14.2. Число самок и экология вида
Степень обособленности отдельной семьи зависит от числа имеющихся в ней яйцекладущих самок (маток). Моногинные семьи всегда очень резко обособлены, и их члены самоотверженно защищают свое гнездо от всех животных, за исключением некоторых паразитов, которым удается в него проникнуть. Полигинные же семьи весьма различны в этом отношении: некоторые защищают свое гнездо во всех случаях, а другие - только от представителей иных видов. У ос число маток зависит от климата: род Polistes в значительной степени полигенен в тропиках, но моногинен в умеренных областях (Yoshikawa, 1963). Обычно это объясняют тем, что в тропиках такие хищники, как муравьи и птицы, вездесущи и что при наличии нескольких маток и большого числа рабочих особей осиная семья может в случае необходимости разделиться, перелететь на другое место и начать все сначала. В умеренных областях необходимость пережить долгую зиму дает преимущество одиночным самкам, так как они могут впасть в диапаузу и перезимовать в сухих прохладных местах, а затем начать самостоятельно строить гнездо. Кроме того, весной корма очень мало и он распределен диффузно, а поэтому очень важно, чтобы насекомые были сильно рассредоточены. У одного из видов Mischocyttarus, живущего во Флориде, где, конечно, тепло и сезонные колебания климата незначительны, семьи бывают моногинными весной, но полигинными осенью. Однако даже в тропиках роящиеся Polybiini могут периодически становиться моногинными, как, например, Metapolybia aztecoides, и создается впечатление, что это связано с такими более серьезными причинами, как необходимость естественного отбора на уровне семей. Возможно, что потребность в половом размножении, расселении и отборе наиболее приспособленных семей носит периодический характер. Численность семьи у полигинных Polistes достигает 20 000 особей (тогда как у тропических моногинных Polistes - лишь около 200), и у них выработалась способность к обмену информацией, необходимому для того, чтобы стимулировать образование роя (Jeanne, 1981). Враждебность между матками Vespula germanica возникает только в тех случаях, когда они выходят из диапаузы после прохладной зимы; Spradbery (1973b) отмечает, что на Тасмании, где зимы теплые, матки относятся друг к другу дружелюбно и образуют гигантские семьи.
Род Bombus сходен с Polistes тем, что в нем тоже известны виды (хотя их относительно немного), которые в тропиках образуют полигинные семьи (например, В. atratus), хотя, кроме того, самки основывают семьи и в одиночку (Sakagami, 1967а). Для рода Apis характерны моногинные семьи, а вид Apis mellifera, распространенный по всему земному шару, как в тропиках, так и в умеренных областях, повсеместно является строго моногинным. То же самое можно сказать о Meliponini, у которых репродуктивные самки находятся в запечатанных ячейках, пока не наступит время для соревнования между ними или не будет найдено место для нового гнезда. Очевидно, одна матка со значительными возможностями к экспансии может обеспечить семье все необходимое количество яиц, а слишком крупные семьи невыгодны - целесообразнее разделяться и расселяться.
В то время как для ос моногиния - это один из факторов, позволяющих им проникнуть в умеренные зоны, для муравьев эта проблема не связана с климатом. Моно- и полигинные муравьи встречаются во всех областях земного шара; иногда полигинным бывает даже один из видов-двойников или же полигиния образует пятно в популяции, которая в целом моногинна. У моногинных видов крылатые самки способны находить новые места и основывать новые семьи в одиночку, за счет собственных резервов; они могут поэтому выступать в роли первопроходцев, осваивая новые местообитания, в частности начальные стадии сукцессии. Полигинные формы могут повысить плодовитость уже существующей семьи быстрее, так что она растет и расселяется, но поскольку самки зависят от неспособных летать рабочих особей, они все время вынуждены передвигаться по земле. Таким образом, если у моногинных форм самка независима, самостоятельна и способна проникать в новые местообитания по воздуху, то у полигинных форм самки зависят от рабочих особей и ограничены в своем распространении теми новыми местообитаниями, до которых рабочие могут добраться по земле. Однако пешее передвижение дает им важное преимущество, так как в плотно заселенных местах рабочие могут испытывать и оценивать интенсивность и природу конкуренции и затем, если будет нужно, покидать эти места. Одни идут на риск, а другие действуют осмотрительно.
В идеальном случае вид оставляет за собой обе возможности, используя ту или другую в зависимости от обстоятельств, подобно тому как растения производят семена (моногиния) и распространяются, образуя корневища (полигиния). Накапливается, однако, все больше данных о том, что один и тот же вид часто дивергирует по этим признакам, т. е. происходит экологическое разделение его на виды-двойники, один из которых распространяется по воздуху, а другой - по земле, один захватывает свободные участки, а другой медленно пробивается в занятые местообитания. Приведем теперь несколько примеров.
14.2.1. Два вида Pseudomyrmex
В тропических районах Америки оба вида живут в стеблях Acacia: у одного, Pseudomyrmex ferruginea, в семье бывает по одной самке, а у другого, P. venefica, - по многу (Janzen, 1973): Самцы P. ferruginea собираются около какого-нибудь хорошо заметного дерева, к которому примерно в полночь слетаются самки, тогда как самцы P. venefica не собираются вовсе, а обследуют листву деревьев за несколько часов до рассвета в поисках привлекающих их самок. У обоих видов копуляция происходит только один раз: у Р. ferruginea - на земле около дерева, у которого собираются самцы, а у P. venefica - на "брачном" дереве. Затем самки обламывают свои крылья и начинают передвигаться по земле. Ни один из этих видов не летит на поиски нового места. Самки P. ferruginea ищут молодые растения акации и часто уходят за сотни метров от места сборища; успех этих поисков может в значительной мере зависеть от того, устроят ли самцы свое сборище в подходящем месте. В отличие от этого самки P. venefica стараются вернуться обратно на свое собственное дерево, но могут встретить сопротивление со стороны рабочих особей, которые не дают им войти через существующие отверстия и не позволяют проделать новые отверстия. После нескольких попыток самки падают на землю и начинают искать новые побеги, а не проростки. Ясно, что самая важная проблема состоит для них в том, примут их обратно рабочие особи или нет; факторы, от которых это зависит, неизвестны, но, вероятно, здесь играет роль численное отношение рабочие/самки в семье, а также генотип молодой самки. У P. ferruginea самки настроены очень враждебно друг к другу и вступают в драки из-за проростков акации, которые они не хотят использовать совместно, даже временно. Рабочие особи тоже проявляют агрессивность по отношению к любой самке, кроме собственной матери, и к рабочим из любой другой семьи; при схватках они кусают друг друга и мажут ядом. В тех местах, где кроны акаций, разрастаясь, перекрываются, между семьями часто происходят беспощадные побоища. В отличие от этого самки P. venefica не пытаются устраивать гнезда в проростках; у них выработалось иное поведение: они отыскивают корневые побеги на растениях, размножающихся вегетативно. Рабочие особи P. venefica относятся гораздо менее враждебно не только к другим насекомым, но и к чужим особям своего вида; если в результате роста кустов, на которых живут эти муравьи, они встречаются с представителями другой семьи, то они не конфликтуют, а просто смешиваются и сливаются в единую группу.
Семьи Pseudomyrmex ferrugineus медленно растут в своих проростках, но обеспечивают им полную защиту от других насекомых, повышая тем самым вероятность быстрого возобновления ресурсов, необходимых самим этим муравьям. Зрелые семьи, в которых бывает до 30000 рабочих особей, производят самцов, когда численность взрослых особей достигает 1-2 тысяч, а затем производят как самцов, так и крылатых самок. Самка и рабочие, которые живут в особенно крупных полых колючках акации, переселяются в новые колючки примерно трижды в год; их нельзя считать полностью оседлыми. Самка перед этой миграцией прекращает откладку яиц, так что она может легко передвигаться и проникать в новые колючки. Рабочие особи сопровождают ее и могут даже переходить по земле от одного дерева к другому. Это, конечно, рискованно: если самку съест какая-нибудь птица, то вся семья погибнет, так как рабочие особи не принимают новую самку и не выращивают заместительницу, а остатки семьи в течение некоторого времени производят самцов. Семьи P. venefica растут быстрее, повышают свой репродуктивный потенциал, принимая обратно самок и перетаскивая с места на место яйца, и они способны поспевать за более быстрым ростом дерева, в котором обосновались, хотя защищают его менее активно. Границ между территориями разных семей у них нет, поэтому численность отдельной семьи определить невозможно; Janzen считает, что на нескольких тысячах старых деревьев акации могут мирно существовать четыре миллиона муравьев. Хотя между семьями нет преград, они способны образовывать секции без самок, в которых обитает несколько сотен рабочих, выращивающих половых особей.
Кроме того, как отмечает Janzen, самки у этих двух видов различаются по размерам, а также по поведению. Самки Pseudomyrmex ferrugineus - хорошие летуны с сильно развитыми крыловыми мышцами, которые они используют в качестве пищевого резерва в период основания семьи. Их жвалы тоже очень мощны и легко прогрызают колючку. Очевидно, размеры и сила, в частности развитие крыловых мышц и жвал, способствуют успеху. Самки P. venefica мельче и слабее, и их успех определяется такими более тонкими свойствами, как способность привлекать брачных партнеров, умиротворять рабочих-привратниц и получать от рабочих особей корм. Это дает им возможность откладывать больше яиц, производить больше рабочих и оказывать влияние на рост и воспроизведение семьи. Мало вероятно, что рабочие особи в самом деле оценивают способность самки к откладке яиц, но вполне возможно, что какой-нибудь сцепленный с этим признак (например, уровень активности и энергии, покорность и привлекательность), коррелятивно связанный с пригодностью к роли производительницы, периодически подвергается оценке. Распределение самок P. venefica по величине бимодально, и Janzen заметил, что мелким самкам легче вторично проникнуть в свои колючки, чем более крупным (рис. 14.1). Такого рода распределение по величине встречается у муравьев нередко, причем более мелкие самки всегда очень близки по размерам к самым крупным рабочим особям; их называют "микрогинами".
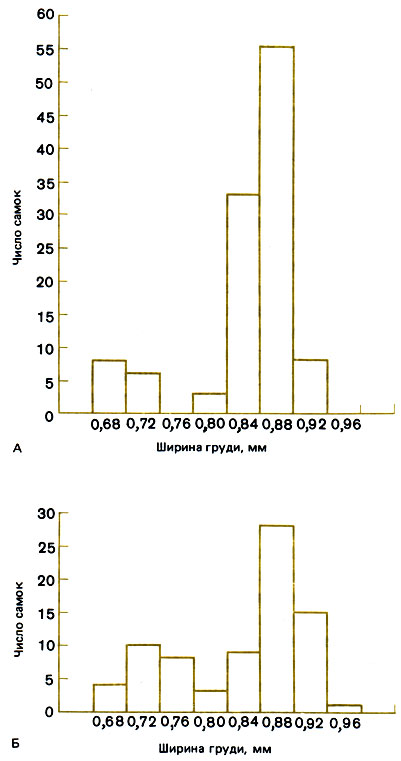
Рис. 14.1. Ширина груди у репродуктивных самок (А) и у маток (Б) Pseudomyrmex venefica, обитающих в колючках акации. (Janzen, 1973.)
Итак, эти два обособленных вида живут оба в тропическом поясе Нового Света в колючках акации, но не скрещиваются друг с другом; собственно, скрещивание между ними невозможно - настолько сильно они различаются по своей биологии и по местообитаниям. Один вид живет на акации, размножающейся семенами, а другой - на акации, размножающейся вегетативным путем (с помощью корневых побегов). Соответственно у каждого из этих видов выработался способ размножения, аналогичный способу размножения растения-хозяина: в первом случае это расселение в одиночку крупных, перегруженных резервными веществами самок - основательниц, во втором -расселение отводками, состоящими из мелких самок и рабочих особей, без которых эти самки не могут обойтись. Janzen указывает, что сами эти растения встречаются в различных, хотя и в значительной степени перекрывающихся местообитаниях: растения, которые размножаются семенами, предпочитают более влажные места, чем те, которые размножаются корневыми побегами.
14.2.2. Виды Myrmica
Пары специализированных сходным образом видов-двойников широко известны у муравьев, хотя они обычно бывают приспособлены к менее специфичным местообитаниям, чем те, которые мы только что рассмотрели. Одна такая хорошо изученная пара - это две формы Myrmica ruginodis, которые можно считать видами-двойниками. У одной формы (macrogyna, М) обычные самки, способные в одиночку основать семью и образующие перманентные группы, в которые входит до шести самок. У другой (microgyna, m) - мелкие самки, или микрогины, неспособные самостоятельно основать семью, так что они отделяются в сопровождении рабочих особей. Их семьи расселяются путем почкования и время от времени принимают новых самок. М производят "нетерпимых" рабочих особей, которые не примут новую самку, даже если у них нет своей; m толерантны и легко смешиваются с представителями своего вида, но не допускают к себе ни одну особь М. Для М характерен аутбридинг; у них происходит брачный лёт, часто на скалистых хребтах, разделяющих долины; самцы собираются у хорошо заметных скал и деревьев, к которым прилетают самки для копуляции. Эти же места могут использовать и т, но они большей частью копулируют поблизости от гнезда и производят половых особей лишь раз в несколько лет. Эта периодичность в сочетании с ясно выраженным предпочтением брачных партнеров собственного вида препятствует гибридизации (Brian, Brian, 1949, 1955).
Описанные репродуктивные различия приводят к тому, что эти две формы живут в разных стациях своего влажного местообитания; обычно это верещатники на кислых почвах (Elmes, 1978b). Распространение М шире, и они могут колонизировать многие совершенно обособленные участки, как, например, в центрах куртин вереска (Calluna vulgaris), которые разрослись и покрылись мхом, или на маленьких залитых солнцем лесных полянках, тогда как т обитают в таких местах, где растительность часто выедается скотом. Elmes, обследовав 18 мест в Англии, в 14 из которых обитали обе формы, нашел, что местообитания, способные предоставить все необходимое нескольким крупным семьям, заняты видом М, а те, которые подходят для многочисленных небольших, сильно разбросанных секций, были заняты видом т; эти последние стации были более богаты злаками и регулярно выедались скотом. Расселение этой формы, несомненно, происходит организованно и при участии всего сообщества.
Эту пару видов рассматривали как единый вид, потому что их нельзя уверенно отличить друг от друга даже по размерам самок; однако рабочие особи распознают самок этих двух форм (видимо, по запаху), хотя ширина головы у них одинакова. Обе формы встречаются вместе по всей палеарктической области от Британских островов до Японии, так что это не подвиды. Форма с мелкими самками размножается в чистоте, а ее рабочие особи лишь ненамного мельче, чем у формы с крупными самками. Вероятное объяснение состоит в том, что форма m возникла в гнездах формы М, основанных достаточно давно, чтобы в них в результате мутаций и инбридинга могла возникнуть микрогинная форма, начавшая свое существование в качестве социального паразита; в дальнейшем, если местообитание может это выдержать и если форма М вымирает, форма m сменяет ее (Pearson, 1981; Pearson, Child, 1980). Обследование Elmes показало, что местообитания этого типа привлекают к себе и другие виды Myrmica, так что форме m приходится существовать в условиях конкуренции, которым хорошо соответствует их расселение отводками. Таким образом, создается впечатление, что у М. ruginodis происходит симпатрическое видообразование и притом непрерывно; форма М адаптирована к жизни в местах с постоянно изменяющимися, эфемерными условиями, к которым устремляются в одиночку ее самки, тогда как форма т приспособлена к стабильным местообитаниям, в которые она расселяется, образуя отводки. Редкость промежуточных форм указывает, вероятно, на то, что они находятся в смысле воспроизведения в невыгодном положении по сравнению как с М, так и с m и поэтому почти полностью отметаются отбором.
Однажды был построен график личиночного развития одной микрогины (рис. 14.2). У этой особи наблюдалось несколько остановок роста, однако ее размеры все же превысили обычные для личинки рабочей особи, хотя и не достигли размеров, обычных для личинки репродуктивной самки на решающей стадии определения касты. Возможно поэтому, что достаточно совсем простого генетического изменения, влияющего на рост личинки, чтобы из нее получилась микрогина, а этого можно ожидать в давно основанной популяции, которая стала изолированной и размножается путем инбридинга. Если только микрогины смогут оставаться в семье, где они возникли, или возвращаться в нее, то, по-видимому, нет причин, почему временный социальный паразитизм не мог бы продолжаться бесконечно.
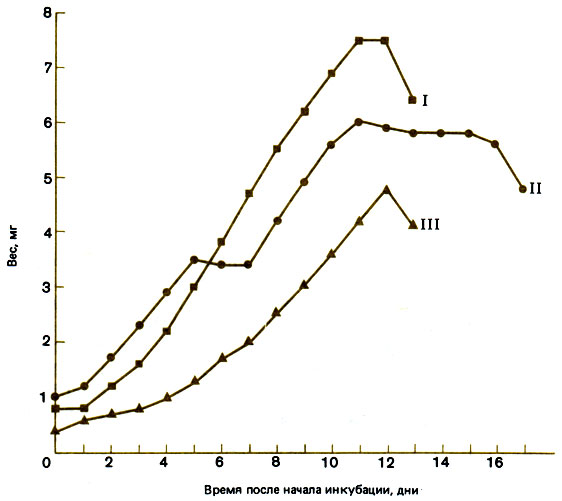
Рис. 14.2. Рост личинок после зимовки. Некоторые из них развились в репродуктивных самок (I), одна - в микрогину (II), а некоторые - в рабочих особей (III). (По данным Brian, 1955b.)
Myrmica rubra - вид, очень сходный с М. ruginodis - это настоящий полигинный вид со средним числом самок в семье, равным 16, и отрицательным биномиальным распределением их по весу тела (рис. 14.3; Elmes, 1973). Отдельные самки могут основывать семьи в одиночку, во всяком случае в лабораторных условиях (Brian, 1951), однако более вероятно, что в природе они основывают семьи небольшими группами, сопровождаемые рабочими особями. У этого вида часть молодых самок, несомненно, возвращается назад в свою семью, а возможно, принимаются также самки и из других семей. Elmes (1973) обнаружил обратную зависимость между числом самок и их размерами, которая может быть обусловлена перенаселенностью в период их выращивания, генетически детерминированным уменьшением их величины в крупных, давно основанных семьях или тенденцией принимать обратно в гнездо мелких, но не крупных самок (как у Pseudomyrmex venefica). Таким образом, М. rubra в отличие от пары видов М. ruginodis представляет собой один вид с обычными самками, способный расселяться двумя способами: во-первых, самки могут улетать и основывать новые семьи без посторонней помощи; во-вторых, семья может пополняться молодыми самками и отпочковывать отводки, которые по мере созревания обособляются и становятся самостоятельными семьями. М. rubra производит большое число микрогин, так что иногда в семье их может быть много наряду с обычными самками. Поодиночке микрогин никогда не находили (Elmes, 1976), и они, по-видимому, не в состоянии создавать жизнеспособные семьи. Одна из причин этого-то, что микрогинам трудно производить рабочих особей: откладываемые ими яйца не детерминированы к развитию в рабочих, и они не могут регулировать выращивание расплода рабочими. Таким образом, микрогины обречены существовать как паразиты особей нормального типа.
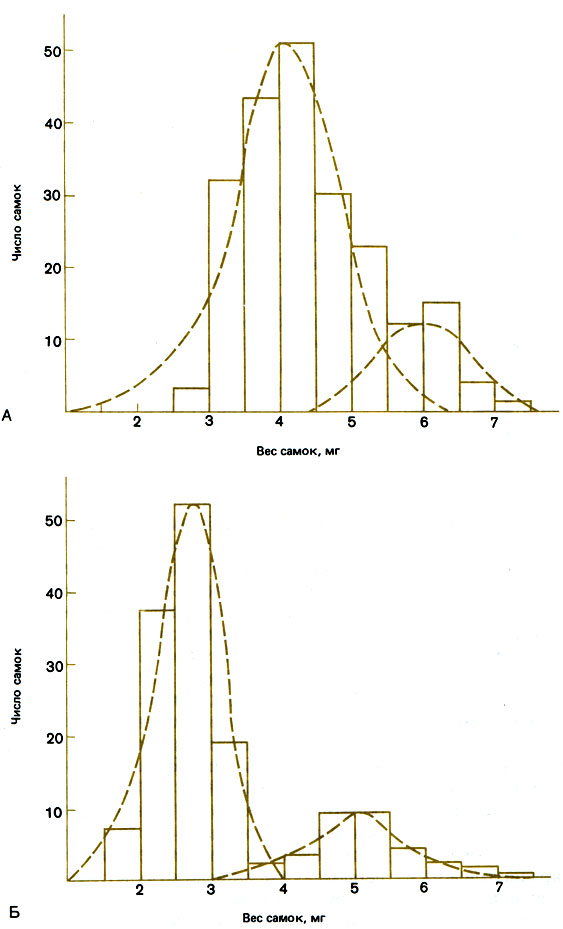
Рис. 14.3. Число самок, имеющих данный сырой вес, у Myrmica ruginodis (А) и М. rubra (Б). Графики показывают резкое различие в весе между микрогинами (слева) и нормальными самками (справа). Прерывистыми линиями показаны гипотетические распределения внутри общей бимодальной кривой. Обратите внимание на то, что мода для нормальных самок М. rubra меньше, чем для нормальных самок М. ruginodis, примерно на 1 мг, а для микрогин эта разница даже несколько превышает 1 мг; иными словами, эти микрогины очень мелкие - фактически они так малы, что не могут производить рабочих особей. (Elmes, 1975.)
Этим дело не кончается. У другого вида Myrmica - М. sabuleti - есть социальный паразит, не имеющий касты рабочих особей; это M. hirsuta, которого раньше считали спорадической микрогиной. Более того, возможно, что все не имеющие рабочих социальные паразиты, объединяемые теперь в род Sifolinia, возникли таким образом, поскольку этот род морфологически очень близок к Myrmica (Elmes, 1878а). Это, по-видимому, логическое следствие преимущественного принятия в семью мелких самок, как у Pseudomyrmex venefica. Им может оказаться трудно производить рабочих особей, и они превращаются в социальных паразитов. Микрогинная форма М. ruginodis, способная вести относительно независимое существование, несомненно, представляет собой исключение. Полигиния может выродиться в социальный паразитизм и привести к вымиранию в результате сверхспециализации.
Принадлежащие к тому же подсемейству часто встречающиеся роды Leptothorax и Monomorium содержат полигинные виды, но такие процветающие роды, как Tetramorium, Solenopsis и Pheidole, а также все Attini обычно моногинны.
У завезенного в Северную Америку огненного муравья Solenopsis invicta, как теперь известно, кое-где встречаются полигинные семьи. В одной особенно большой семье с 20000 осемененных самок (Glancey et al., 1975, 1976) проявлялись признаки инбридинга, о котором свидетельствовала высокая доля диплоидных самцов: у 96% изученных самцов семенников не было или они были развиты слабо, а сперма отсутствовала, тогда как в обычных семьях с гаплоидными самцами стерильность самцов составляла менее 1%. Несомненно, этот инбридинг вел и к полигинии. Увеличение доли диплоидных самцов совместимо с гипотезой о том, что пол определяется генами, лежащими в одном половом локусе, хотя в данном случае у вида Solenopsis, завезенного из Южной Америки, могла резко уменьшиться генетическая изменчивость, так что некоторые семьи могли стать гомозиготными по нескольким главным локусам, определяющим пол (Crozier, 1971). В штате Миссисипи найдено 15 полигинных семей (от 3 до 63 самок), а в штате Джорджия их было больше; 90% самок были осеменены и откладывали яйца, хотя и не в таком количестве, как нормальные крупные физогастрические самки моногинных семей (Fletcher et al., 1980). Полагают, что полигиния у этого вида усиливается; по всей вероятности, когда он впервые попал в Северную Америку, в его семьях было по одной самке.
14.2.3. Dorylinae и Formicinae
Rettenmeyer и Watkins (1978), специально занимавшиеся группой Dorylinae в Америке и изучившие 144 вида этого подсемейства, обнаружили лишь один вид, в семьях которого было более одной самки, - Neivamyrmex carolinensis с числом самок от 3 до 13. По мнению этих авторов, полигиния может быть полезна муравьям при их подземных миграциях в климате с резкими сезонными изменениями, но существует много видов подземных Dorylinae, которые не полигинны.
Среди Formicinae многие роды строго моногинны (например, Oecophylla), но в других родах (например, Lasius) есть виды, образующие кое-где полигинные семьи, а есть и целиком полигинные виды. В семьях Lasius sakagami, по морфологии и поведению очень близкого к L. niger, от 0 до 309 самок (Yamauchi et al., 1981). L. sakagami копулируют в гнезде или около него, но крылатые самки появляются не каждый год, в отличие от L. niger. Самки L. sakagami либо возвращаются в гнездо, либо улетают (в последнем случае, по-видимому, при низкой плотности) и, подобно L. niger, начинают строить новое гнездо. Самки L. sakagami менее плодовиты, но их рабочие особи толерантны к ним и принимают их обратно в гнездо. В результате семьи у этого вида сильно разрастаются и содержат миллионы, а не десятки тысяч особей. Местообитания L. sakagami очень ограниченны: эти муравьи жувут на песчаной почве со скудной травянистой растительностью и строят подземные гнезда, которые в отличие от гнезд L. niger устойчивы к затоплению (ср. L. flavus). L. niger гораздо более эвритопный вид. Таким образом, L. sakagami сохраняет способность как к моно-, так и к полигинии (подобно Myrmica rubra), тогда как L. niger - строго моногинный вид.
14.2.4. Род Formica
Этот род очень интересен с точки зрения числа самок в семье. Экологически наиболее примитивный вид Formica fusca совершенно независим от других, но наряду со своими близкими родичами (например, F. rufibarbis, F. neogagates) служит хозяином многим муравьям из группы F. rufa, а также муравьям-рабовладельцам F. sanguined и Polyergus rufescens. Сам муравей F. fusca - олигогинный вид в Европе (Pamilo et al., 1978, 1979), а в Японии варьирует от моногинного вблизи Токио до перманентно полигинного в другой части Японии (Higashi, 1979)*. У моногинной формы наблюдается сезонная полигиния, создаваемая в результате возвращения в конце лета в семью недавно спарившихся самок (Kondoh, 1968). По окончании зимы этих самок изгоняют, часто с группой рабочих особей, чтобы они основали новые гнезда, и все же с приближением следующей зимы их могут вновь принять назад. Эта сезонная полигиния напоминает полигинию, наблюдаемую у некоторых тропических Polybiini. Давно известно, что в Северной Америке семьи F. pallidefulva могут быть как моно-, так и полигинными (Talbot, 1946; обсуждение см. Hölldobler, Wilson, 1977b, p. 13). Особенно интересна группа видов F. microgyna, у которой бывают только микрогинные самки, закладывающие новые семьи путем временного социального паразитизма и использующие F.fusca, F. neogagates и F. pallidefulva в качестве хозяев. Вид F. microgyna широко распространен и варьирует по хетотаксии, но не обладает другими хорошими диагностическими признаками (Letendre, Huot, 1972). Не произошел ли он от какого-то хозяина вроде F. fusca? Ряд все возрастающей зависимости от хозяина завершает вид F. talbotae - не имеющий рабочих особей социальный паразит, который даже еще мельче и имеет больше волосков, чем типичная F. microgyna (Wilson, 1976с). Он использует в качестве хозяина F. obscuripes (из группы F. rufa); нет нужды подчеркивать параллелизм с Myrmica hirsuta.
*(В настоящее время дальневосточная форма, обитающая, в частности, в Японии, рассматривается как самостоятельный вид F. japonica. - Прим. ред.)
В группе F. rufa имеются разнообразные виды -от моногинных до полигинных. Сам вид F. rufa представлен моногинной формой на Европейском континенте (Collingwood, 1979), но олигогинной - в Англии*. Морфологически близкий к нему вид F. polyctena ограничен материком, и в его колонии, объединяющей 50-100 гнезд на площади в 30 га, может быть тысяча и более мелких самок (Raignier, 1948). Этот вид легче принимает новых самок поздней осенью или ранней весной, а также сразу после того, как от семьи начал отпочковываться зачаток новой семьи (Elton, 1958). Вопрос о том, существуют ли семьи, несомненно защищающие свои территории, вызывает разногласия; защита муравьями своих границ, по-видимому, зависит от плотности популяции и от истории локального местообитания. Mabelis (1979а) подробно описывает схватки, возникающие при защите границ, и указывает, что они особенно часты весной, когда семьи вновь устанавливают свои территории после зимней спячки. Разделение семьи с образованием одного или двух новых гнезд за раз может занять больше месяца и обычно начинается под действием таких неприятных факторов, как затенение, обнаружение того, что гнездо находится недостаточно близко к центру данной территории, или вторжение других видов (Mabelis, 1979b). Рабочих особей и расплод переносят при этом взад и вперед, но в среднем большая часть семьи переходит в новое гнездо; самка либо переходит сама, либо ее переносят, но уничтожение самок хищниками у полигинных видов не приводит к таким катастрофическим последствиям, как у моногинных. Процент выживающих гнезд не очень велик; по данным Klimetzek (1981), обновляется от 8 до 32% гнезд. Welch (1978) выявил значительное обновление гнезд F. rufa в Англии.
*(В СССР найдены полигамные семьи F. rufa. - Прим. ред.)
У Formica opaciventris - одного из представителей группы exsecta в Северной Америке* - новые семьи образуются путем почкования (Scherba, 1964); F. exsecta в Польше - полигинный муравей, которого много лет подробно изучал Pisarski (1973)**. Семьи F. pratensis в Евразии по большей части моногинны, но в Сибири иногда их гнездо состоит из центра, который окружен территорией, перерезанной отходящими от центра тоннелями, куда направляются рабочие и где они ожидают сигналов, мобилизующих их на поиски пищи; в определенное время года эти находящиеся в тоннелях группы приобретают собственных самок и отделяются (Резникова, 1979). В части ФРГ и в Нидерландах этот вид полигенен и обитает в тенистых лесах, в связи с чем половое созревание наступает у него позднее, чем в моногинных семьях, живущих на открытых местах; эти различия в сроках созревания снижают частоту скрещивания между разными семьями (Collingwood, 1979). Вид F. lugubris в Ирландии моногинен или слегка полигинен, с несколько чрезмерно разбросанными гнездами, существующими по 23-38 лет (Вrееn, 1977). Особенно большой комплекс гнезд был найден в лесах Юры (Франция-Швейцария) на высоте более 1300 м, где с октября до конца мая лежит снег и ежегодно выпадает около 2 м осадков, а среднегодовая температура всего 4-5°С. Этот комплекс занимает площадь 70 га, и в него входит 1200 гнезд, обитатели которых настроены враждебно по отношению к другим полигинным группам на их границах (Cherix, 1980; Cherix, Gris, 1977). Столь же обширный комплекс Formica truncorum известен в Японии; в этой колонии рабочие особи тоже совершают переходы (или их переносят) от одного гнезда к другому, но отдельные особи лишь в редких случаях проходят больше 4 м (Higashi, 1976, 1978).
*(Палеарктические виды Coptoformica ("группа F. execta" по терминологии автора) не родственны неарктическим видам из группы F. exectoides.)
**( Писарский обнаружил в Польше две формы F. execta - полигинную и моногинную. - Прим. ред.)
Популяционная генетика европейских видов Formica изучалась методом изозимного анализа (Pamilo et al., 1978). Географическая изменчивость у такого вида, как F. sanguinea, очень незначительна, но у полигинного лесного вида F. aquilonia, имеющего разорванный ареал, один и тот же локус содержит в Финляндии и в Шотландии разные аллели; то же самое обнаружено у F. lugubris. Различия между локальными популяциями обычно невелики: 5,1% у F. polyctena, до 24,7% у F. fusca, хотя у F. aquilonia они превышают 50%. Генное разнообразие очень мало - 0,043 в отличие от 0,150 у диплоидных насекомых (Pamilo et al., 1978, цит. по Selander, 1976), что, несомненно, обусловлено элиминацией рецессивных субвитальных генов у гаплоидов; это приносит пользу, устраняя вредные последствия инбридинга, часто весьма значительного у Formica.
В идеале отдельный вид должен применять смешанную стратегию - иметь достаточно крупных самок, способных основать семью в одиночку, и наряду с этим более мелких, которые могут вернуться в основную семью и отпочковаться, взяв с собой рабочих, при наличии интенсивной конкуренции. Возможно, что так обстоит дело у Myrmica rubra. Однако чаще каждый конкретный вид становится либо моногинным, и самка улетает, чтобы строить гнездо в одиночку, либо полигинным, расселяясь в результате роения (как у М. ruginodis). Не оказалась ли смешанная стратегия неустойчивой в эволюционном смысле? С этой дилеммой сталкиваются многие цветковые растения, которым приходится делать выбор между рассеиванием семян и вегетативным размножением; хотя некоторые из них отдают предпочтение одному или другому из этих способов, есть и такие виды, которые сохранили обе возможности и используют их в подходящее время в подходящих условиях (Harper, 1977). Многие виды Formica выбрали иное решение: их самки либо возвращаются в родное гнездо, либо занимают гнездо другого вида и расширяют свой ареал; самке никогда не приходится основывать семью в одиночку.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'