Экстерьера и внешние условия

Каждый вид животного или растения формировался в зависимости от того, какое он занял "кресло" в бескрайнем "зале" природы, как приспособился к окружающей среде. Она-то и сформировала его внешний вид - экстерьер. При слове "экстерьер" прежде всего обычно вспоминают о животных, внешне резко отличающихся одно от другого. Взять хотя бы наших четвероногих помощников - собак. Отчего экстерьер фокстерьера не похож на экстерьер таксы, лайки, овчарки, дога? Да потому, что каждая из этих пород из поколения в поколение приспосабливалась к тому виду использования, которое определил для нее человек. Именно данный экстерьер, а в соответствии с ним и поведенческие реакции способствовали лучшему осуществлению специальных целей: охота на норную дичь (такса), обнаружение и преследование зверя, быстро передвигающегося в кроне деревьев (лайка), сторожевая служба (овчарка) и т. д. Отсутствие переходных форм здесь легко объяснить вмешательством человека, который четко знал, чего хотел, и твердо вмешивался в наследственность животных, подбирая пары и решая вопрос жизни или смерти удавшегося или неудавшегося, на его взгляд, потомства. Вне контроля человека экстерьер животных лепит среда, то есть условия, в которых приходится жить организму. Выживает тот, у кого форма и поведение оказываются наиболее "прилаженными" к условиям жизни в той или иной экологической нише. Итак, в каждой специфической среде возникают свои жизненные формы. У прямокрылых жизненные формы были описаны в 30-х годах Б. П. Уваровым и Г. Я. Бей-Биенко. Затем Ф. Н. Правдин и его ученики с учетом различий во внешних признаках саранчовых выделили главные жизненные формы: геобионты, или геофилы,- обитатели поверхности земли; фитобионты, или фитофилы, - обитатели растений; хортобионты - обитатели трав; дендрофилы - обитатели деревьев; тамнобионты - обитатели кустарников; псаммофилы - обитатели песка.
Авторы книги на основании детального обмера частей тела саранчовых, хронометрирования их поведения в природе и данных о строении хромосом* предложили новый метод определения жизненных форм саранчовых.
*(Хромосомы - структурные элементы ядра клетки, содержащие ДНК, в которой заключена наследственная информация отдельных признаков, полученных от родителей. Новые признаки, не свойственные отцу и матери, могут появляться в результате мутаций, то есть перестройки ДНК, и сочетания информации об унаследованных признаках. Это так называемая рекомбинативная изменчивость, важнейший механизм которой - перекрест хромосом, приводящий к перераспределению (рекомбинации) локализованных в них генов.)
Оказалось, что все виды дендрофилов и тамнобионтов обладают вальковатым телом, относительно выпуклым профилем, средней длиной усиков и задних ног. Они окрашены в коричневые, желтоватые или землисто-зеленые тона, иногда имеют яркоокрашенные участки ног, служащие как бы сигнальными флажками при общении полов. Это недифференцированные** формы. Почти все их представители отдыхают и двигаются по стволам деревьев, поверхности широких листьев, опускаются на землю только в местах, прикрытых подстилкой, и явно избегают открытой поверхности почвы. Интересно, что подавляющее большинство таких видов склонны к генетически обусловленной изменчивости, связанной с перекрестом хромосом. Таковы представители наименее дифференцированных форм древнего подсемейства катантопин, связанные с тропическими и широколиственными лесами, и очень немногие из других подсемейств, близких к катантопинам.
**(Дифференциация - расчленение целого на части, формы, ступени.)
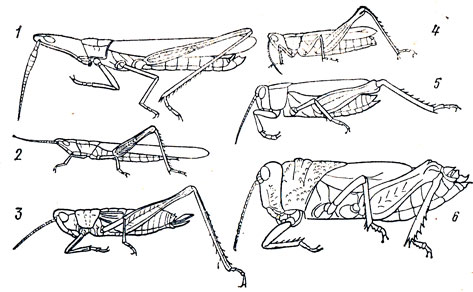
Жизненные формы саранчовых: 1, 2 - осоково-злаковые хортобионты; 3 - специализированный фитофил; 4 - настоящий хортобионт; 5 - факультативный хортобионт; 6 - петро-бионт (обитатель каменистых участков)
Остальных саранчовых можно выстроить в два ряда: обычных, или слабо специализированных, и специализированных. У первых число перекрестов хромосом несколько меньше, чем у недифференцированных форм, а у вторых, приспособленных к строго определенным условиям, рекомбинационная изменчивость встречается значительно реже. По-видимому, за хорошую "подогнанность" к условиям жизни насекомым приходится расплачиваться: изменчивость и видообразование у них ограничены. При резком изменении условий жизни они скорее погибнут, чем изменятся. Так, саранчовые первого ряда среди геобионтов характеризуются расширенным телом лепешкообразной формы, увеличенным лицевым углом профиля до прямого, более или менее толстыми и короткими усиками и задними бедрами. Окраска, насекомых землистая, подчас пестрокаменистая, яркие сигнальные пятна остаются, но перемещаются на внутренние поверхности ног. Насекомые как бы растворяются на открытой поверхности почвы сухих степей и пустынь и делаются очень заметными лишь в тот момент, когда "показывают" себя половому партнеру. При хронометрировании поведения саранчовых выявлено, что они, особенно самцы, почти не обнаруживаются на растениях, а живут на поверхности почвы - на открытых участках.
Специализированные геобионты (то есть формы второго ряда) имеют камнеобразный облик и число перекрестов хромосом в 2,5 раза меньшее, чем у обитающих в лесах недифференцированных форм. Они встречаются в каменистых пустынях центральной части Евразийского континента.
Слабоспециализированные хортобионты с менее резко выраженными признаками называются факультативными (частичными). Типичный факультативный хортобионт - пустынный прус.
У специализированных хортобионтов суженное тело, обостренный лицевой угол, удлиненные усики и бедра. В окраске тела преобладают зеленые, соломенно-желтые цвета и продольный рисунок, что помогает насекомым скрываться среди стеблей и трав. Вместо "размахивания" друг перед другом "цветными флажками" эти обитатели густого травостоя переходят на призывные серенады.
Мысль о конкурентном взаимодействии в эволюции покровительственной и сигнальной окрасок, а также пения впервые высказал создатель службы защиты растений от вредителей в России И. А. Порчинский (1848-1916). Труды ученого, посвященные азиатской саранче, так же как исследования К. Э. Линдемана, помогли выяснить географическое распространение перелетной саранчи, цикл ее развития, основные особенности образа жизни.
Благодаря работам ученых, проводивших хронометрирование поведения саранчовых в природе, мы знаем, что хортобионты почти не покидают растений, а если такая необходимость возникает, то они в мгновение ока перебегают по земле от одного растения к другому. В экспериментальных условиях садка хортобионты стремятся расселиться на моделях стебельков, избегая плоской поверхности садка. Продвигаясь вверх, попав на концы моделей, они легко спускаются по ним, чаще вниз головой. У наиболее специализированных хортобионтов ослабляется способность рекомбинативной изменчивости. При этом не просто уменьшается количество перекрестов хромосом, но и сокращается число самих хромосом с 23 до 17, что характерно, например, для большого рода хортиппусов.
Изучение структуры хромосом у саранчовых разных семейств помогло авторам книги разработать систему жизненных форм этих насекомых, состоящую из 22 групп элементарных форм. В группы объединены все саранчовые, встречающиеся в СССР, то есть почти 200 видов. Система жизненных форм облегчает определение насекомых - обитателей разных ландшафтов, помогает установить эволюционные связи одних видов с другими.
Чуть раньше речь шла о возникновении предков саранчовых в глубине древних тропических лесов. Теперь, опираясь на работы Б. П. Уварова, А. Г. Шарова, Ф. Н. Правдина, Л. Л. Мищенко и собственные исследования, мы можем представить себе, как насекомые вышли из лесов и расселились по молодым, возникшим лишь в третичном периоде травянистым сообществам.
Этим сообществам свойственна огромная скорость круговорота веществ и воспроизведения новых генераций растений, что резко отличает их от древесных сообществ. Так, однолетние травы в конце лета все, что накопили за год в своих "подземельях", выбрасывают в подстилку, как бы становясь частью органического вещества верхнего, пахотного слоя земли. Жизнь деревьев длиннее. Они затрачивают десятки лет на то, чтобы вырасти, созреть, состариться, одряхлеть и превратиться в подстилку. Пока живет одно поколение деревьев, сменяются десятки поколений трав, идет процесс их видообразования, обогащаются сообщества растений, почва наполняется питательными веществами. Все это способствует расширению пищевых связей насекомых-фитофагов, в том числе и саранчовых, улучшает условия их жизни. Травянистые ландшафты с их богатыми почвами (степи, луга, прерии) издавна являются районами наиболее продуктивного земледелия.
Конечно, далекие предки саранчовых "не прошли мимо" возможности поселиться в райских условиях травянистых сообществ степей. Наиболее древние и в то же время наиболее перспективные по возможностям рекомбинативной изменчивости саранчовые появились на полянках уже среди раннетретичных субтропических лесов, которые некогда пересекали территорию нашей страны с востока на запад, а теперь сохранились только на Дальнем Востоке (отчасти также в Западной Европе). В центре же древнего материка Ангариды (северо-восточная часть Евразии), где раньше всего стали изреживаться и исчезать лиственные леса и на смену им приходить заканчивающие свой жизненный цикл за один год травы, начало складываться подсемейство катантопин. Эти насекомые приобрели способность перемещаться по узколистным травам и земле, подобно бескрылой кобылке - Podismа pedestris (группа родов, или триба, подизмини подсемейства катантопин). Чуть позже сформировались и настоящие травообитающие виды из подсемейства акридин, но по приспособительным, поведенческим и кариологическим* признакам еще мало отличающиеся от катантопин. Это представители особой трибы арциптерин (например кобылка пестрая - (Arcyptera fusca), которые распространены сейчас на границе лесов и степей, проходящей через всю Сибирь (иначе говоря, бывшую Ангариду). Большая часть их сосредоточена на луговых опушках рощ и кустарников. Освоение открытых луговых пространств потребовало еще большей специализации саранчовых и повлекло за собой выработку у них фитобионтного облика и поведения. Это вызвало блокировку рекомбинативной изменчивости и уменьшение числа хромосом. Так, по-видимому, возникли род ставродерус и многовидовой род хортиппус, типичный представитель которого - белополосая кобылка (Chorthippus albomarginatus), связанная с горными кустарниковыми степями и субальпийскими высокотравными лугами. Какие-то представители предковых форм в самом начале освоения травянистых ландшафтов Ангариды "скатились" в понижение, занятое сырыми болотами. Здесь и возникла своеобразная группа болотных кобылок - мекостетусов.
*(Кариология - наука о строении ядра клетки.)
Переход к травообитанию происходил не только в середине Ангариды, но и ближе к теплой и засушливой области на берегах древнего океана Тетиса*, существовавшего в мезозое - начале кайнозоя. У северных берегов этого океана, на территории современного Казахстана, происходил переход обитателей лесных полян к жизни среди сухолюбивых кустарников и более приземистых полукустарников (полыни), а также дерновинных сухолюбивых злаков (типа ковылей, типчаков). Здесь, по-видимому, сложилась группа крестовичек (триба доциоставрини), предки которых, возможно, были близки к арциптеринам. Нарастание заслушливости заставляло потомков некоторых крестовичек прятаться в плотных, маленьких кронах полыни - наиболее распространенного растения полупустынь. Там и сформировался своеобразный род крестовичек - эремипус**, само название которого говорит о приспособленности к жизни в пустынных ландшафтах. Такая узкая специализация саранчовых привела к сокращению у них числа хромосом и выработке фитобионтного облика.
*(Остатками Тетиса являются Средиземное, Черное, Каспийское моря, Персидский залив и моря Малайского архипелага.)
**(Эремия - пустыня.)
В юго-западной Азии, в условиях более мягкого и теплого климата, насекомые в процессе эволюции привыкая к жизни на территории, где исчезли леса, заселяли кроны крупных кустов и полыней. Примером могут служить саксауловые горбатки (Dericorys tibialis), относящиеся все к тому же в основе своей лесному подсемейству катантопин, эгнатиусы (Egnatius desertus) из семейства егнатин, близкого к катантопинам. И те и другие - специализированные фитобионты пустыни. Здесь же, по-видимому, складывалась и особая пустынно-степная триба катантопин: насекомые обитают не в самих кронах кустарников и полукустарников, а вблизи их. Это триба калиптамин, или широкоизвестных прусов.
Теперь посмотрим, как возникло подсемейство оедиподин, типичных геофилов, ареал которых связан с берегами древнего Тетиса. Интересно, что группы видов, по характеру кариотипа ближе всего стоящие к лесным катантопинам, оказались переходными от недифференцированных форм одновременно к фито- и геобионтам, причем фитобионтность выражена у них сильнее. Такой особенностью обладают представители трибы эпакромиини (Epacromiini). Их современное распространение в Евразии связано с местами, имеющими разреженную растительность и сырые солончаки на берегах озер. В Африке насекомые трибы эпакромиини встречаются на присаванных опушках тропических лесов и берегах некоторых морей. Видимо, подобные условия существовали на территориях, омываемых Тетисом или оставшимися на его месте изолированными морями. Когда Тетис отступил еще дальше, устьевые части впадавших в него рек начали превращаться в глинистые и песчаные пустыни (например, Каракумы).
Глинистые и песчаные пустыни... По-видимому, именно здесь из предковых форм появились близкие к трибе эпакромиини пустынницы (Sphingonotini) - типичные открытые геофилы.
Из лесов через травянисто-кустарниковые опушки в степи и саванны засушливого облика вышли и когда-то саранчовые трибы локустини подсемейства оедиподин. Сейчас большая часть видов оедиподин обитает в Африке, Индии, Южном Китае. Одни из них перешли к жизни среди тростников на берегах озер (знаменитая перелетная саранча, гнездящаяся в устьях больших рек), другие еще сохранили связь с травянистыми, но хорошо прогреваемыми лесными опушками (трескучая кобылка), третьи широко расселились по саваннам и степям (кобылка украшенная). Несмотря на такое распространение, многие из них сохранили главнейшие особенности строения, свойственные недифференцированным жизненным формам, хотя могут быть отнесены к геобионтам.
Какие-то близкие к ним формы, по-видимому, издавна начали осваивать водораздельные опустыненные степи и пустыни, где и сформировалась триба оедиподини, отличающаяся ярко выраженной геофильностью. Еще больший сдвиг в сторону геобионтности произошел у обитателей каменистых пустынь бессточной области Центральной Азии - представителей трибы бриодемини.
Так формировались саранчовые на территории нашей страны, но сюда же попадали и группы, сложившиеся за ее пределами, например представители подсемейства памфагин (Pamphaginae) пиргомофин (Pyrqomorphinae). Первые из них, по-видимому, возникли миллионы лет назад в сухих климатических условиях гор Восточной и Южной Африки. Вторые образовались в близлежащих районах, но только во влажных местах по берегам рек. И у тех, и у других гораздо раньше, чем у выходцев из Ангариды, по мере специализации уменьшилось число хромосом до 19-ти. В те далекие времена Африка, входившая в огромный континент Гондвану*, была отделена от Евразии Тетисом. Но вот от Гондваны отделился осколок - так называемый Иранский микроконтинент. Он "подплыл" и "припаялся" к Евразии. С него потомки исходных памфагин и пиргоморфин "десантировались" в Евразию и превратились в новые виды, расселившиеся на этом материке. Удивительные по облику саксетании (Saxetania), похожие на комочки почвы, обосновались в горно-каменистых пустынях Армении и Южной Туркмении, другие - на твердых поверхностях глинистых пустынь и по их рекам, третьи проникли на юг степей современного Казахстана и Нижнего Поволжья. Это уже упоминавшиеся нами азиотметисы.
*(Гондвана - единый южный континент, существовавший в палеозойскую эру. Распался на отдельные массивы, из которых сформировались Южная Америка, Африка и Австралия.)
Интересно, что некоторые из этих видов, в том числе связанные с приречными лесами пиргоморфины, остались верны тропической сезонности и зимуют не в виде спрятанных в землю яиц (кубышки), а во взрослом состоянии, что весьма необычно для остальных наших саранчовых.
Читатель вправе спросить: имеет ли какое-нибудь практическое значение эта почти геологическая история? Имеет, и немалое. Зная особенности условий жизни насекомых в местах эволюционного развития триб, родов и жизненных форм, мы можем прогнозировать численность вредителя при изменении этих условий в результате хозяйственной деятельности человека (рубка леса, орошение, осушение земель и т. д.). Причем виды, принадлежащие к узкоспециализированным жизненным формам и имеющие большую заблокированность рекомбинативной изменчивости (мекостемини, сфингонотини, бриодемини) окажутся менее пластичными. Они будут вымирать, и, может быть, даже придется позаботиться об их сохранении. Виды, же принадлежащие к группе мало или вовсе недифференцированных жизненных форм и обладающие соответственно низкой степенью блокировки рекомбинативной изменчивости (прус, сибирская кобылка), наоборот, окажутся очень живучими и даже повышающими свою численность. В этом случае придется подготовиться к борьбе с ними.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://insectalib.ru/ 'Насекомые - библиотека по энтомологии'