2. Таксисы и тропизмы
Общепринятой классификации форм двигательной активности и способов ориентации насекомых до сих пор нет. Отчасти повинны в этом исследователи, описавшие под одним и тем же названием разные явления, но главная причина кроется в многообразии и сложности самих изучаемых явлений.
Чтобы составить верное представление о проблеме ориентации, условимся о следующем. Таксисом будем называть только принцип ориентации, или алгоритм наведения на цель, который по идее не зависит от рода ориентирующего стимула и управляет не обязательно поступательным движением организма. Таксисов известно пока всего пять: фобо-, клино-, тропо-, тело- и менотаксис. Таксису будем противопоставлять понятие "тропизм" - целенаправленный двигательный акт, мотивированный внутренним состоянием организма. В основе тропизма лежит концентрация внимания животного на определенном сигнальном стимуле, выбор которого определяет внутренняя мотивация действий.
Обычно термины "таксис" и "тропизм" употребляют как синонимы, а многие авторы их вообще не различают. Однако в таком случае невозможно отличить принцип ориентации от биологической мотивировки выбора привлекающего стимула. Таксис позволяет движению быть направленным, а тропизм делает его целенаправ-ленным. Тропизмы указывают на признаки (сигналы), по которым животное находит пищу, укрытие или полового партнера, а таксисы объясняют способ, каким животное локализует в пространстве требуемый объект и удерживает путь к нему. Таксисы целесообразно классифицировать и именовать по алгоритму ориентации, а тропизмы- по ориентирующему фактору (фото-, хемо-, геотропизм и т. д.). Из сказанного понятно, что тропизм всегда имеет знак - положительный или отрицательный и что один и тот же тропизм может осуществляться на основе разных таксисов. Например, положительный фототропизм может осуществляться с помощью механизма тело- или тропотаксиса, в таком случае соответственно через фототело- или фототропотаксис. Тропизмы включаются, видимо, при действии "ключевых стимулов" эталогов, и к их запуску могут быть причастны узконастроенные чувствующие или вставочные нейроны-детекторы, о которых упоминалось в предыдущих главах. Сам выбор насекомым того или иного таксиса зависит от окружающей обстановки, расстояния до цели, а также от индивидуальных особенностей организма. Ниже мы будем рассматривать только таксисы и не коснемся тропизмов. Механизм последних почти совсем не расшифрован, да и относятся тропизмы скорее к области этологии, чем к физиологии органов чувств.
Фоботаксис - простейший по принципиальной схеме и исполнению способ ориентации, который следует рассматривать филогенетически самым древним. В его основе лежат кинезы, которыми принято именовать изменения локомоторной активности организма в ответ на изменение внешнего воздействия. Такие изменения активности можно назвать вынужденными: замедление или ускорение движения - ортокинез и изменение частоты смены направления движения - клинокинез. Например, у многих насекомых и их личинок наблюдается фотоортокинез, выражающийся в ускорении движения при увеличении интенсивности освещения. Столь же обычен термоортокинез - ускорение движения с ростом температуры. Клинокинетические реакции известны в отношении температуры, влажности, запаха и других факторов. Например, в зоне угрожающих температур насекомое чаще изменяет направление движения, чем в зоне оптимума. Таким образом, алгоритм фобо-таксиса предельно прост: чем дальше условия от оптимума, тем быстрее двигайся и чаще изменяй направления движения. Такой алгоритм случайного самонаведения позволяет насекомому в конце концов найти оптимальную зону и дольше удерживаться в ней, чем где-то в стороне. Фоботаксисом легко объяснить происхождение скоплений насекомых в местах с предпочитаемыми температурой, влажностью, освещенностью. Насекомые прибегают к фобо-таксису обычно в обстановке с малым градиентом стимулов, в диффузном поле, где нет четко выраженных ориентиров, либо когда неожиданно попадают в неблагоприятные условия или бывают резко возбуждены.
Все остальные таксисы отличаются от фоботаксиса тем, что их алгоритмы, во-первых, балансные, предусматривающие выравнивание последовательных или одновременных возбуждений следящих рецепторов, во-вторых, они обеспечивают направленное наведение на объект поиска.
Клинотаксис - ориентация путем сравнения и достижения постоянного уровня возбуждения одних и тех же рецепторов в процессе активного перемещения их в разные точки стимулирующего поля. Наведение локомоторной оси организма на стимул (или от него, в зависимости от знака тропизма) достигается путем последовательного маятникообразного движения антеннами или всей головой. В положении максимального (минимального) возбуждения рецепторов насекомое поворачивает тело так, чтобы оно приняло нормальное относительно этих рецепторов положение. Ориентация на основе клинотаксиса напоминает способ локализации источника звука человеком, глухим на одно ухо: чтобы точнее определить направление, откуда исходит звук, он вынужден поворачивать голову влево и вправо, сравнивая при этом ощущения громкости.
Фотоклинотаксис легко наблюдать у личинок комнатной мухи Musca domestica, избегающих свет вплоть до окукливания. Благодаря последовательным движениям передней частью тела они выбирают такое положение, в котором наименее возбуждены фоторецепторы, лежащие в складке головоглоточного скелета. Клинотаксическая ориентация по градиенту запаха описана у медоносной пчелы. Когда пчелу с одной антенной (вторая антенна ампутирована) помещали в V-образную камеру, одно колено которой содержало пахучую приманку, то в ходе поиска пчела поворачивала свою единственную антенну из стороны в сторону. Если же антенну приклеивали к голове, то у пчелы наблюдались маятни-кообразные колебания всего тела. При относительно больших расстояниях до источника запаха насекомое прибегает к клино-таксической ориентации, если даже у него функционируют обе антенны. Так, например, поступает самец тутового шелкопряда Bombyx топ на расстоянии свыше 20 см от самки. Характерный "рыскающий" полет типичен для крупных навозных и падальных жуков, когда они ищут пищу. Но здесь клинотаксис, вероятно, комбинируется с типичным фоботаксисом, так как по мере приближения к источнику запаха - зоне оптимума - скорость полета и число смен направления резко уменьшаются.
Если алгоритм клинотаксиса предусматривает баланс последовательных возбуждений одних и тех же рецепторов, то в основе остальных таксисов лежит уравнивание синхронных возбуждений разных рецепторов.
Тропотаксис - наведение локомоторной оси тела на стимул (или от него) путем достижения равновесия в возбуждении симметричных рецепторов (глаз, антенн и др.). Тропотаксическая ориентация известна по отношению к источнику света, тепловых лучей, звука, гравитации и других стимулов. Примером такой ориентации может служить поведение медоносной пчелы в упомянутой выше V-образной камере, в одном из колен которой заключена пахучая приманка. В норме пчела, расставив антенны в стороны, безошибочно находит приманку. Если антенны закрепить на лбу в перекрещенном положении, то пчела всякий раз поворачивает в непахнущее колено. Насекомое удаляется от источника запаха потому, что более сильные сенсорные импульсы вызывают соответствующие моторные команды не на той стороне тела, которой пчела должна повернуться для достижения баланса возбуждения рецепторов правой и левой антенн. Когда же антенны лежат параллельно друг другу или расставлены под некоторым углом, то насекомое ориентируется тропотаксически. Если антенны сведены так, что их вершины почти соприкасаются, то пчела ищет приманку только клинотаксически.
Ось, вокруг которой лежат антенны и другие парные рецепторные органы, хотя и совпадает с морфологической серединой, основа ее в конечном счете остается физиологической. Это доказывается, в частности, сравнением поведения интактной и ослепленной на один глаз личинки жука-плавунца Acilius, которая тропотаксически ориентирует свое тело по источнику света как в горизонтальной, так и в вертикальной плоскостях. В норме она удерживает положение спиной к свету, предупреждая не только вращательные движения по горизонтали (путем сохранения равенства возбуждения правой и левой групп стемм), но и килевую качку, что достигается за счет баланса между возбуждением передних и задних стемм. Односторонне ослепленная личинка непрерывно поворачивается.
Тропотаксис отличается от во многом сходно проявляющегося в поведении организма телотаксиса (см. ниже) следующими признаками.
- Насекомое, помещенное перед двумя одинаковыми источниками раздражения, например лампами, движется по средней линии между ними и только в непосредственной близости сворачивает к одному из них, т. е. переходит к телотаксису.
- При нарушении симметрии рецепторов, например путем закрашивания одного глаза, насекомое совершает даже в параллельном пучке света манежные (круговые) движения, оставаясь зрячей стороной к стимулу.
Телотаксис - наведение на стимул путем достижения такого положения тела, когда особая зона осевых, направленных прямо вперед чувствующих элементов, например зона фиксации глаза, обращена строго к стимулу. Каждое смещение стимула из зоны фиксации вызывает компенсационный поворот тела, "возвращающий" стимул в положение фиксации. Телотаксис позволяет избирать направление исключительно в сторону интересующего стимула, а не от него. Телотаксис сохраняется у насекомого после закрашивания одного из двух симметричных глаз. Поэтому можно думать, что в основе его лежит сложный монолатеральный баланс возбуждения рецепторных клеток, симметричных относительно других рецепторных клеток, ориентированных строго вперед.
Чаще всего телотаксис направляет зрительные поведенческие реакции насекомых, такие, как ориентация самца бабочки или мухи, преследующего самку; ориентация стрекозы, догоняющей комара; ориентация пчелы, когда она садится на цветок. Другим хорошо проанализированным примером телотаксичесшго наведения может служить уже не раз упоминавшийся нами хватательный бросок богомола. Однако не всегда легко решить, ориентируется ли насекомое в данный момент на основе телотаксиса или тропо-таксиса. Известны случаи, когда у одного и того же насекомого, например пчелы, в разных условиях и в разное время жизни проявляется либо телотаксис, либо тропотаксис. Изменяется способ ориентации и в зависимости от того, какой участок глаза возбужден, поскольку для телотаксиса пригодна только его фовеальная часть.
Менотаксис - наведение на цель путем выбора и сохранения некоторого постоянного угла между продольной осью тела и направлением на какой-либо посторонний (случайный) стимул-ориентир, например источник света или звука. Менотаксическая ориентация по существу аналогична ориентации по компасу или движению по азимуту, изобретенному человекам, и, строго говоря, включает в себя телотаксис, который можно рассматривать частным случаем менотаксиса, когда угол между направлением на ориентирующий стимул и продольной осью тела равен нулю.
Менотаксис открывает насекомому принципиально иные, отличные от других таксисов возможности ориентации в пути или в покое. При этом нет необходимости ориентироваться непосредственно по привлекающему или отталкивающему стимулу, а можно выбрать любой посторонний ориентир или веху. В качестве последней служат источник света (солнце, небо, луна) или броский, выделяющийся среди других предмет, а также источник звука, направление ветра или течения, направление силы гравитации.
По-видимому, особенно широко прибегают насекомые к мено-таксической ориентации по отношению к оптическим вехам. Их фасеточный глаз является удобным угломерным инструментом, позволяющим, например медоносной пчеле, вести отсчет углов с точностью вплоть до 1°. Ориентация по солнцу впервые была замечена у муравьев, затем астрономическую ориентацию подробно изучали у медоносной пчелы, клопа Velia и многих других насекомых, способных избирать в качестве вехи не только солнце, но и направление поляризации света голубого неба.
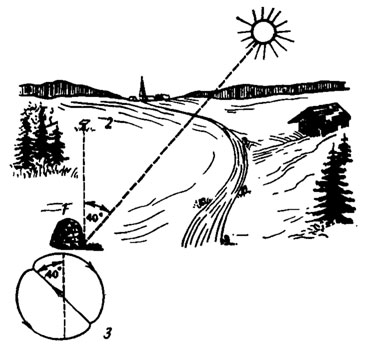
Рис. 68. Указание направления медоносной пчелой Apis mellifera, 'танцующей' на вертикальных сотах (по Frisch von,1956-1957). 1 - улей; 2 - место кормления; 3 - схема виляющего 'танца' пчелы, прямолинейный пробег которого отклонен на 400 слева от вертикали и тем указывает на цель под углом 40° слева от солнца
Большой интерес представляет способность насекомых транспонировать (переносить) менотаксический угол ориентации с одного ориентира на другой, например переносить угол относительно источника света к направлению силы тяжести и наоборот. Когда насекомое попадает в темноту, то на наклонной поверхности оно ползет под определенным углом к направлению силы гравитации, зависящем от предшествовавшего светового угла ориентации. Медоносная пчела, как это показано К. Фришем, исполняет так называемый виляющий "танец" на вертикальных сотах улья под тем же углом к гравитации, под которым она видела солнце во время полета к источнику пищи (рис. 68). Полет в направлении к солнцу здесь отмечается направлением "танца" прямо вверх. Отклонение в ту или другую сторону от направления на солнце передается равным угловым отклонением влево или вправо от направления вверх. Присутствующие при "танце" разведчицы пчелы-фуражиры, покидая улей, поступают наоборот: транспонируют увиденный геоменотаксический угол ориентации к углу фотоменотаксическому.
Экспериментально установлено, что геоменотаксические углы пчела отсчитывает главным образом по степени наклона головы относительно груди, регистрируемого проприоцепторами в области шеи: дорсальными и вентральными волосковыми пластинками. Когда пчела стоит на горизонтальной поверхности, те и другие волосковые пластинки равномерно прижаты головой. Когда пчела ползет вверх, то голова наклоняется вентрально вследствие ее низкого центра тяжести. В этом положении вентральные волосковые пластинки сдавливаются сильнее дорсальных, причем тем больше, чем круче подъем. Когда же пчела ползет вниз, то максимально возбуждаются дорсальные пластинки. Причастность этого механизма к оценке гравитационных углов доказывают опыты с перерезанием нервов, идущих от волосковых пластинок, или изменением положения центра тяжести головы (дополнительным грузом), после чего ухудшалась способность пчелы ориентировать свой "танец" относительно направления силы тяжести.
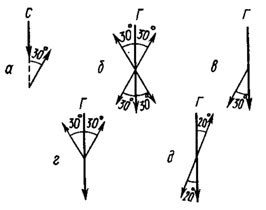
Рис, 69. Перенос менотаксического светового угла (а) к гравитационному муравьем Myrmiса (б), жуком Geotrupes (в), жуком Coccinella (г) и ручейником Limnophilus (д) (по Jander, 1957; Frisch, Lindauer, 1963). С - направление светового луча; Г - направление силы тяжести. Тонкими стрелками показано направление движения насекомых под соответствующими углами к С и Г
Поворачивая в вертикальное положение поверхность, по которой ползет муравей Myrmica, ориентирующийся фотоменотаксически по лампе, можно испытать его способность к транспонированию угла ориентации, если выключить свет в помещении. Оказалось, что для муравья световой угол равняется четырем зеркально-симметричным гравитационным углам (рис. 69, б). В аналогичных опытах у жука Coccinella (г) обнаружены два зеркально-симметричных гравитационных угла, а у другого жука - Geotrupes (в) - один, причем при ориентации головой вниз. Совсем иным оказался перенос углов ручейником Limnophilus (д), который относит световой угол к гравитационному как 3:2, т. е. не соответственно, как другие насекомые, а пропорционально.
Исходный биологический смысл переноса менотаксических углов с одного ориентира на другой состоит в том, чтобы в неожиданно изменившейся обстановке удержать прямолинейное движение, которое дает больше шансов найти "выход", чем неориентированное хаотичное движение. Последнее в силу всегда существующей незначительной асимметрии локомоторного аппарата неизбежно выведет насекомое на круговую траекторию. Известно, например, что заблудившийся человек описывает круги именно по этой причине. Но у медоносной пчелы транспонирование ориентационных углов приобрело сигнальное значение: направлением "танца" разведчица сообщает другим членам общины о направлении полета за взятком или к новому улью (в период роения).
Способность переносить угол световой ориентации к гравитационному углу и наоборот прямо указывает на существование в центральной нервной системе насекомого единого центра менотаксиса, являющегося, возможно, общим для всех балансных таксисов центром управления. Топография и морфология этого центра пока точно неизвестны. Предположительно его отождествляют с грибовидными телами переднего мозга, где прослеживаются волокна нейронов - детекторов движения, безусловно входящие в состав физиологического механизма, таксисов. Хорошим примером внешнего выражения сложности такого управляющего центра может служить, например, внесение поправки к курсу на смещение солнца, что необходимо насекомым при астрономической ориентации.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'