяпроводы, 8 - семенной пузырек, 9 - придаточные железы, 10 - семяизвергательный канал, 11 - совокупительный орган
Одна из форм партеногенеза - педогенез, или детское размножение. В этом случае в яичниках личинки происходит партеногенетическое развитие яиц, на которых возникают личинки. Такое размножение повторяется в течение нескольких поколений, а затем сменяется серией обоеполых поколений со взрослой фазой. Оно наблюдается у галлиц и отдельных видов жуков и клопов.
У наездников бывает дробление яйца, отложенного в другое насекомое. В результате в теле "хозяина" развивается не один, а множество (до 100) особей наездников. Такой способ размножения называется полиэмбрионией.
Иногда у кровососущих мух и других насекомых наблюдается живорождение. При этом вместо откладки яиц на свет производятся личинки.
Развитие
Развитие насекомых делится на два периода: зародышевое, или эмбриональное (внутри яйца), и внеяйцевое, или постэмбриональное.
Эмбриональное развитие
Развитие зародыша в яйце начинается дроблением яйца с момента оплодотворения или откладки яиц. Яйцо имеет круглую, овальную, удлиненную или другую форму и представляет собой одну крупную клетку, включающую питательный желток, необходимый для роста и развития зародыша. Снаружи яйцо покрыто оболочкой (хорион), играющей роль скорлупы. На одном из полюсов яйца имеется одно или несколько мельчайших отверстий (микропиле), через которые проникают сперматозоиды при оплодотворении. Яйцо после откладки часто всасывает влагу из окружающей среды и вследствие этого увеличивается в два-три раза.
Зрелые яйца самки откладывают поодиночке или группами на листья, ветви и стволы деревьев, на почву, травянистую растительность и другие предметы. Часто яйца бывают погружены в субстрат (в древесную ткань или почву) или защищены пушком, снятым с конца брюшка самки (у златогузки, непарного шелкопряда и других бабочек), или щитком, образовавшимся из выделений придаточных желез (у зеленой узкотелой златки).
Развитие зародыша в яйце продолжается от нескольких дней до месяца и более. Оно заключается в ряде сложных преобразований и заканчивается вылуплением личинки, прогрызающей оболочку яйца (рис. 8).
Постэмбриональное развитие
Начинается после вылупления из яйца личинки и продолжается от нескольких дней до нескольких лет. Рост личинки сопровождается линьками, которые стимулируются деятельностью желез внутренней секреции. При этом в кровь поступает гормон, вызывающий деление клеток гиподермы и выделение ими новой кутикулы. В то же время кожные личиночные железы начинают изливать под старую кутикулу секрет, который растворяет внутреннюю кутикулу снизу, отчего она лопается по средней линии спины и сбрасывается. Линьками разделены возрасты личинок. Число возрастов различно. У многих личинок жуков всего три, у гусениц бабочек чаще всего пять, а у поденок до 45. Количество возрастов может меняться в зависимости от температуры и пищи, при голодании.
В процессе постэмбрионального развития насекомые не только растут, но и претерпевают ряд изменений. Совокупность этих изменений называется превращением или метаморфозом. Основной момент метаморфоза состоит в приобретении крыльев. В зависимости от этого различают два основных типа метаморфоза: неполный (насекомые с неполным превращением - прямокрылые, клопы, цикады, тли, стрекозы и др.) и полный (насекомые с полным превращением - жуки, бабочки, перепончатокрылые, мухи).
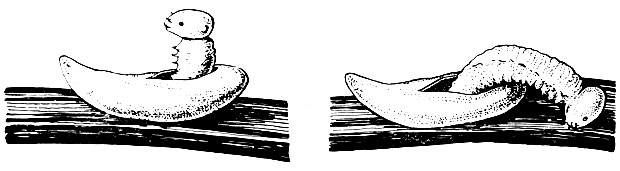
Рис. 8. Вылупление личинки из яйца звездчатого ткача
Насекомые с неполным превращением характеризуются наличием трех фаз развития: яйца, личинки и взрослого насекомого (имаго). Выходящая из яйца личинка у них близка по внешнему виду и биологически к взрослому насекомому: туловище расчленено на грудь и брюшко, которые несут соответствующие придатки. Крылья появляются в виде наружной складки кожи на краю грудного сегмента. Она увеличивается при каждой линьке и постепенно достигает окончательной величины и формы. Пищевой режим у личинки такой же, как у взрослого насекомого (рис. 9, А).
Насекомые с полным превращением имеют четыре фазы развития: яйцо, личинку, куколку и взрослое насекомое (имаго). Выходящая из яйца личинка червеобразной формы. Ее туловище состоит из серии одинаковых сегментов. Брюшко и грудь личинки не обособлены; имеются только сегменты, из которых они потом образуются. Таким образом, личинка резко отличается от взрослого насекомого своим внешним видом.
Личинка питается чаще всего совершенно по-другому и ведет образ жизни иной, нежели взрослое насекомое. После последней линьки личинка превращается в куколку, по своему внешнему виду похожую на взрослое насекомое, иногда покрытую оболочкой (рис. 9, Б). Перед превращением в куколку личинка часто покрывает себя коконом, материал для которого готовят шелкоотделительные железы (бабочки), а иногда выделяют мальпигиевы сосуды (некоторые жуки, сетчатокрылые).
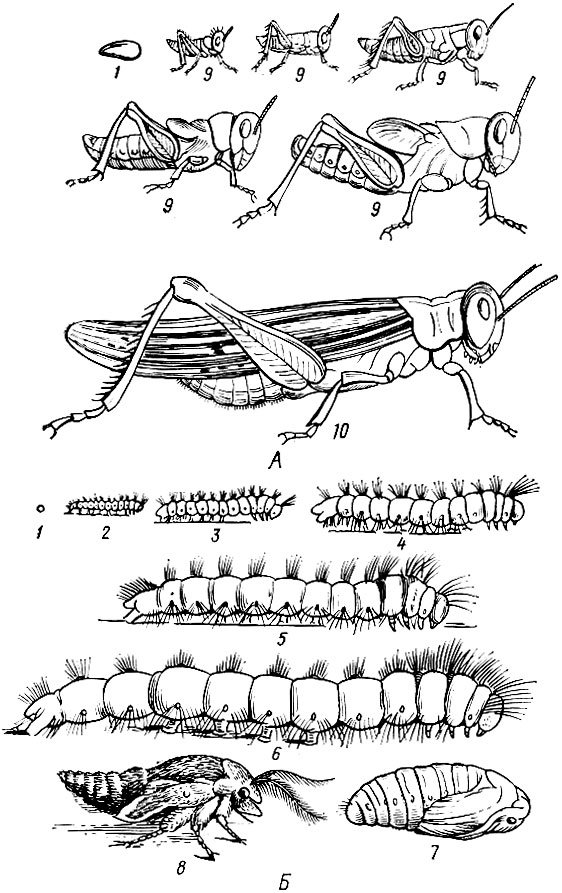
Рис. 9. Метаморфоз у насекомых. А - неполное превращение (разные возрасты саранчи); Б - полное превращение соснового шелкопряда: 1 - яйцо, 2-6 - гусеницы в различных возрастах, 7 - куколка, 8 - только что вылупившаяся бабочка, 9 - личинки, 10 - взрослое насекомое
При полном превращении происходит сложная перестройка организма, начинающаяся у личинки перед окукливанием и заканчивающаяся у куколки. На протяжении этой перестройки временные органы личинки заменяются органами, характерными для взрослого насекомого. Зачатки органов взрослого насекомого скрыты у личинки в разных частях тела и называются имагинальными дисками. Из них развиваются крылья, ноги и ротовые органы взрослого насекомого, а также трахеи, мускулатура, пищеварительная система и др. Перестройка слагается из двух процессов: гистолиза и гистогенеза.
Гистолиз состоит в разрушении тех органов, которые функционируют у личинки и отсутствуют у взрослого насекомого. Разрушение происходит благодаря оживившейся деятельности кровяных клеток, которые активно внедряются в органы и поглощают куски ткани. Существует мнение, что этот процесс происходит химически, а кровяные клетки только подхватывают продукты распада.
Гистогенез представляет образование различных тканей из недифференцированного клеточного материала - продуктов гистолиза. Активным элементом в гистогенезе служат имагинальные диски. Количество имагинальных дисков зависит от интенсивности метаморфоза и особенно велико у мух.
Весь процесс метаморфоза тесно связан с работой желез внутренней секреции. Гормон экдизон регулирует нормальное развитие личинки, способствует развитию у нее гонад, дифференциации тканей и линьке. Ювенильный гормон препятствует линьке во взрослую фазу и стимулирует рост и развитие личиночных органов.
У насекомых встречается ряд видоизменений неполного и полного метаморфоза. Так, при неполном превращении иногда наблюдается почти полное отсутствие изменений у личинки в процессе ее жизни или, наоборот, приближение к полному метаморфозу (гиперморфоз). В этом случае у взрослой личинки приобретается неподвижность, и она получает название нимфы, напоминая куколку при полном превращении. Однако она не утрачивает прежнего сходства с личинкой.
При полном превращении наблюдается иногда несколько форм личинок, последовательно сменяющих друг друга в процессе роста и развития. Это связано с резкими изменениями их образа жизни в течение развития. Примером могут служить личинки нарывников. Сначала личинка очень неподвижна. Она проникает в кубышки саранчовых и гнезда, где питается яйцами диких пчел, а затем покидает их и проходит ряд последующих фаз. Такое превращение называется гиперметаморфозом.
Жизненный цикл
Период развития насекомого от яйца до половозрелого состояния называется жизненным циклом или генерацией (поколением). Разные виды насекомых развиваются с различной быстротой. У одних в течение года бывает одно поколение, у других два или больше. Так, у тлей в году бывает до 15 поколений, непарный шелкопряд всегда развивается в одном поколении, древесница въедливая имеет двухлетний, большой дубовый усач - трехлетний, а майский хрущ - четырехлетний цикл развития.
Жизненный цикл насекомых сложился исторически в процессе их эволюционного развития в условиях определенной географической среды. Он может быть одинаковым в пределах всего распространения вида и может изменяться в зависимости от географических условий. Так, майский хрущ в лесостепной зоне имеет четырехлетнюю генерацию, а на севере лесной зоны развивается пять лет. Обыкновенный сосновый пилильщик в лесной зоне развивается за год, а в степной дает два поколения в год. Генерация берестового листоеда под Москвой одногодовая, а в Средней Азии тройная (три поколения в год).
Ряд видов в зависимости от погодных условий может иметь в одном и том же географическом районе одну или две генерации в год. Так, у короеда-типографа под Москвой в теплое лето бывает два поколения, а в холодное и дождливое - одно. Сибирский шелкопряд также может развиваться в зависимости от температурных условий по одно- или двухлетнему циклу.
Продолжительность развития насекомых в любой их фазе в известной мере зависит от климата и погодных условий местности, в которой они обитают. Поэтому и сроки появления отдельных фаз развития насекомых будут различны в разных местах: они меняются по годам и даже в пределах одного и того же места. Лёт непарного шелкопряда под Москвой в середине июля, а в Молдавии - в конце июня. В годы с ранней дружной весной майский жук появляется в средней полосе во второй половине апреля, а в годы с поздней весной - в мае.
Установление сроков появления отдельных видов фаз развития насекомых имеет огромное значение для борьбы с вредителями. Многолетние наблюдения за насекомыми позволяют установить средние многолетние даты их появления и связать их с наступлением ряда других природных явлений. Такие наблюдения получили название фенологических, а наука, изучающая сезонные, повторяющиеся ежегодно явления природы и зависимость их от окружающей среды, называется фенологией.
Для того чтобы не упустить сроки появления насекомых, можно пользоваться совпадением отдельных фаз их развития с хорошо заметными фенологическими явлениями, которые используются как феносигналы. Сигналами служат метеорологические данные (вскрытие рек, таяние снега, суммы среднесуточных температур выше нуля и др.) и появление отдельных фаз развития обычных лесных растений. Так, лёт майского жука совпадает с распусканием листьев березы, а кладка яиц короедом-типографом - с цветением одуванчика.
Жизненный цикл насекомых (генерация) обычно изображается графически в виде схемы с условным обозначением отдельных фаз развития. Пример такого изображения, получившего название календаря развития, или фенограммы, приводится на рис. 10.
Для определения районов одновременного появления отдельных фенофаз насекомых составляются фенологические карты, основанные на фенологических наблюдениях в ряде пунктов. На карту наносятся изофены - линии одновременного наступления какого-либо фенологического явления в данном районе (начало лета насекомых, появление личинок, окукливание и т. д.). Такие карты могут сыграть большую роль для своевременного проведения мер борьбы с вредителями.
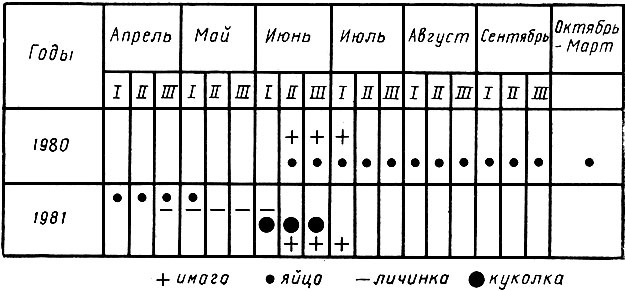
Рис. 10. Календарь развития дубовой зеленой листовертки
Диапауза
Возникновение в истории Земли ритмично повторяющихся периодов, когда потребности организма насекомых в тепле и пище не могли быть удовлетворены, явилось причиной формирования различных приспособлений к этим условиям среды. Такими приспособлениями у насекомых служат замедления и перерывы в развитии, вызываемые торможением обмена и переходом насекомого из состояния активности в состояние покоя разной глубины и продолжительности. Самым глубоким состоянием физиологического торможения обмена веществ является диапауза. Ее не следует смешивать с оцепенением, которое может наступать в любой момент под влиянием неблагоприятных (сублетальных) условий, бывает чаще всего кратковременным и сразу же проходит при наступлении нормальных условий. В отличие от оцепенения диапауза представляет собой специальное приспособление в жизненном цикле насекомых. В умеренном климате диапауза обеспечивает синхронизацию жизненного цикла насекомых с сезонной сменой внешних условий. Она может наступать на всех фазах развития насекомых. В соответствии с этим различают диапаузу эмбриональную (на фазе яйца), личиночную (на фазе личинки), куколочную (на фазе куколки) и имагинальную (на фазе имаго). В разных систематических группах лесных насекомых выражено преобладание определенной формы диапаузы. Так, листоеды имеют чаще всего имагинальную диапаузу, совки и многие пяденицы - куколочную, листовертки - эмбриональную. Иногда один и тот же вид насекомого может иметь несколько форм диапаузы. Так, для зимней пяденицы характерны эмбриональная и куколочная формы диапаузы. Чаще всего диапауза зависит от экологии вида. Даже у очень близких видов одного рода, но живущих в разных условиях, диапауза бывает на разных фазах развития, а у видов неродственных семейств - на одной и той же фазе, если они имеют сходный образ жизни.
Основной чертой диапаузы служит особая физиологическая подготовленность организма, позволяющая длительно выдерживать условия, летальные для недиапаузирующих фаз насекомых. Диапауза характеризуется приостановкой роста и развития, падением интенсивности дыхания и потребления кислорода, глубокими изменениями в системе окислительных ферментов, уменьшением общего количества воды в теле, наличием в тканях обильных отложений резервных питательных веществ, отсутствием или снижением подвижности.
Для диапаузирующих фаз развития насекомых характерна широкая комплексная устойчивость к внешним условиям. При диапаузе насекомые могут долгое время оставаться без пищи. Низкий уровень обмена веществ обусловливает очень экономное расходование резервных питательных веществ, в связи с чем диапауза может длиться долгое время, иногда несколько лет. Так, у пяденицы Oporinia autumnata в условиях Швеции она длилась 10 лет. Жизнедеятельность организма во время диапаузы обеспечивается за счет жировых отложений - особых жиро-белковых гранул и гликогена.
В период диапаузы насекомые очень устойчивы к иссушению. Это обусловлено отсутствием питания, а следовательно, и поступления воды через кишечник. Устойчивость к сухости особенно характерна при диапаузе летнего типа, но иногда бывает и при зимней диапаузе, определяя исход зимовки.
Различают диапаузу обязательную, или облигатную, и необязательную, или факультативную.
Обязательная диапауза обеспечивает прохождение в течение года только одной генерации. Она свойственна насекомым с годичным циклом развития, обычным обитателям лесов умеренного пояса. Так, у непарного шелкопряда, дубовой зеленой листовертки и ряда других листогрызущих насекомых существует обязательная эмбриональная диапауза. Она создает условия для весеннего питания молодыми листьями, содержащими большое количество необходимых питательных веществ (см. гл. VIII). У златогузки молодые гусеницы впадают в диапаузу еще задолго до листопада и наступления холодов. Такая диапауза обеспечивает им возможность закончить развитие на более питательном корме весной.
Диапауза может наступать под влиянием различных причин. Однако подготовка организма к переживанию неблагоприятных условий начинается задолго до их наступления, вне прямой зависимости от них. Поэтому регуляция диапаузы в природе может осуществляться только факторами сигнального порядка, закономерно предшествующими наступлению сезонов, неблагоприятных для развития. А. С. Данилевский установил, что таким фактором является годовой ход длины дня. Он отличается устойчивостью и астрономической точностью. Работы А. С. Данилевского (1961), а затем В. П. Тыщенкова (1977) и их учеников, изучавших в течение ряда лет роль фотопериодизма как регулятора сезонной цикличности насекомых, имеют большое значение для понимания диапаузы и других сезонно-циклических приспособлений насекомых. Однако они еще не объясняют физиологический механизм диапаузы. Регуляция диапаузы связана с работой гормонов, деятельность которых направляется внешними стимулами. Ее закономерности до конца не изучены, но выяснено, например, что нейросекреторные клетки тутового шелкопряда выделяют гормон диапаузы. Он попадает в гемолимфу самки и через яичники передается яйцам, которые под его воздействием впадают в диапаузу.
При диапаузе происходят скрытые процессы физиологической перестройки, в конечном итоге приводящие к восстановлению способности к активному развитию. Этот процесс получил название реактивации.
Один из широко распространенных механизмов реактивации - воздействие пониженных температур. Охлаждение, по-видимому, стимулирует деятельность нейросекреторных клеток головного мозга, и они начинают выделять активационный гормон. Однако это не единственный путь, ведущий к реактивации. Механизм ее разнообразен и определяется теми условиями среды, в которых существуют виды насекомых.
Общественный образ жизни и защитные приспособления
Насекомым свойствен целый ряд биологических особенностей, знание которых необходимо. К их числу относятся защитные приспособления и общественный образ жизни.
Среди защитных приспособлений различают активную защиту и отпугивание, мимикрию и криптизм. Пчелы и осы используют жало для активной защиты и нападения. Жужелицы выбрасывают ядовитую жидкость из анального отверстия. Очень эффективно выбрасывание богомолом крови в виде пены. Бабочка ночной павлиний глаз сидит со сложенными крыльями, а при приближении птиц раскрывает их и "угрожает" четырьмя громадными глазчатыми пятнами. Так же поступает глазчатый бражник и ряд других бабочек. Лжегусеницы соснового пилильщика с приближением опасности одновременно изгибают тело, принимая S-образную форму. Отпугивающей окраской обладают кокцинеллиды, бабочки-пестрянки и многие другие насекомые. Их окраска, обычно состоящая из сочетания резких контрастных тонов, "пугает" врага, когда он приближается.
Другим защитным приспособлением у насекомых является мимикрия. Ряд насекомых, лишенных надежных форм защиты, имеет вид, напоминающий других, более вооруженных средствами защиты. Насекомое, которому подражает другое, называется моделью, а самое явление подражания - мимикрией. Примером может служить бабочка-стеклянница, подражающая осе, шершню или пчеле. Мимикрия очень распространена среди этого семейства, поэтому многие его представители получили названия пчеловидная, осовидная, муравьевидная стеклянницы и т. д.
Особенно широко распространено среди лесных насекомых явление покровительственной окраски и формы. Криптизм (крипто - скрытый) состоит в том, что насекомые имитируют предметы окружающей обстановки и потому становятся незаметными на фоне своего местообитания. Криптическая внешность не дает полной гарантии сохранения, но помогает особи в борьбе за жизнь. Так, гусеницы пядениц часто имитируют сухие сучки на деревьях, богомолы и кузнечики имеют окраску под цвет зеленой травы на лугах, крылья у соснового шелкопряда трудно отличить на фоне коры соснового ствола, а совка-лишайница почти незаметна на коре дуба. Иногда различные темные и светлые пятна на плоском крыле бабочки похожи на впадины и выпуклости. Тропическая бабочка - каллима, или бабочка-лист, замечательно имитирует таковой, когда сидит в состоянии покоя со сложенными крыльями. Окраска, форма и поза покоя координированы и составляют целую систему или криптому. Все описанные выше защитные приспособления были в свое время использованы Ч. Дарвином при доказательстве естественного отбора и широко известны в биологии.
Некоторые группы высших перепончатокрылых и термиты ведут так называемый общественный образ жизни. Они живут крупными семьями. Главное при таком образе жизни - развитие полиморфизма, когда существует несколько внешне различающихся форм одного и того же вида. Эти формы, как правило, приспособлены к выполнению своих особых функций в популяциях или семьях видов. Обычно в одной семье имеются самки, самцы и рабочие особи (рис. 11). В семье имеется одна или несколько половозрелых самок, называемых матками. Основную массу семьи составляют неполовозрелые самки, называемые рабочими. Самцы присутствуют в семье чаще всего только в период спаривания самок. Матка обычно откладывает огромное число яиц. Воспитание потомства поручается рабочим. Все общественные насекомые строят сложные гнезда.
Механизм полиморфизма у общественных насекомых весьма сложен. Здесь оказывают влияние обмен пищей между всеми членами семьи (трофотаксис), при котором разносятся феромоны, выделяемые самкой и тормозящие развитие половых желез у рабочих особей, а также направленное воспитание личинок и ряд других воздействий.
Одной из форм полиморфизма является образование стадной и одиночной форм у одного вида насекомого, например у саранчовых, кузнечиков и некоторых видов гусениц бабочек. Эти фазы возникают вследствие группового эффекта, т. е. реакции насекомых на увеличение численности популяций. Взаимодействие особей в популяции при их высокой численности отличается от взаимодействия в малочисленных популяциях. При большой плотности стимулируется нейросекреция, повышается их активность, ускоряется развитие, изменяется окраска, а иногда даже морфологические элементы.
Итак, в целом полиморфизм - одна из форм приспособления насекомых к внешней среде, выработанной в процессе естественного отбора.
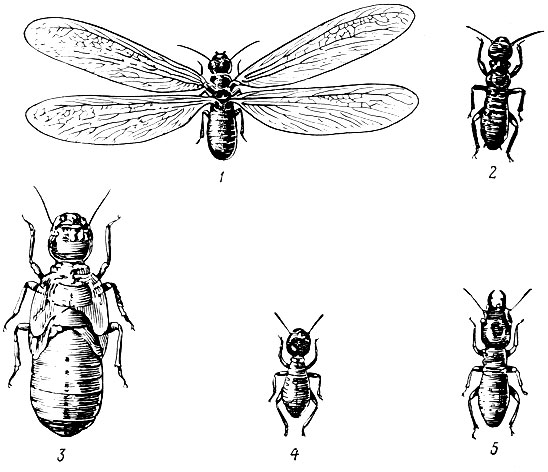
Рис .11. Полиморфизм у туркестанского термита: 1 - крылатая особь, 2 - половозрелая особь с отброшенными крыльями, 3 - нимфа, 4 - рабочий, 5 - солдат (увеличено в 1,5 раза)
Литература
Бей-Биенко Г. Я. Общая энтомология. 3-е изд. М., 1980.
Воронцов А. И., Мозолевская Е. Г. Практикум по лесной энтомологии. 2-е изд. М., 1978.
Данилевский А. С. Фотопериодизм и сезонное развитие насекомых. Л., 1961
Добровольский Б. В. Фенология насекомых. М., 1969.
Догель В. А. Зоология беспозвоночных. 7-е изд. М, 1981.
Захваткин Ю. А. Эмбриология насекомых. М., 1975.
Зенкевич Л. А. (ред.). Жизнь животных, т. 3. Беспозвоночные. М., 1969.
Мазохин-Поршняков Г. А. Зрение насекомых. М., 1965.
Малышев С. И. Перепончатокрылые, их происхождение и эволюция. М., 1959
Малышев С. И. Становление перепончатокрылых и фазы их эволюции. М.-Л., 1966.
Тыщенко В. П. Основы физиологии насекомых, ч. 1. Физиология метаболических систем. Л., 1976.
Тыщенко В. П. Основы физиологии насекомых, ч. 2. Физиология информационных систем. Л., 1977.
Тыщенко В. П. Физиология фотопериодизма насекомых.- Тр. Всесоюзн. энтомолог. об-ва, т. 59, Л., 1977.
Фабр Ж. А. Инстинкт и нравы насекомых. 2-е изд./Под ред. И. Я. Шевырева. Спб., 1912.
Фабр Ж. А. Жизнь насекомых. Л., 1963.
Шванвич Б. Я. Курс энтомологии. М., 1949.
Шванвич Б. Н. Введение в энтомологию. Л., 1959.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'